
细胞周期蛋白依赖性激酶抑制剂Roscovitine对3种蓝藻生长和活性的影响
2014-10-25 08:40:42周晓潮王春梅
微生物学杂志 2014年2期
周晓潮,王春梅
(北京中医药大学中药学院,北京 100102)
随着越来越多的原核生物基因组测序工作的完成,一些有趣的信息引起了科学界的关注,如原核中竟然存在大量丝氨酸/苏氨酸激酶,它们是否与真核生物中的激酶一样可作为细胞生长分裂与分化的信号调控系统的组成部分?在已研究的21个蓝藻菌株基因组中,发现了286个基因编码丝氨酸/苏氨酸蛋白激酶(serine/threonine protein kinase,STPK),其中,鱼腥藻 7120(Anabaena sp.PCC 7120)含有52个基因编码 STPK[1]。尽管这些STPK的功能大多还不明确,但如此大量的基因表明,其编码的蛋白在蓝藻细胞生命中一定起到了重要的作用。真核细胞中,细胞周期蛋白依赖性激酶(cyclin-dependent kinase,CDK)属于丝氨酸/苏氨酸蛋白激酶家族[2],与细胞周期蛋白(cyclin)结合后被激活,可催化底物磷酸化[3]。CDKs在细胞周期循环过程中发挥重要作用,依序完成DNA合成和有丝分裂,引起细胞的生长和增殖[4]。Roscovitine 是 CDC2/cyclin B、CDK2/cyclin A、CDK2/cyclin E、CDK5/p35的选择性抑制剂。可竞争性地与CDKs的腺嘌呤位点结合,从而将细胞阻止于G1/S期和G2/M期,使细胞在G2期积累,最终导致死亡[5]。蓝藻中也存在与CDKs基因序列同源性较高的激酶基因,这些蓝藻中的激酶是否也与细胞分裂调控相关,本文考察了CDKs的抑制剂roscovitine对丝状多细胞蓝藻鱼腥藻7120、单细胞蓝藻集胞藻6803(Synechocystis sp.PCC 6803)和聚球藻 7002(Synechococcus sp.PCC 7002)生长的影响,将为进一步揭示STPK在蓝藻细胞分裂过程中的作用奠定基础。
1 材料与方法
1.1 材料
1.1.1 材料与试剂 丝状蓝藻鱼腥藻7120(Anabaena sp.PCC 7120)由中国科学院植物研究所施定基博士赠送,单细胞蓝藻集胞藻6803(Synechocystis sp.PCC 6803)、单细胞蓝藻聚球藻7002(Synechococcus sp.PCC 7002)由中国科学院植物研究所黄芳博士赠送;实验用试剂Roscovitine和二甲基亚砜(DMSO)为Sigma公司产品,MTT(3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)为Amresco公司产品。
1.1.2 培养基和培养条件 鱼腥藻7120和集胞藻6803采用BG-11(加氮)培养基[6],置于转速为120 r/min,培养温度(28±1)℃,光照强度约为80 μmol/m2/s的HPG-280BX光照控温旋转培养箱(哈尔滨市东联电子技术开发有限公司)中培养。聚球藻7002采用Medium A培养基,置于转速为120 r/min,培养温度(30±1)℃,光照强度约为80 μmol/m2/s的光照控温旋转培养箱中培养。
1.2 方法
1.2.1 实验设计 选取3种藻对数生长期的细胞培养物,接种于相应的新鲜培养基中,起始OD750达0.3左右。将上述每种藻培养液分别加置A、B两块24孔板上,每孔培养体积1 mL。每块板分别设置空白组、对照组、加药组,每组3个平行。空白组分别为BG-11(加氮)或Medium A培养基,对照组为不加roscovitine的藻培养液,加药组中 roscovitine的浓度分别为50、100、200和300 μmol/L。A板培养至4 h,B板培养至 4 d取样。
1.2.2 显微镜下观察藻细胞形态 取各组藻细胞液20 μL于玻片上,使用TE2000-S倒置显微镜(日本,Nikon),在10倍目镜、40倍物镜下进行观察,照相。
1.2.3 不同浓度roscovitine对3种藻生长的影响使用UV2100紫外分光光度计(美国,UNICO)测取波长750 nm下,3种藻各组藻培养液的吸光度值[7]。
1.2.4 MTT法测3种藻的活力变化[8-9]从上述各板中每孔分别取300 μL藻培养液至1.5 mL EP管中,向培养液中加入100 μL浓度为0.5 mg/mL的MTT溶液,转速为120 r/min,培养温度(35±1)℃,光照强度约为 80 μmol/m2/s,孵育 2.5 h。12000 r/min离心10 min,弃上清,向沉淀中加DMSO(A 板:150 μL;B 板:750 μL),充分溶解。12000 r/min离心10 min,取上清100 μL 至96 孔板,使用ELx800酶标仪(美国,BioTek)测取波长490 nm下的吸光度值。
2 结果与分析
2.1 形态观察

图1 roscovitine作用4 d对3种蓝藻生长的影响Fig.1 The effects of roscovitine on the growth of three species of cyanobacterial strains after treated 4 days

图2 不同浓度roscovitine作用4 h和4 d对鱼腥藻7120生长的影响Fig.2 The effects of roscovitine on the growth of Anabaena sp.PCC 7120 after treated 4 hour or 4 days
如图1所示,鱼腥藻7120加药组藻丝状体较短,所含藻细胞数较少;对照组丝状体较长,所含藻细胞数较多。集胞藻6803和聚球藻7002加药组与对照组比较,藻细胞在形态上无差别。
2.2 Roscovitine对鱼腥藻7120生长和活性的影响
研究了短期(4 h)和较长期(4 d)roscovitine对3种藻的作用效果。藻液吸光度OD750数值反映了roscovitine对细胞数的影响。MTT检测结果OD490数值反映了细胞的活性。如图2A所示,加药4 h,不同浓度roscovitine对鱼腥藻7120的细胞数没有影响,加药4 d,浓度大于100 μmol/L的roscovitine对鱼腥藻7120的细胞数具有明显的降低作用(P<0.05),且 roscovitine浓度越高细胞数越少。图2B显示加药4 h,浓度大于100 μmol/L的roscovitine对鱼腥藻7120细胞活性有极显著的抑制作用(P<0.01),且这种抑制作用呈剂量依赖性;加药4 d 200 μmol/L的roscovitine对细胞活性有显著的抑制作用(P<0.05),300 μmol/L的roscovitine对细胞活性有极显著的抑制作用(P <0.01)。
2.3 Roscovitine对集胞藻6803生长和活性的影响
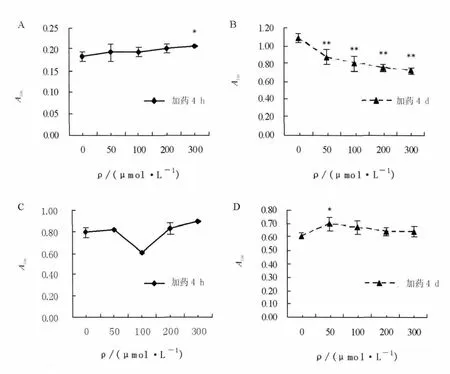
图3 不同浓度roscovitine作用4 h和4 d对集胞藻6803生长的影响Fig.3 The effects of roscovitine on the growth of Synechocystis sp .PCC 6803 after treated 4 hour or 4 days
如图 3A 所示,加药 4 h,300 μmol/L 的roscovitine对集胞藻6803有轻微的刺激作用,短时间内增加了细胞数(P<0.05);加药4 d,不同浓度的roscovitine均对集胞藻6803的细胞数具有极显著的降低作用(P<0.01),且roscovitine浓度越高细胞数越少。图3B显示加药4 h,不同浓度roscovitine对集胞藻6803细胞活性无显著的抑制作用(P>0.05);图3C显示加药4 d,只有50 μmol/L的roscovitine对细胞活性促进作用(P<0.05),其他浓度无影响。
2.4 Roscovitine对聚球藻7002生长和活性的影响
如图4A所示,加药4 h,不同浓度的roscovitine均对聚球藻7002细胞数具有极显著的增多作用 (P<0.01),且roscovitine浓度越高细胞数越多;加药4 d,不同浓度roscovitine对聚球藻7002的细胞数无影响 (P>0.05)。图4B显示加药4 h,不同浓度 roscovitine对聚球藻7002细胞活性有显著的增强作用 (P<0.05),且roscovitine浓度越高细胞活性越强;图4C显示加药4 d,不同浓度roscovitine对细胞活性无影响 (P>0.05)。
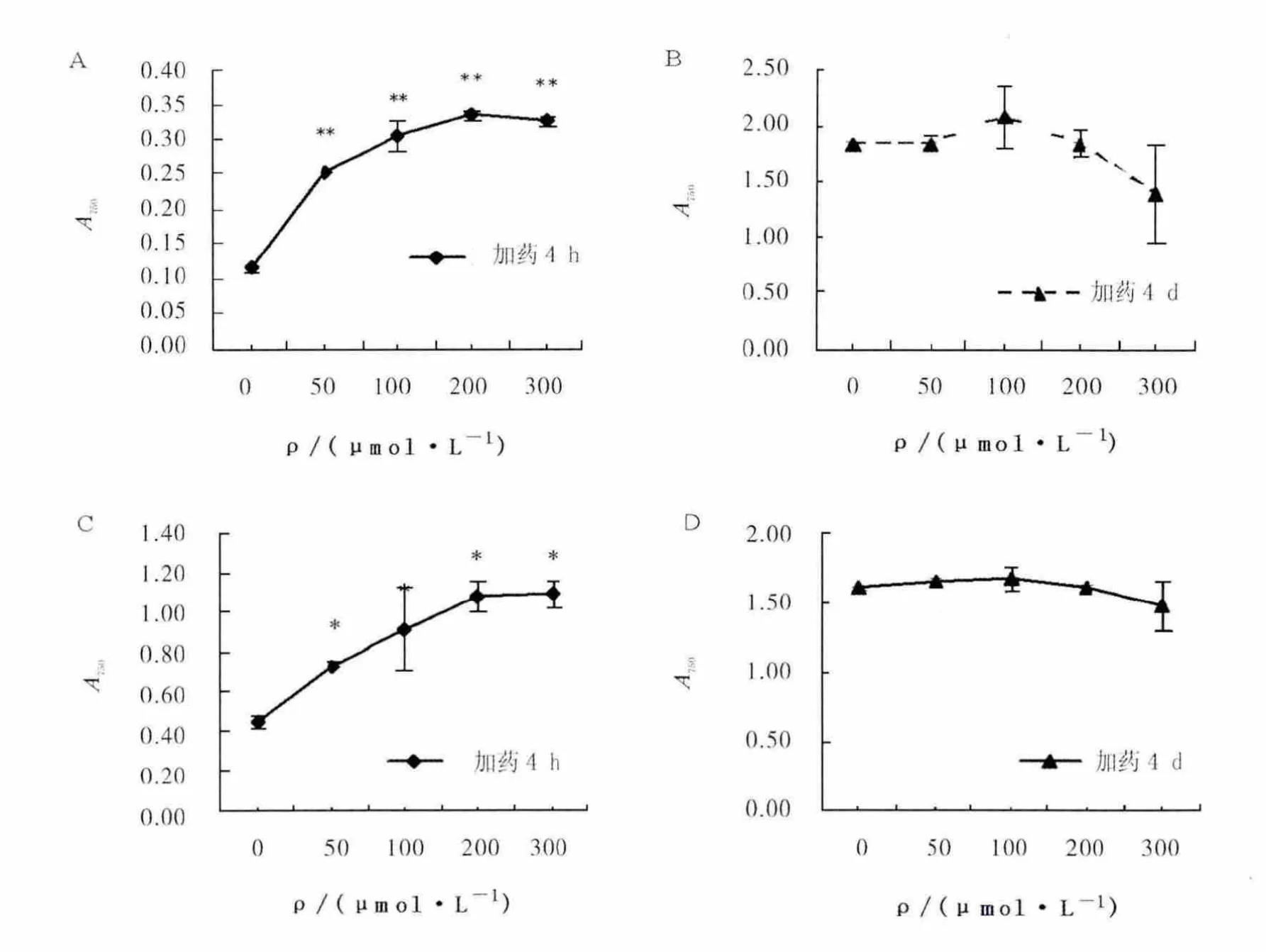
图4 不同浓度roscovitine作用4 h和4 d对聚球藻7002生长的影响Fig.4 The effects of roscovitine on the growth of Synechococcus sp .PCC 7002 after treated 4 hour or 4 days
3 讨论
本研究结果显示,roscovitine抑制丝状多细胞鱼腥藻7120的活性,从而抑制藻的生长。从形态上,长时间作用后的鱼腥藻7120藻丝体较短。roscovitine短时间作用对聚球藻7002的活性和生长有刺激作用,但长时间作用不显著。对于单细胞集胞藻6803,虽然roscovitine并没显示出对其活性的抑制,但长时间作用后藻的生长受到抑制。从以上结果来看,roscovitine影响了蓝藻的生长。roscovitine对不同的蓝藻表现出不同的影响,可能对于不同蓝藻其作用靶点不同,表现出不同程度的效应。roscovitine影响了每种蓝藻细胞中的哪些激酶还有待深入研究。
对蓝藻中激酶的功能研究已经引起多个实验室的注意。鱼腥藻7120的PknA(Alr4366)是第一个被发现的在丝氨酸/苏氨酸位点上磷酸化的蛋白激酶,是蓝藻正常生长和分化所必需的[10]。pkn41(alr0709)和 pkn42(alr0710)参与氮代谢调控[11],PknD(Alr4368)是固氮条件下正常生长所必需的,可能参与异形胞与营养细胞间氮代谢产物的运输[12]。Pkn30和Pkn44用于合成异形胞特定糖脂 HGL2[13],HepS(All2760)在异形胞多糖层的形成过程中起调节作用[14]。pknE(alr3732)参与异形胞被膜的形成,并间接控制固氮作用[15]。pknH(alr1336)只在异形胞中表达,用于稳定地连接异形胞与营养细胞[16]。目前对于鱼腥藻STPK的研究主要集中在细胞分化过程,异形胞形成和氮代谢调控三方面,关于STPK在细胞分裂过程中的作用未见报道,但细胞分裂过程与蓝藻的生长密切相关。在真核生物中STPK参与调控细胞分裂,那么在原核生物中STPK是否还具有这一作用呢?roscovitine是真核细胞分裂的抑制剂,在本研究中显示能抑制鱼腥藻7120的生长,推测roscovitine可能抑制了蓝藻细胞的分裂。roscovitine对鱼腥藻7120生长和活性的影响是通过抑制某一种或某一类蛋白起作用的,其结构和功能与真核细胞中的CDKs类似,参与调控细胞的生长与增殖。因其与roscovitine结合活性受到影响,从而抑制了鱼腥藻7120的生长和活性。在 NCBI(National Center for Biotechnology Information)中对鱼腥藻7120的总蛋白进行序列比对,寻找与人CDK1(NCBI登录号:P06493)、CDK2(NCBI登录号:CAA43985)和CDK5(NCBI登录号:CAG33322)类似的蛋白激酶。检索结果显示,与人的CDK1进行比对,有一个蛋白总分值最高,为82.0,相似度为30%;总分值在50~80的蛋白有20个,相似度在23% ~29%之间,其中包含已知的蛋白激酶PknE。与CDK2和CDK5进行序列比对,总分值在50~80的蛋白分别有33和24个,相似度分别在24% ~31%和23% ~35%之间,其中已知的蛋白激酶有PknA、PknD和PknE。另外还有许多蛋白与CDK1、CDK2和CDK5有相对更低的相似性,分值小于50。上述所有蛋白均为丝氨酸/苏氨酸激酶,而roscovitine很可能作用于这些蛋白中的某一个或几个,从而抑制了鱼腥藻7120的活性和分裂。此推测还有待进一步深入研究确定。
[1]Zorina,A.A.Eukaryotic protein kinases in cyanobacteria[J].Russian Journal of Plant Physiology ,2013,60(5):589-596.
[2]Pavletich N P.Mechanisms of cyclin-dependent kinase regulation:structures of cdks,their cyclin activators,and cip and INK4 inhibitors[J].Journal of molecular biology,1999,287(5):821-828.
[3]Sharma P S,Sharma R,Tyagi R.Inhibitors of cyclin dependent kinases:useful targets for cancer treatment[J].Current cancer drug targets,2008,8(1):53-75.
[4]Malumbres M,Barbacid M.Cell cycle,CDKs and cancer:a changing paradigm[J].Nature Reviews Cancer,2009,9(3):153-166.
[5]Meijer L,Borgne A,Mulner O,et al.Biochemical and cellular effects of roscovitine,a potent and selective inhibitor of the cyclin-dependent kinases cdc2,cdk2 and cdk5[J].European Journal of Biochemistry,1997,243(1-2):527-536.
[6]Castenholz R W.Culturing methods for cyanobacteria[J].Methods in enzymology,1988,167:68-93.
[7]金传荫,宋立荣.鱼腥藻 HB1017株化能异养生长的研究[J].水生生物学报,1995,19(1):13-20.
[8]Li J,Song L.Applicability of the MTT assay for measuring viability of cyanobacteria and algae,specifically for Microcystis aeruginosa(Chroococcales,Cyanobacteria)[J].Journal Information,2007,46(5):593-599.
[9]吴窈画,谈书华,范超超,等.MTT法检测细菌细胞数的主要影响因素分析[J].微生物学杂志,2011,31(3):67-72.
[10]Zhang C C.A gene encoding a protein related to eukaryotic protein kinases from the filamentous heterocystous cyanobacterium Anabaena PCC 7120[J].Proceedings of the National A-cademy of Sciences,1993,90(24):11840-11844.
[11]Cheng Y,Li J H,Shi L,et al.A pair of iron-responsive genes encoding protein kinases with a Ser/Thr kinase domain and a His kinase domain are regulated by NtcA in the cyanobacterium Anabaena sp.strain PCC 7120[J].Journal of bacteriology,2006,188(13):4822-4829.
[12]Zhang C C,Libs L.Cloning and characterisation of the pknD gene encoding an eukaryotic-type protein kinase in the cyanobacterium Anabaena sp.PCC7120[J].Molecular and General Genetics MGG,1998,258(1-2):26-33.
[13]Shi L,Li J H,Cheng Y,et al.Two genes encoding protein kinases of the HstK family are involved in synthesis of the minor heterocyst-specific glycolipid in the cyanobacterium Anabaena sp.strain PCC 7120[J].Journal of bacteriology,2007,189(14):5075-5081.
[14]Tom S K,Callahan S M.The putative phosphatase All1758 is necessary for normal growth,cell size and synthesis of the minor heterocyst-specific glycolipid in the cyanobacterium Anabaena sp.strain PCC 7120[J].Microbiology,2012,158(2):380-389.
[15]Zhang C C,Friry A,Peng L.Molecular and Genetic Analysis of Two Closely Linked Genes That Encode,Respectively,a Protein Phosphatase 1/2A/2B Homolog and a Protein Kinase Homolog in the CyanobacteriumAnabaena sp.Strain PCC 7120[J].Journal of bacteriology,1998,180(10):2616-2622.
[16]Ehira S,Ohmori M.The pknH gene restrictively expressed in heterocysts is required for diazotrophic growth in the cyanobacterium Anabaena sp.strain PCC 7120[J].Microbiology,2012,158(Pt 6):1437-1443.
猜你喜欢
湖泊科学(2022年4期)2023-01-04 14:20:54
可再生能源(2022年8期)2022-08-17 06:37:52
东坡赤壁诗词(2022年2期)2022-04-15 02:32:43
东坡赤壁诗词(2019年5期)2019-11-14 10:36:10
当代水产(2019年9期)2019-10-08 08:02:42
大自然探索(2019年2期)2019-03-01 02:23:30
当代水产(2018年8期)2018-11-02 05:30:42
生态学报(2018年14期)2018-08-29 11:08:32
上海农业学报(2016年5期)2016-02-10 06:53:01
分析测试学报(2015年9期)2015-12-17 16:44:27
