
盐度对暗纹东方鲀生长、非特异性免疫和抗氧化酶活力的影响
2014-11-05 06:32:52边平江邱成功徐善良林少珍
水生生物学报 2014年1期
边平江 邱成功 徐善良 林少珍,
(1. 浙江省海洋水产养殖研究所, 温州 325000; 2. 宁波大学海洋学院, 宁波 315211;3. 浙江省近岸水域生物资源开发与保护重点实验室, 温州 325000)
对水生生物而言, 盐度是重要的环境影响因子。盐度作为一种与渗透压密切相关的环境因子,对鱼类呼吸代谢、生长、存活及免疫防御影响显著[1]。养殖环境可能受到诸多人为因素和自然因素的影响而导致养殖区域盐度的变化, 盐度变化会导致鱼类耗氧率升高、代谢加速、对能量的需求增加,严重时甚至造成体内生理机能失常。同时, 盐度变化还能促使鱼体产生较多的自由基(ROS), 造成体内蛋白和脂质的氧化及 DNA损伤, 最终因细胞膜损伤和酶活力丧失影响细胞机能[2,3]。长期处于这种氧化压力之下, 将导致鱼体免疫防御能力和抗病力下降, 影响鱼类的正常生长[4]。
非特异性的体液免疫和细胞免疫系统是鱼类抵抗外来病原入侵的第一道防线, 溶菌酶(Lysozyme)是体液免疫因子, 由嗜中性粒细胞和巨噬细胞分泌,起溶菌作用[5]; 而碱性磷酸酶(AKP)是一种重要的调控酶, 直接参与磷酸基团的转移, 参与钙代谢,是动物溶酶体的重要组成部分, 在免疫反应中发挥重要的作用[6]。同时, 生物体内还存在抗氧化防御机制以应对氧化压力, 它能消除体内的自由基, 增强吞噬细胞防御能力、机体免疫功能, 维持细胞内部动态平衡。抗氧化系统主要由抗氧化酶和非酶类复合物构成, 其中, 抗氧化酶主要包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和谷胱甘肽 S-转移酶(GST)等[4,7]。目前, 国内外针对鱼类盐度胁迫的生理生化等方面开展了较多的研究。例如, 赵峰等[8]比较了盐度变化时施氏鲟不同组织的抗氧化酶活性的变换情况, 发现超氧化物歧化酶和过氧化氢酶在肝脏中的含量最高, 随着盐度的升高, 超氧化物歧化酶的活力呈现下降的趋势, 显著低于对照组; 而在心脏等中, 过氧化氢酶活力无显著差异; 当采用高盐度处理条石鲷时, 幼鱼肝脏中的超氧化物歧化酶和过氧化氢酶活力均受到较大的影响, 并最终形成新的平衡[4]; 同样, 降低盐度(10‰)可导致银鲳肾脏中谷胱甘肽 S-转移酶活力显著降低[9]; 而在盐度12‰和 18‰的处理组中, 斜带石斑鱼特定生长率显著高于盐度 6‰和高盐度处理组, 并且其血清超氧化物歧化酶和溶菌酶活性也显著高于其他组, 因此,适当降低盐度可能提高斜带石斑鱼养殖效益[10]。鱼类盐度变化相关方面研究的开展, 不仅有助于了解盐度变化时鱼体的生理学变化规律, 同时研究结果还能应用于养殖生产中, 改善养殖环境, 提高养殖效益。
暗纹东方 鲀(Takifugu obscurus)隶属于硬骨鱼纲,辐鳍亚纲, 鲈形总目, 鲀形目(Tetrodontiforms), 鲀科(Tetrodontidae), 东方鲀属(Takifugu)。在我国主要分布于渤海、东海、黄海及通海的江河中下游, 为近海与河川底栖鱼类[11]。其肉味鲜美、生长速度快、经济价值高, 开发利用前景广阔, 为我国重要养殖品种[12—13]。目前, 在暗纹东方 鲀的种质资源、人工繁育、生理及发育等方面均有研究报道[12—14]。有关盐度对暗纹东方 鲀的生理生化和生长影响的研究表明, 盐度变化能导致其血浆渗透压、鳃丝和肾脏中Na+-K+-ATPase酶活力显著变化[15], 还可能影响暗纹东方 鲀的生长速率[13]。本研究测定了盐度胁迫下暗纹东方 鲀脾脏组织中溶菌酶、碱性磷酸酶和肝脏组织中超氧化物歧化酶、过氧化氢酶的活力水平,并分析其生长状况, 旨在明确盐度胁迫过程中暗纹东方 鲀非特异性免疫酶活力和抗氧化酶活力的变化情况及对生长的影响, 为其健康养殖和水环境调控提供科学依据。
1 材料与方法
1.1 实验鱼养殖
暗纹东方 鲀购自江苏中洋集团河豚养殖厂, 实验前在2×104kg的水泥池中暂养1个月, 投喂河豚鱼饲料。养殖与暂养均采用曝气自来水, 暂养池为20 m2的水泥池, 注水水深0.8 m, 保持24h不间断充气, 每天换水1/3, 保持水质清新。选择无病无伤、行动迅速、生命力强的个体作为实验用鱼。实验暗纹东方 鲀的规格为体重(35.12±5.15) g、全长(12.12±0.95) cm。
1.2 实验方法
实验在3000 kg的PVE塑料缸中进行, 实验用水为一级过滤海水和经曝气的自来水。实验期间水温为20.5—23.9℃, 连续充气, 每2日换水一次。设置盐度梯度为6‰、12‰、18‰、24‰、30‰, 以淡水组作为对照, 每个梯度设 3个平行组, 用曝气的自来水和粗盐调节盐度, 每个设定盐度的缸中均放入随机选取的河豚90尾, 饲养50d。
1.3 非特异性免疫和抗氧化物酶活力测定
分别在实验第 0、第 1、第 5、第 10、第 15、第20、第30、第40和第50天取样进行酶活力测定,每个时间点每缸随机取5尾, 用MS-222进行快速麻醉, 解剖后取脾脏进行溶菌酶(LZM)和碱性磷酸酶(AKP)活力测定, 取肝脏进行超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活力测定, 试剂盒购自南京建成生物研究所。样品经匀浆和离心(4000 r/min,15min)后取得上清液待测。蛋白含量采用考马斯亮兰法测定, 以牛血清蛋白为标准。溶菌酶活性测定采用比浊法, 用酶标仪在22℃室温测定1—5min内吸光值(OD)变化, 以 OD值每分钟减少 0.001为 1个活性单位(U); 碱性磷酸酶活力采用比色法测定,测定在405 nm波长下的OD增高率; SOD活力采用黄嘌呤氧化酶法测定, 在波长为550 nm处比色测定吸收光度值计算其活力, 活力单位定义为: 每毫克组织蛋白在1 mL 反应液中SOD抑制率达到50%时所对应的SOD量为1个SOD活力单位(U/mg prot);CAT活力通过405 nm波长下测定H2O2减少的量测定,活力单位定义为: 每毫克组织蛋白每秒钟分解1 μmol的H2O2的量为1个CAT活力单位 (U/mg prot)。
1.4 生长指标计算方法
实验结束时, 对每个平行计算死亡率, 并称重和测量体长。按照下列公式计算相对增重率RW(%)和相对增长率RL(%)。

式(1、2)中, Wt为终末平均体重(g), W0为初始平均体重(g); Lt为终末平均体长(cm), L0为初始平均体长(cm)。
1.5 数据处理
实验结果用平均值±标准差(Mean±SE)表示。所有数据采用SPSS 10.0 统计软件进行单因素方差分析(One-way ANOVA), 采用Duncan法比较组间差异,当P<0.05时被认为差异显著。
2 结果
2.1 暗纹东方生长与死亡率
实验期间, 各组暗纹东方 鲀的生长率和死亡率如表 1所示。随着盐度的增高, 相对增长率逐渐降低, 而在 24‰和 30‰的高盐度组, 鱼体生长受到抑制, 相对增长率显著低于对照组; 同样, 随着盐度的增加, 鱼体的相对增重率也呈现相同的趋势, 其中盐度 24‰和 30‰的处理组中的相对增重率数值均显著低于对照组。在低盐度组(0、6‰和12‰)中,未发现死亡现象; 而在其他处理组中, 随着盐度的增加, 暗纹东方 鲀应激死亡率逐渐增加, 分别为5.93±1.70、8.89±3.33 和 17.04±2.80。
2.2 盐度对暗纹东方非特异性免疫酶活力的影响
结果表明, 盐度变化对暗纹东方 鲀脾脏中溶菌酶活力影响显著(图1)。在盐度处理 1d后各组的溶菌酶活力均逐渐增加, 其中盐度为 6‰的处理组中溶菌酶活力在处理后持续增加至第 20天并达到最高值, 其活力显著高于对照组(P<0.05); 随后, 溶菌酶活力开始降低并最终与对照组保持相似的水平。在盐度≥12‰的 4个处理组中, 暗纹东方 鲀的溶菌酶活力也随着处理时间的增加而逐渐升高, 并在第10(最高盐度组)或第 15天达到最高值, 显著高于对照组中的酶活力(P<0.05); 之后, 溶菌酶活力也开始逐渐降低并最终低于对照组水平, 特别是最高盐度(30‰)处理组, 其最终溶菌酶活力显著低于对照组水平(P<0.05)。
当经过相同的处理时间后, 不同的盐度处理组之间的溶菌酶活力也存在差异。在处理后的第5和第10天, 各组鱼体溶菌酶活力随着盐度的增高而逐渐升高, 并在最高盐度处理组达到最大值, 其活力显著高于淡水组(P<0.05); 而在处理15d以后, 高盐度组中鱼体溶菌酶活力降低, 特别是在处理40d后,最高盐度组中溶菌酶活力显著低于淡水组(P<0.05)。
表1 盐度对暗纹东方生长和存活的影响Tab. 1 Effects of salinities on the growth and mortality in Takifugu obscurus
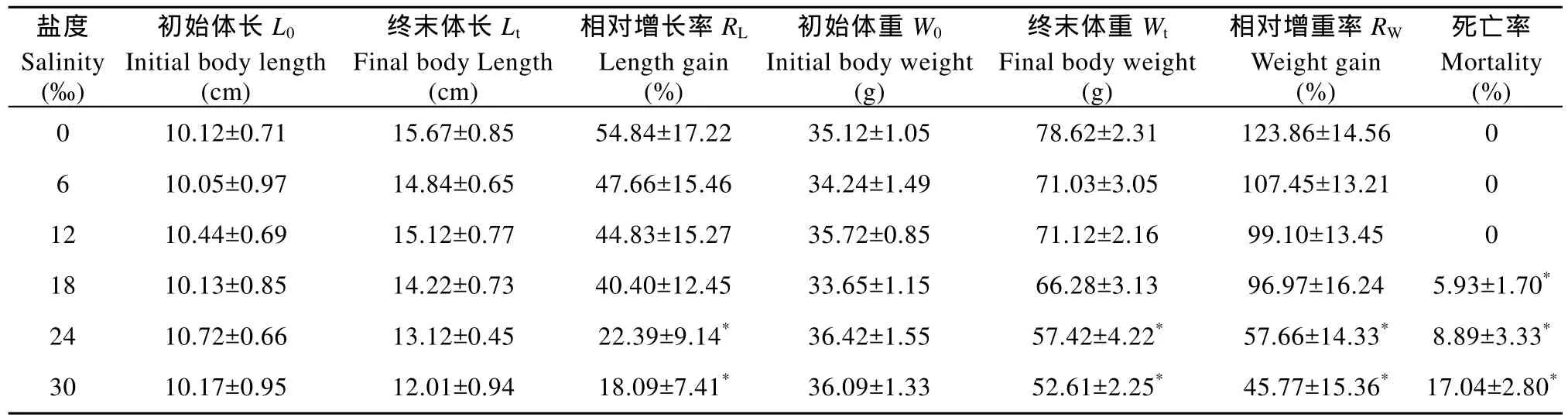
表1 盐度对暗纹东方生长和存活的影响Tab. 1 Effects of salinities on the growth and mortality in Takifugu obscurus
注: *表示差异显著(P<0.05)Note: * indicates significant difference (P<0.05)
盐度Salinity(‰)初始体长L0 Initial body length(cm)终末体长Lt Final body Length(cm)相对增长率RL Length gain(%)初始体重W0 Initial body weight(g)终末体重Wt Final body weight(g)相对增重率RW Weight gain(%)死亡率Mortality(%)0 10.12±0.71 15.67±0.85 54.84±17.22 35.12±1.05 78.62±2.31 123.86±14.56 0 6 10.05±0.97 14.84±0.65 47.66±15.46 34.24±1.49 71.03±3.05 107.45±13.21 0 12 10.44±0.69 15.12±0.77 44.83±15.27 35.72±0.85 71.12±2.16 99.10±13.45 0 18 10.13±0.85 14.22±0.73 40.40±12.45 33.65±1.15 66.28±3.13 96.97±16.24 5.93±1.70*24 10.72±0.66 13.12±0.45 22.39±9.14* 36.42±1.55 57.42±4.22* 57.66±14.33* 8.89±3.33*30 10.17±0.95 12.01±0.94 18.09±7.41* 36.09±1.33 52.61±2.25* 45.77±15.36* 17.04±2.80*
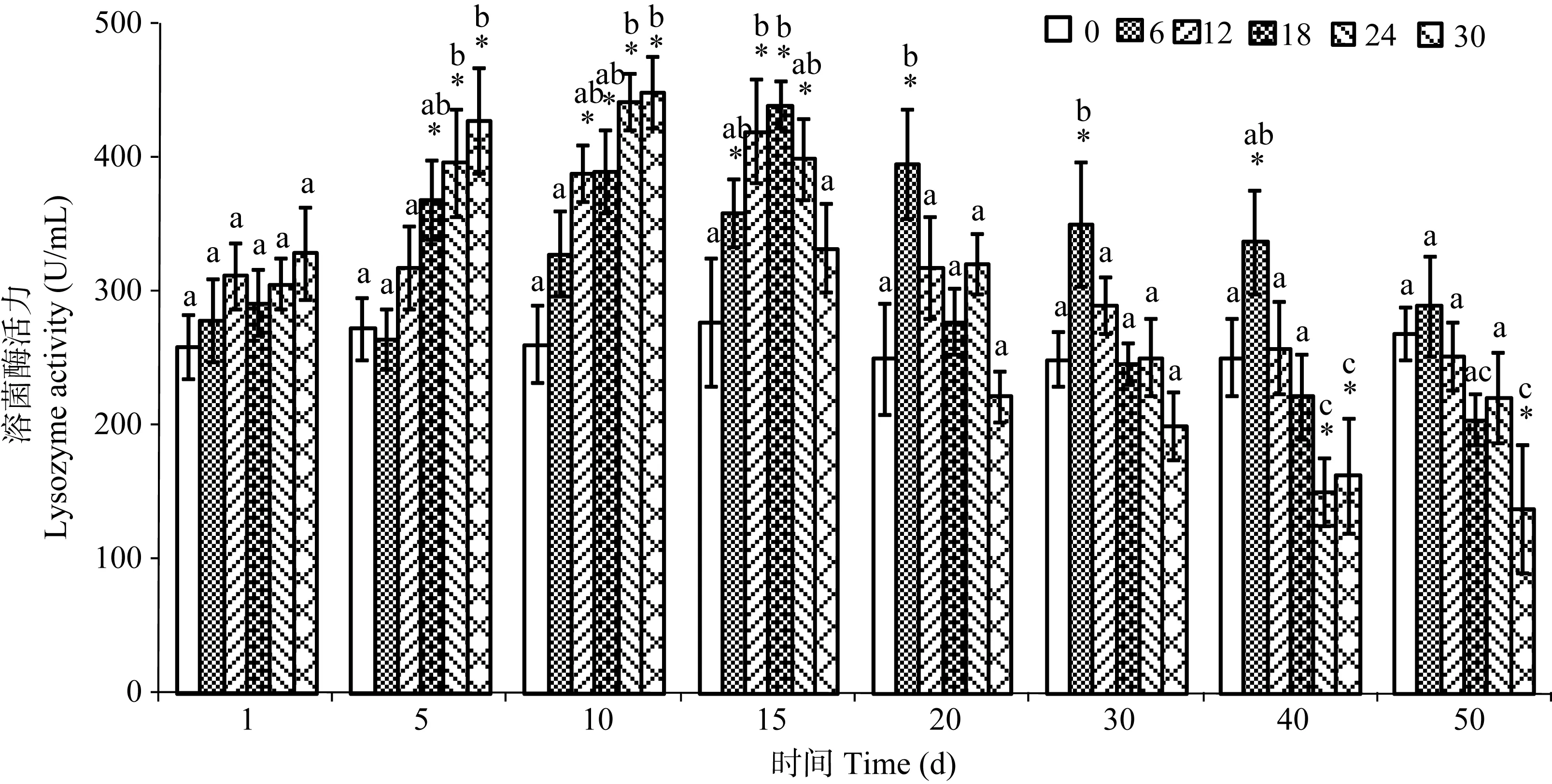
图1 不同盐度处理下暗纹东方 鲀的溶菌酶活力Fig. 1 Activity of lysozyme in Takifugu obscures under different salinity levels
盐度变化对暗纹东方 鲀脾脏中碱性磷酸酶(AKP)活力的影响如图2所示。在盐度为6‰和12‰的两个处理组中, 脾脏 AKP活力随着处理时间的延长而持续上升, 并在处理后第15天达到最大值,其活力显著高于对照组(P<0.05); 之后, AKP活力又逐渐降低, 并最终与对照组水平相似。盐度为18‰的处理组中, 脾脏 AKP活力也随着处理时间的增加而迅速增高, 并在处理后第 10 天达到最大值, 其活力显著高于对照组(P<0.05); 之后, 酶活力迅速降低, 最终水平稍低于对照组水平。盐度为24‰和 30‰的两个处理组中, 脾脏 AKP活力在处理后也逐渐上升, 并在处理第5天达到最高值; 之后, 酶活力逐渐降低并最终显著低于对照组(P<0.05); 特别是盐度为30‰的处理组, 其AKP酶在处理10d后活力一直较低, 显著低于对照组水平(P<0.05)。
比较发现, 经过相同的处理时间后, 不同的盐度处理组之间的碱性磷酸酶活力也存在差异。在处理的第5、第10和第15天, 最高碱性磷酸酶活力出现的盐度逐渐前移, 分别出现在盐度为 24‰、18‰和 12‰的处理组中, 且随着盐度的增加, 酶活力逐渐降低; 在处理的第10至第30天, 最高盐度组中碱性磷酸酶活力显著低于淡水组(P<0.05); 随后, 各组之间的活力值差异有所减少。
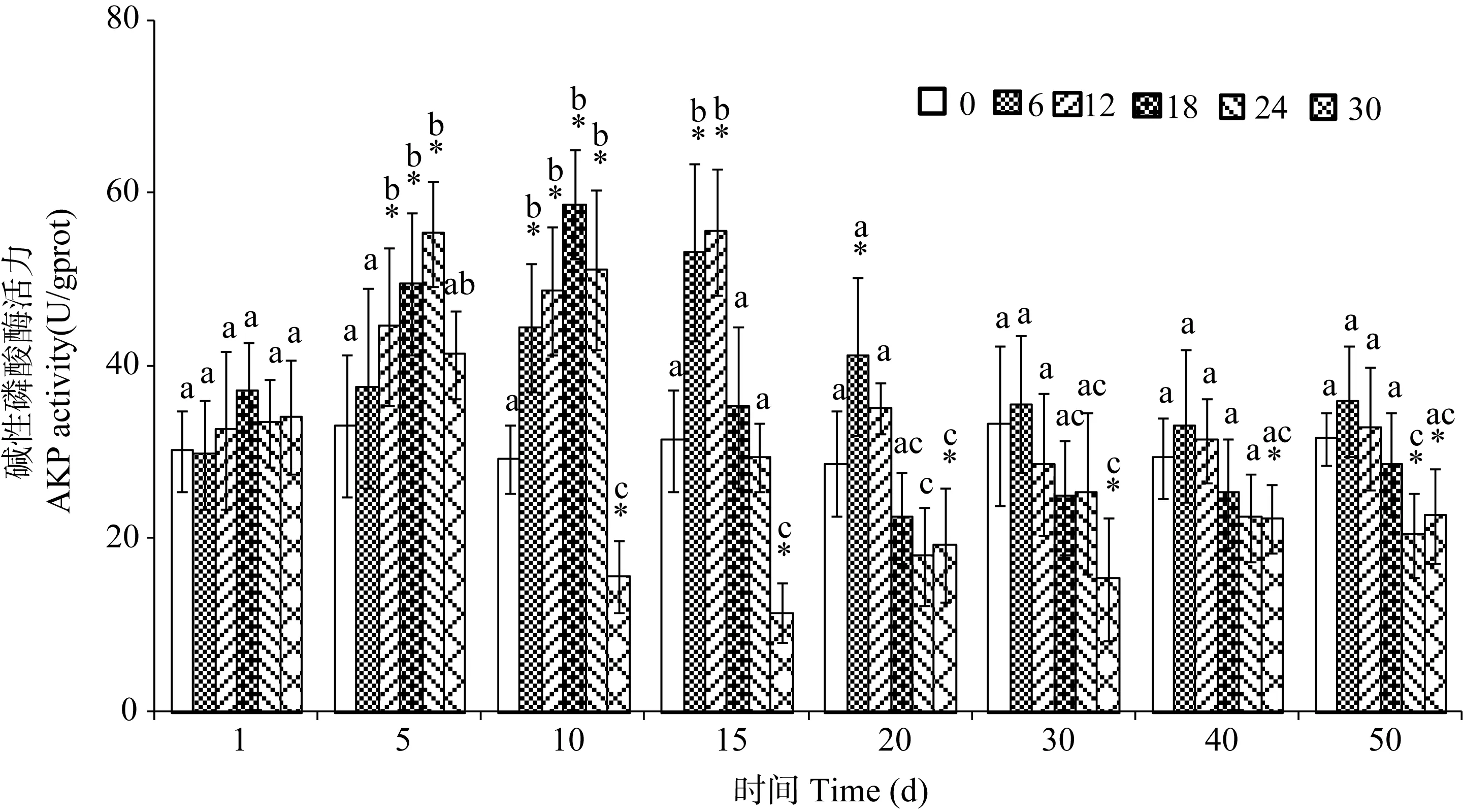
图2 不同盐度处理下暗纹东方鲀的碱性磷酸酶活力Fig. 2 Activity of AKP in Takifugu obscures under different salinity levels
2.3 盐度对暗纹东方抗氧化酶活力的影响
盐度处理后暗纹东方 鲀的超氧化物歧化酶(SOD)活力变化如图3所示。结果表明, 在处理初期各处理组中暗纹东方 鲀肝脏 SOD活力均呈现增高的趋势; 除了最高盐度处理组外, 其余各组在处理第10天时SOD酶活力达到最大并显著高于对照组(P<0.05), 之后, 逐渐降低并恢复至对照组水平。在盐度为30‰的最高盐度处理组中, SOD酶活力最大值出现在处理后第 5天, 随后酶活力开始降低, 并最终与对照组无显著性差异(P>0.05)。而经相同处理时间后, 除了处理后第5至第10天外, 其余时间的各盐度组间差异不大。
盐度变化对暗纹东方鲀过氧化氢酶(CAT)活力的影响如图4所示。在盐度6‰的处理组中, 肝脏中CAT活力随着处理时间的增长而逐渐增高, 并在处理后第 5天达到最高峰; 之后, 酶活力开始下降并最终保持与对照组相似的水平。在其他处理组中,当处理1天时CAT活力处于较高水平, 但是与对照组无显著性差异(P<0.05); 之后, CAT活力逐渐降低,并在处理后第15天时达到最小值, 其中盐度较高的处理组(盐度分别为18‰、24‰和30‰)中的酶活力显著低于对照组水平; 随后, 肝脏中 CAT活力又开始回升, 并最终接近对照组水平, 与对照组无显著性差异(P>0.05)。经过相同的处理时间, 在处理后第10至第 15天时, 高盐度组酶活力显著降低(P<0.05)。而在其他时间点, 不同盐度组间过氧化氢酶活力变化不大。
3 讨论
3.1 盐度变化对暗纹东方鲀非特异性免疫能力的影响

图3 不同盐度处理下暗纹东方鲀的SOD活力Fig. 3 Activity of SOD in Takifugu obscures under different salinity levels

图4 不同盐度处理下暗纹东方鲀的CAT活力Fig. 4 Activity of CAT in Takifugu obscures under different salinity levels
非特异性体液免疫和细胞免疫系统是鱼类抵抗外来病原入侵的第一道防线。溶菌酶在鱼体的黏液、血清和某些淋巴组织中广泛分布。它参与机体免疫活动, 是鱼类防御微生物入侵的最有效因子之一。研究表明, 盐度变化能影响多种鱼类体内溶菌酶活力。当牙鲆(Paralichthys olivaceus)培育盐度升高或者降低时,溶菌酶活力将会升高, 并在处理第5天时达到最高值,随后逐渐降低[16]; 军曹鱼(Rachycentron canadum)在低盐度处理后, 溶菌酶活力升高, 而随着处理时间的增加也逐渐降低至于对照组相似的水平[17]; 随着海水盐度的降低, 许氏平(Sebastes schlegeli)的溶菌酶活力在处理 24h时达到最高峰值, 之后逐渐下降[18]; 当用盐度处理日本鳗鲡(Anguilla japonica)时,溶菌酶活力也随着盐度的增加而增高, 之后也是逐渐恢复至对照组水平[19]。本研究结果发现, 溶菌酶在暗纹东方鲀非特异性免疫中也具有重要的作用。在处理初期, 溶菌酶活力增加以应对外源压力; 但是, 鱼体在经受长期的胁迫之后, 自身各方面的技能势必受到影响, 从而降低了自身的抵抗力, 这也将影响鱼体的生长甚至存活情况。
当养殖在不同盐度时, 日本鳗鲡体内碱性磷酸酶活力随盐度的增加而升高, 表明其在鱼类免疫中具有重要的作用; 同时, 其活力变化规律也与鱼体生长变化规律呈正相关, 推测AKP活力的降低可能有碍机体内钙磷代谢的正常进行,从而影响鱼体的生长[19,20]。同样, 本研究发现暗纹东方鲀AKP酶活力也随盐度处理时间的增加而变化, 且酶活力与暗纹东方鲀的生长也呈一定的对应关系, 推测高盐度组中AKP酶活力的降低可能也影响了暗纹东方鲀机体磷钙代谢的正常进行, 致使其生长受到影响, 但这尚需进一步研究的验证。
同时, 多重比较结果发现不同盐度处理对于暗纹东方鲀的免疫相关酶活力的影响存在显著差异, 而这种差异多表现在处理后第 5至第20天内。推测该阶段是暗纹东方鲀主要的胁迫后应激反应和自身平衡调节的关键时期。而对于其具体的调节机制, 还有待研究的继续深入。
3.2 盐度变化对暗纹东方鲀抗氧化能力的影响
超氧化物歧化酶(SOD)是超氧阴离子自由基(O2–)的清除剂, 过氧化氢酶能将过氧化氢(H2O2)转变为水,二者可保护机体细胞免受活性氧自由基的伤害, 是抗氧化防御体系的关键酶[21]。研究表明, 许氏平急性低盐胁迫96h后, 血液SOD和CAT活力出现波动, 且较初始值均有不同程度的降低; 而当许氏平处于低盐度水体中30d后, 其血液中SOD和CAT活力明显升高[18]。条石鲷在低盐度处理后, 其SOD也呈现增高的趋势, 随后逐渐恢复至对照组水平[4]。当用盐度对日本鳗鲡进行处理时, 其体内SOD活力也出现先升高后降低至对照组水平的情况[19]。
在本研究中, 暗纹东方鲀经盐度处理后, SOD和 CAT的变化趋势与其他鱼类相似, 表明 SOD和CAT在暗纹东方鲀抗氧化防御系统中具有重要的作用。盐度胁迫导致鱼体内产生大量活性氧自由基,而SOD和CAT活力升高可清除这些自由基, 从而保护体内细胞免受其伤害, 而这也主要发生在处理后的第5至第20天; 随着鱼体对盐度的逐渐适应, 其抗氧化系统重新建立平衡, 抗氧化酶活性逐渐恢复至与对照组无显著差异。
3.3 盐度变化对暗纹东方鲀生长的影响
当鱼类受到环境胁迫时, 机体处于应激状态,导致鱼类耗氧率升高、对能量的需求增加、代谢加速、体内生理机能失常, 并容易受到环境中细菌的侵入, 短期内非特异性免疫相关酶和抗氧化酶活力的升高有助于增加鱼体对外界环境的抵抗力。而随着胁迫时间延长, 可能会因鱼体多种生理功能的紊乱而导致溶菌酶等非特异性免疫因子的功能抑制,鱼体免疫防御能力和抗病力下降。在高盐度水环境中, 暗纹东方鲀鱼体为应对胁迫应激而导致能量消耗和多种生理变化, 推测这种影响是导致鱼体生长受到抑制, 甚至出现死亡的原因。
不同鱼类对于盐度的耐受力不同, 而同种鱼类在发育不同时期的盐度耐受力也存在差异。Yang, et al.[22]研究了盐度对暗纹东方鲀胚胎发育的耐受力,发现其可以耐受 8‰的盐度, 当处理盐度低于 8‰时胚胎发育和孵化没有差异。Shi, et al.[13]对孵化后第3至第 19天的暗纹东方鲀初孵仔鱼进行了盐度处理,结果发现, 当处理盐度为 25‰时将严重降低仔鱼的生长率和存活率, 从处理后第 11天开始, 盐度 25‰处理组中仔鱼的全长、特定生长率和存活率均显著低于低盐度组和对照组; 当处理盐度低于 20‰时, 存活率没有明显的差异, 推测暗纹东方鲀最适合的仔稚鱼培育盐度为8‰—10‰。本实验结果表明, 随着盐度的增高, 暗纹东方鲀幼鱼的相对增长率逐渐降低, 24‰—30‰的高盐度可显著地抑制鱼体生长; 同样, 随着盐度的增加, 鱼体的相对增重率也逐渐降低,而24‰和30‰的高盐度可导致处理组中鱼体的相对增重率显著降低。在低盐度组(0、6‰和12‰)中, 没有出现死亡现象, 但随着盐度的增加, 暗纹东方鲀开始出现死亡个体, 且死亡率随着处理盐度的增高逐渐增加。可见, 暗纹东方鲀幼鱼和仔鱼具有相似的盐度耐受能力, 能适应一定程度的盐度变化, 且当培育盐度低于12‰时, 生长效果优于高盐度处理组。
猜你喜欢
智族GQ(2024年5期)2024-06-03 15:15:11
农产品质量与安全(2023年1期)2023-02-18 14:22:04
成都大学学报(自然科学版)(2021年1期)2021-05-22 01:31:18
水电与抽水蓄能(2021年2期)2021-05-14 01:42:26
农业工程学报(2021年4期)2021-05-09 08:23:58
农业工程学报(2020年6期)2020-05-19 07:46:38
睿士(2020年12期)2020-01-03 10:21:20
太阳能(2014年5期)2014-05-12 12:57:18
食品工业科技(2014年9期)2014-03-11 18:15:56
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:21:01
