
乳山湾及邻近海域桡足类种类组成和时空分布的初步研究*
2014-10-16 07:22:12徐潇峰朱丽岩徐风风巩文静王俊健
中国海洋大学学报(自然科学版) 2014年4期
徐潇峰,朱丽岩,徐风风,巩文静,王俊健
(中国海洋大学海洋生命学院,山东 青岛266003)
乳山湾位于山东半岛南部海域,作为呈Y字型的半封闭海湾,其沿海滩涂广阔,是天然的海洋经济生物的养殖场所[1-2],然而几十年的海水养殖和工业发展使得乳山湾附近的海洋环境受到了一定的影响,氮磷等营养物质的增加和化学需氧量(Chemical oxygen demond,COD)含量的升高表明,该海域生态环境出现了一定程度的恶化[3-4]。因此,开展对乳山湾及邻近海域的生物生态研究对于了解该海域环境质量现状,实现海洋生态环境保护和资源开发利用协调发展具有重大意义。
桡足类动物隶属于节肢动物门(Arthropoda)甲壳亚门(Crustacea)桡足纲(Copepoda),广泛分布于世界海洋中,其种类和数量在海洋浮游动物群落中均处于优势地位,是海洋浮游动物重要的组成部分[5-8]。在海洋生态功能方面,桡足类在能量流动和物质循环中也有着不可忽视的作用。研究表明,它既是浮游植物等初级生产者的主要摄食者,又是包括许多经济鱼类在内的上层捕食者的主要食物来源,是海洋生态系统中初级生产力与高营养级之间的重要枢纽[9-11]。海洋桡足类自身较差的游泳能力决定了其对于水体环境变化敏感的特性,其种类组成和数量变动都能反映其所处生态环境的变化[12-13]。因而开展对桡足类的研究对于了解其所处区域的生态环境状况和生态系统功能具有重要意义[14-15]。与此同时,国内对于乳山湾及附近海域桡足类的研究报道也较为少见。由此,本文拟通过对乳山湾及邻近海域浮游桡足类的种类组成和分布情况进行分析和探讨,为了解当地的海洋生态资源状况、环境评价与保护等提供基础资料和科学依据。
1 材料与方法
分别于2011年5和10月在乳山湾及附近海域进行春季和秋季2个航次的生态调查,共设20个站位(见图1),包括湾内站位8个,湾外海域站位12个。根据《海洋调查规范GB/T 12763.6-2007》,采用浅水Ⅰ型网和浅水Ⅱ型浮游生物网进行由底到表的垂直拖网。2个航次共采得浮游动物样品80瓶,样品加入5%甲醛溶液固定,并带回实验室在体式显微镜下进行分类鉴定和计数。

图1 站位分布图Fig.1 The map of sampling stations
分别选取采样过程中同步测得的水温和盐度以及实验室后期分析测得的浮游植物密度与桡足类香农威纳多样性指数H′进行pearson相关性分析。其中水温和盐度数据来自国家海洋局威海市海洋监测中心,浮游植物密度数据来自中国海洋大学海洋生命学院浮游植物实验室。
参照文献[16],桡足类优势度计算公式为:

式中:ni是桡足类第i个种的丰度值;fi是第i个种出现的频率;N为桡足类总丰度值;优势度大于0.02的种类为优势种。
参照刘红等[17]研究,群落多样性分析采用香农威纳多样性指数(H′)、Pielou均匀度指数(J)和 Margalef丰富度指数(d)进行:

式中:H′为多样性指数;Pi为第i个种的丰度ni与桡足类总丰度N 的比值(ni/N);S为样品中总种类数。
2 结果
2.1 种类组成和分布
不计桡足类幼体和无节幼虫,本次调查共采集鉴定出桡足类28种,其中包括哲水蚤19种,剑水蚤4种,猛水蚤5种。在春季航次共采集到桡足类22种,其中,湾内采集到全部种类,而湾外则有19种。秋季航次共采集到桡足类20种,其中湾内仅有12种,而湾外达到了19种。海湾内外桡足类的种类分布不均匀,在湾内秋季各个站位的种类数要明显少于春季,而在湾外,春秋季节种类数差距并不明显(见表1、图2)。
2.2 桡足类丰度的平面分布和季节分布
本次调查中,春季航次桡足类平均丰度为13 967.19ind/m3,其 中 湾 内 平 均 丰 度 为 10 698.95 ind/m3,湾外为15 544.97ind/m3。整体上看,春季桡足类丰度平面分布不均匀,低值区出现在湾口附近海域,并逐渐向湾内和湾外递增(见图3a)。
在秋季航次,桡足类平均丰度为12 044.52 ind/m3,其中湾内平均丰度达到17 308.98ind/m3,湾外仅为8 534.87ind/m3。整体上看,桡足类丰度密集区(>24 000ind/m3)出现于乳山湾西流区海域,并向周围海域逐渐递减(见图3b)。

表1 乳山湾及附近海域桡足类种类组成Table 1 Species composition of copepods in Rushan Bay and adjacent sea area
从平面分布的季节变化上看,春季湾外的桡足类丰度要大于湾内;而秋季则相反,呈现湾内大于湾外的状况。随着季节变迁,湾内的桡足类丰度变化比湾外更小。经two-way ANOVA分析,各站位浮游动物丰度在季节和海域两者交互作用下差异显著。由此可见,丰度平面分布为综合因素影响的结果。

图2 海湾内外各站位种类数分布图Fig.2 The distribution of the number of species in the stations

图3 春、秋两季桡足类生物密度平面分布/ind·m-3Fig.3 The horizontal distribution of copepods,abundance in spring and autumn
2.3 桡足类多样性指数和分布

图4 春、秋两季桡足类香农威纳指数(H′)平面分布Fig.4 The distribution of Shannon-Wiener index(H′)in spring and in autumn
本次调查中,春季航次桡足类香农威纳多样性指数H′在1.532~2.803之间,平均值为2.156,其中湾内为2.297,湾外为2.061。相比于湾外,湾内桡足类多样性水平较高(H′>2.2),H′值的分布呈现从湾内逐渐向外海递减的趋势(见图4a)。均匀度指数范围在0.443~0.782之间,以站位A5最高,A11站位最低,丰富度指数0.919~1.555之间,C2站位最高,C8站位最低(见图5a)。秋季航次桡足类的香农威纳多样性指数H′平均值为2.122,其中湾内为1.887,湾外2.278。香农威纳指数H′值的波动范围为1.364~2.667,在湾内,桡足类多样性水平普遍较低(H′<2),且分布无明显规律,而湾外的多样性水平相对较高(H′>2),并随着远离近岸而逐渐增大(见图4b)。均匀度指数在0.455~0.751之间,以A1站位达到最高,C6站位最低。丰富度指数在0.555~1.506之间波动,最高值和最低值分别为A12站位和C7站位(见图5b)。

图5 春、秋两季各站位多样性指数分布Fig.5 Diversity indices of copepods in spring and in autumn among different stations
整体上看,春季航次桡足类香农威纳指数H′的分布表明湾内的桡足类群落多样性高于湾外,而秋季航次正好相反,湾外桡足类群落的多样性要高于湾内。经two-way ANOVA检验,香农威纳指数H′在季节和海域两因素交互作用下差异显著,这一点与丰度的检验结果相类似。
2.4 优势种及其分布
调查海域桡足类优势种见表2,在春季航次中,小拟哲水蚤(Paracalanus parvus)、双刺纺锤水蚤(Acartia bifilosa)和克氏纺锤水蚤(A.clausi)成为优势种。整体上看,春季全部优势种的高值区都出现在外海西部区域,其平面分布都呈现以湾口为低值中心向湾内和湾外递增的趋势(见图6)。
在秋季航次中,优势种有小拟哲水蚤、强额拟哲水蚤(P.crassirostris)、双刺纺锤水蚤和拟长腹剑水蚤(Oithona similis)。从平面分布上看,小拟哲水蚤在湾内丰度较低,且分布无明显规律,在湾外,该种类的丰度呈现近岸向外海逐渐递增的趋势。而强额拟哲水蚤、双刺纺锤水蚤和拟长腹剑水蚤的聚集区均在乳山湾西流区,其平面分布也均表现出从乳山湾西流区向周围逐渐递减的趋势,这与秋季桡足类总丰度的分布状况相一致(见图7)。

表2 调查海域各季节桡足类优势种Table 2 The dominant species of copepods in every season

图6 春季桡足类优势种丰度的平面分布/ind·m-3Fig.6 Distribution of abundances of the dominant copepods species in spring

图7 秋季桡足类优势种丰度的平面分布/ind·m-3Fig.7 Distribution of abundances of the dominant copepods species in autumn
2.5 环境因子与桡足类群落多样性的相关性
乳山湾及邻近海域的环境因子分布状况见图8,其中水体温度在春季呈现从外海逐渐向湾内递增的趋势,而秋季表现为从湾内向外海逐渐递增。在盐度方面,春季调查海域盐度相差较小,分布较为均衡,而秋季的盐度差距较大,最低值出现在西流区附近,整体呈现从湾内向湾外逐渐递增的趋势。春、秋两季调查海域浮游植物密度分布不均匀,整体呈现出湾内大于湾外的的状况,并且春季浮游植物密度要高于秋季。
桡足类香农威纳多样性指数H′与环境因素的相关性分析结果见表3:在春季航次,香农威纳指数H′与盐度呈现极显著的负相关,而水温和浮游植物与香农威纳指数H′的相关性不甚明显;而在秋季航次,香农威纳指数H′则与水温和盐度都呈现出极其显著的正相关,与浮游植物密度相关性不甚明显。
3 讨论
3.1 乳山湾及附近海域桡足类种类生态特征及与邻近海域差异比较
调查海域桡足类的生态类群可分为4大类,第一大类是以真刺唇角水蚤(Labibocera euchaeta)、背针胸刺水蚤(Centropages dorsispinatus)为代表的暖温带近岸低盐种,其次是如小拟哲水蚤、拟长腹剑水蚤等广温广盐性种类,此外还包括了以中华哲水蚤为代表的外海种和以细巧华哲水蚤(Sinocalanus tenellus)、火腿伪镖水蚤(Schmackeria poplesia)为代表的河口种等。整体上看,该海域桡足类的结构较为简单,是典型的温带近岸群落类型。
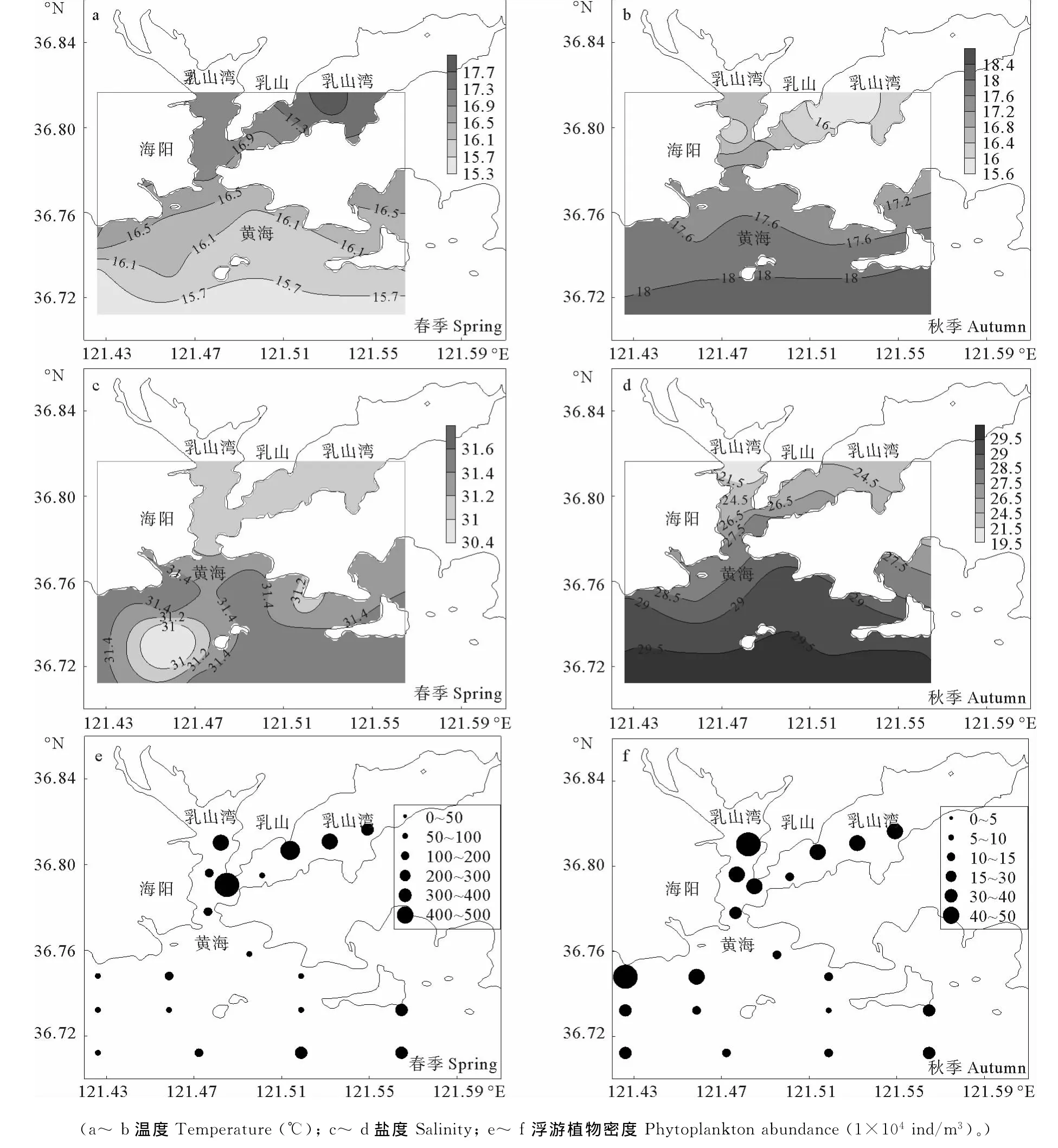
图8 乳山湾及邻近海域同步环境因素分布图Fig.8 Distribution of the environmental factors in Rushan Bay and adjacent sea area

表3 环境因子与桡足类多样性相关性分析Table 3 The correlation between environmental factors and the diversity of copepods community
调查共鉴定出桡足类28种,对比黄凤鹏和寇建山等在附近海区的调查研究[21-22],发现调查海域桡足类种类数与邻近海域相近,并且种类也具有相似性,均为南黄海常见种。然而,与上述邻近海区研究结果所不相同的是,本次调查中所采集到的桡足类多为小型种类,其丰度在整个海域更是具有绝对优势,像中华哲水蚤(Calanus sinicus)等大中型桡足类在种类和数量上均处于劣势。研究表明,小型桡足类兴盛是水体中浮游植物趋于小型化的必然结果[23],同步的浮游植物群落调查结果也与这一观点相吻合。同时,调查海域附近大量的养殖区也对此造成了影响。王岩、闫启伦等证实由于养殖生物的一系列摄食选择作用,使得养殖水体的浮游动物多以小型种类为主[24-25]。另外,选取采样的工具不同,调查结果也会产生差异。为更加全面地研究小型桡足类在近海浮游动物群落中的地位和作用,本次研究中桡足类丰度的计算以浅水Ⅱ型浮游生物网的调查结果为主,浅水Ⅰ型浮游生物网的调查结果仅作为补充。这是因为在大中型浮游动物的捕获效率差别极小的情况下,浅水Ⅱ型的调查结果在小型浮游动物种群数量上的估计更加准确[26-27]。
3.2 桡足类的时空分布特征
整体上看,调查海域的桡足类在分布上呈现季节和海域位置上的差异。首先,无论在种类组成、数量波动以及优势种类等方面,桡足类春秋2个季节都呈现出明显差异。这是因为对于南黄海近岸水体的桡足类而言,夏季成为群落变化的一个重要时期。一系列环境因素的变化使得如中华哲水蚤、克氏纺锤水蚤等部分桡足类被认为无法适应夏季该海域的生存环境,其种群数量减少甚至难以捕获[28-29],直至秋季水体环境适合后,其种群数量才逐渐恢复。然而,一些种类适应能力较强,在水体环境发生较大变化的时候也能存活,从而在春季和秋季都成为优势种类[30-31],本次研究中的小拟哲水蚤和双刺纺锤水蚤便是典型代表。另外,本次研究中拟长腹剑水蚤、强额拟哲水蚤等种类在春季原本不处优势地位,但在秋季却能占据优势地位。这是由于广温广盐性种类的适应能力较强,在度过夏季以后能够继续大量繁殖,从而占据由于部分大型桡足类的衰退而带来的生态位[32]。
然而,季节变迁带来桡足类群落的变化程度却也与其海域相关。调查结果发现,本次研究中包括细巧华哲水蚤、海洋伪镖水蚤(Pseudodiaptomus marinus)和火腿伪镖水蚤等的河口种类只在春季航次的湾内海域出现,在秋季航次,该类群在整个海域都难以寻觅。与此同时,包括背针胸刺水蚤等部分种类也仅仅出现在秋季航次的湾外海域。另外,从春季到秋季的过程中,湾内各站位的种类数也普遍下降,而湾外各站位种类数变化则相对较小。同时,春秋两季乳山湾湾内桡足类丰度的均值变化要小于湾外,而这最终导致了调查海域香农威纳多样性指数H′在2个季节中与丰度呈现不同分布特征。研究认为,半封闭的海湾向来都被认为是高丰度的区域[33],海水交换的限制带来的是相对稳定的生存环境,因而湾内种群数量得到保持,而多样性会相对减少。而外海的生物群落会有更高的几率得到由潮流运输作用带来新的种类[34]。因此随着季节变化,外海生物多样性水平将会得到保持,而半封闭的海湾反而会出现丰度保持稳定而种类数下降的状况。
3.3 桡足类群落与环境影响因素
调查海域隶属于山东半岛南部近岸水体,湾外受渤海低盐沿岸流[18]的支配,湾内受到陆地径流输入的强烈影响,因此,该海域的水团主要为湾内径流与外海水混合的咸淡水和湾外南黄海陆架水,整体呈现低盐环境[19-20]。对于半封闭海湾及邻近海域而言,季节和地域双重作用造就了环境因素的差异,这也成为本次研究中桡足类群落结构时空差异的直接原因[35-36]。
温度对桡足类的存活和分布具有重要影响,而温度的变化却与季节息息相关[37-38]。本次研究中春季和秋季温度大体相近,但桡足类多样性指数H′只在秋季与温度呈现显著的负相关。一般来说,南黄海区域的桡足类多能适应春、秋、冬3个季节的水温,因此,春季环境的温度不会限制桡足类的发展。然而,当夏季来临,一些桡足类被证明无法适应夏季水温升高带来的变化而进入消退期。如中华哲水蚤的最适水温一般在20℃以下[39],因而在夏季调查中难以寻觅。当进入秋冬季节时,水温开始回落,桡足类开始大量繁殖,此时随着水温逐渐下降至适合该类桡足类生存的温度,桡足类的繁殖也将达到更高的水平,其种类和数量也会逐渐增多。因此,在秋季,水体温度的回落对桡足类的分布有重要影响。
同时,研究表明,对于狭长型海湾地区而言,盐度是影响桡足类分布的主要因素[40]。研究中2个季节桡足类多样性指数H′与盐度均显著相关,表明盐度对于该海域桡足类分布十分关键。有学者证实,超过桡足类适应的盐度范围,桡足类的渗透压就会发生变化,生存就会受到制约,存活率逐渐下降[41],从而导致桡足类整个群落发生变化。在调查海域,湾外的海水要受到低盐度渤海沿岸流的强烈控制,湾内要受到径流输入的影响。无论湾内还是湾外,盐度都是水体生物重要的制约因素。在春季航次,海湾内外的盐度差距较小,这与该季节调查中海湾内外桡足类种类数相差较小相一致。而在秋季航次,盐度在海湾内外差距较大,并呈现典型的河口式盐度梯度分布趋势。同时,该季节盐度最低值出现在湾内西流区,这可能与乳山河的河水注入有关。该区域的环境正好符合低盐种类的生存环境,因此这也成为导致乳山湾西流区成为秋季桡足类密集区域的重要原因。Badylak认为,当盐度变化和桡足类的分布状况具有一致性时,盐度将成为影响桡足类分布的重要因素[42]。而本研究的结果也与之相一致,因此,盐度对于调查海域桡足类的分布起到了关键作用。
同步调查数据显示出乳山湾内浮游植物密度为春季湾内>春季湾外>秋季湾内>秋季湾外。在乳山湾内,浮游植物的密度始终都大于湾外,这可能也是导致桡足类能够在湾内保持数量相对稳定的重要原因。浮游植物是桡足类食物的重要组成部分,而食物只有短缺时才会制约整个动物群落发展[43-45],因此在秋季,外海区域桡足类丰度大量减少很可能与食物的减少有关。值得一提的是,研究表明,春季藻华使得大中型桡足类大量繁殖,夏秋季节浮游植物数量减少,小型桡足类将取代大型桡足类的地位[46]。而这与本次研究中拟长腹剑水蚤等在秋季的优势地位相吻合。然而,决定浮游动物对于浮游植物的摄食策略的关键是浮游植物的营养元素含量而不是密度[47]。而浮游植物也会通过其他机制[48-49]影响桡足类的种类和数量分布。同时,浮游植物本身也会受到温度和盐度等环境因素的影响而产生波动。因次,浮游植物密度与桡足类多样性之间的关系具有复杂性。因此环境因素对于桡足类的影响是交互的,应进一步结合实验室内生理生态研究和野外调查的方法来共同研究,其具体机制还有待深入探讨。
4 结论
(1)本次调查共采集到桡足类28种,其中春季22种,秋季20种,湾内种类数的变化要大于湾外。
(2)本次调查采集到的桡足类以近岸低盐种为主,兼有广温广盐种、少量的外海高盐种和河口半咸水种。
(3)春季桡足类优势种为小拟哲水蚤、克氏纺锤水蚤、双刺纺锤水蚤等,秋季为小拟哲水蚤、双刺纺锤水蚤、拟长腹剑水蚤和强额拟哲水蚤等。
(4)调查海域桡足类平面分布呈现出季节性和地域性的双重特点。不同季节,海湾内外的水文条件也不相同,而这也导致了桡足类时空分布的差异。
[1] 陈聚法,马绍赛,周诗赉,等.乳山湾东流区水文环境变化特征[J].海洋水产研究,1997,18:61-67.
[2] 王迪迪,孙耀,石晓勇,等.乳山湾东流区沉积物中氮形态的分布特征[J].海洋环境科学,2009,28:639-642.
[3] 冉祥滨,臧家业,韦钦胜,等.乳山湾口及其邻近海域溶解氧分布特征及影响因素研究[J].海洋学报,2011,4:173-180.
[4] 辛福言,陈碧鹃,曲克明,等.乳山湾表层海水COD与氮、磷营养盐的分布及其营养状况[J].海洋水产研究,2004,5:52-56.
[5] Longhurst A R.Relationship between diversity and the vertical structure of the upper ocean[J].Deep-Sea Res,1985,32:1535-1570.
[6] Enric S A,Calbet D,Atienza M A.Feeding and production of zooplankton in the Catalan Sea(NW Mediterranean)[J].Prog Oceanogr,2007,74:313-328.
[7] 郑重,张松踪,李松,等.中国海洋浮游桡足类(上卷)[M].上海:上海科学技术出版社,1965:1-2.
[8] 林茂,王春光,王彦国,等.西太平洋浮游动物种类多样性[J].生物多样性,2011,19(6):646-654.
[9] 唐启生,苏纪兰,孙松,等.中国近海生态动力学研究进展[J].地球科学进展,2005,20(12):1288-1289.
[10] Roff J C,Turner J T,Webber M K.Bacterivory by tropical copepod nauplii:extent and possible significance[J].Aquat Microb Ecol,1995,9:165-175.
[11] Bollens S M,Frost B W.Ovierity,selective predation,and variable diel vertical migration in Euchaeta elongate (Copepoda:calanoida)[J].Oecologia,1991,87:155-161.
[12] 徐兆礼.东海普通波水蚤种群特征与环境关系研究[J].应用生态学报,2006,17(1):107-112.
[13] Akithiro Shiomoto,Hiroki Asami.high-west and low-east distribution of chlorophyll a,primary productivity and diatoms in the subarctic North Pacific surface waters,midwinter 1996[J].J Ocanogr,1999,55:493-503.
[14] Calbet A,Landry M R,Scheinberg R D.Copepod grazing in a subtropical bay:species-specific responses to a mid-summer increase in nanoplankton standing stock [J].Mar Ecol Prog Ser,2000,193:75-84.
[15] Sieburth J M,Smetaeek V,Lenz J.Pelagic ecosystem structure:heterotrophic compartments of the plankton and their relationship to plankton size fraction[J].Limnol Oceanogr,1978,23:1256-1263.
[16] 徐兆礼,陈亚瞿.东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J].生态学杂志,1989,8(4):13-15.
[17] 刘红,袁兴中,张承德.山东曲阜地区蚂蚁群落结构及物种多样性研究[J].生物多样性,2002,10(3):298-304.
[18] 黄凤鹏,黄景洲,杨玉玲,等.胶州湾浮游桡足类时空分布[J].生态学报,2009,29(8):4045-4052.
[19] 寇建山,朱丽岩,戚本金,等.青岛南部近海浮游桡足类群落特征与影响因子[J].中国海洋大学学报:自然科学版,2011,4(10):52-60.
[20] 张才学,龚玉艳,王学锋,等.湛江港湾浮游桡足类群落结构的季节变化和影响因素[J].生态学报,2011,23(31):7086-7096.
[21] 王岩,张鸿淹.不同单养的混养海水实验围隔浮游生物的研究[J].中国水产科学,1999,6(3):49-54.
[22] 闫启伦,郭皓,王真良,等.浮游动物在贝类筏式养殖区内、外生态特征研究[J].黄渤海海洋,1999,17(1):46-50.
[23] 朱艺峰,王银,林霞,等.象山港两种网目网采浮游动物群落比较[J].应用生态学报,2012,23(8):2277-2286.
[24] 王荣,王克.两种浮游生物网捕获性能的现场测试[J].水产学报,2003,27(增刊):98-102.
[25] 王世伟.黄海中华哲水蚤繁殖、种群补充与生活史研究[D].青岛:中国科学院海洋研究所,2009.
[26] 黄世玫.胶州湾的浮游动物[J].山东海洋学院学报,1983,2(13):43-60.
[27] 张展.双刺纺锤水蚤Acartia bifilosa基础生物学与生态学研究[D].青岛:中国科学院海洋研究所,2006.
[28] Viitasalo M.Calanoid resting eggs in the Baltic Sea:implications for the population dynamics of Acartia bifilosa (Copepoda)[J].Mar Biol,1992,114:397-405.
[29] 霍元子.黄海浮游动物功能群的研究[D].青岛:中国科学院海洋研究所,2008.
[30] Archambault P,Roff J C,Bourget E,et al.Nearshore abundance of zooplankton in relation to coastal topographical heterogeneity and mechanisms involved [J].J Plankton Res,1998,20:671-690.
[31] Tse P,Hui S Y,Wong C K.Species composition and seasonal abundance of Chaetognatha in the subtropical coastal waters of Hong Kong[J].Estuar Coast Shelf S,2007,73:290-298.
[32] Rawlison K A,Davenport J,Barnes D K A,Tidal exchange of zooplankton between Lough Hyne and the adjacent coast[J].Estuar Coast Shelf S,2005,62:205-215.
[33] Archambault P,Roff J C,Bourget E,et al.Nearshore abundance of zooplankton in relation to coastal topographical heterogeneity and mechanisms involved [J].Journal of Plankton Research,1998,20:671-690.
[34] Webber M K,Roff J C.Annual biomass and production of the oceanic copepod community off Discovery Bay,Jamaica[J].Mar Biol,1995,123:481-495.
[35] 郑执中.黄海和东海西部浮游动物群落的结构及其季节变化[J].海洋与湖沼,1965,7(3):199-204.
[36] 苏育嵩.第二章:黄东海地理环境概况、环流系统与中心渔场[J].山东海洋学报,1986,16(1):16-27.
[37] 韦钦胜,于志刚,冉祥斌,等.黄海西部沿岸流系特征分析及其对物质输运的影响[J].地球科学进展,2011,26(2):145-156.
[38] 黄加琪,郑重.温度和盐度对厦门港几种桡足类存活率的影响[J].海洋与湖沼,1986,17(2):161-167.
[39] 张芳,孙松.中华哲水蚤生态学研究进展[J].海洋科学进展,2001,25(11):16-19.
[40] Marques S C,Pardal M A,Pereira M J.Zooplankton distribution and dynamic in a temperate shallow estuary[J].Hydrobiologia,2007,587:213-223.
[41] 黄加琪,郑重.九龙江口桡足类和盐度关系的初步研究[J].厦门大学学报,1984,23(4):497-505.
[42] Badylak S,Phlips E J.Spatial and temporal distributions of zooplankton in Tampa Bay,Florida,including observations during a HAB event[J].J Plankton Res,2008,30:449-465.
[43] Steele J H.The role of predation in ecosystem models[J].Mar Biol,1976,35:9-11.
[44] Kleppel G S.On the diets of calanoid [J].Mar Ecol Prog Ser,1993,99:183-195.
[45] Fessenden L M,Cowles T J.Copepod predation on phagotrophic ciliates in Oregon coastal waters[J].Mar Ecol Prog Ser,1994,107:103-111.
[46] 李超伦,王克.植食性浮游桡足类设施生态学研究进展[J].生态学报,2002,22(4):593-596.
[47] 苏强.浮游动物化学计量学稳态性特征研究进展[J].生态学报,2012,32(22):7213-7219.
[48] 杨位迪,杨青,林元烧,等.厦门港表层水体磷周转的生物学过程Ⅱ:桡足类对浮游植物的摄食压力[J].应用生态学报,2008,19(12):2774-2778.
[49] 韩刚.东海大规模赤潮对中华哲水蚤和黑褐新糠虾的生态毒理学研究[D].青岛:中国科学院海洋研究所,2006.
猜你喜欢
文化产业(2022年36期)2023-01-10 01:12:48
今日农业(2021年7期)2021-11-27 13:44:48
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
海洋通报(2020年5期)2021-01-14 09:27:02
河北渔业(2019年1期)2019-02-13 01:35:34
河南科技(2015年21期)2015-03-26 21:21:21
湖南农业(2015年7期)2015-03-20 01:48:37
海洋与湖沼(2014年4期)2014-03-09 06:57:32
