
地被菊‘火焰’对干旱胁迫的响应研究
2014-09-16 06:08赵夏陆任磊梁丽宝赵丽君王金将王晨赵靓吕晋慧张春来
山西农业大学学报(自然科学版) 2014年5期
赵夏陆,任磊,梁丽宝,赵丽君,王金将,王晨,赵靓,吕晋慧,张春来
(1.山西农业大学 林学院,山西 太谷 030801;2.山西省农业科学院高粱研究所 山西省高粱工程技术研究中心,山西 榆次 030600)
地被菊‘火焰’对干旱胁迫的响应研究
赵夏陆1,任磊1,梁丽宝1,赵丽君1,王金将1,王晨1,赵靓1,吕晋慧1,张春来2
(1.山西农业大学 林学院,山西 太谷 030801;2.山西省农业科学院高粱研究所 山西省高粱工程技术研究中心,山西 榆次 030600)
为探讨地被菊‘火焰’在干旱胁迫下的响应机制,采用盆栽控水法,研究不同干旱胁迫强度下地被菊‘火焰’生长、生理生化和光合生理的变化。结果表明:(1)干旱胁迫抑制‘火焰’生长发育,其中,中度和重度胁迫对植株生长影响较大。(2)随着干旱胁迫强度加剧,叶片相对含水量(LRWC)降低,叶片脯氨酸(Pro)、可溶性糖(Ss)和可溶性蛋白(Sp)含量增加,丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性和叶绿素(Chl)含量升高。(3)光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)随干旱胁迫程度加大而降低,胞间CO2浓度(Ci)呈先降后升趋势。结果表明土壤含水量为(13.33±0.2)%时对‘火焰’生长影响较小;土壤含水量为(10±0.2)%时,植株生长缓慢;土壤含水量为(6.66±0.2)%时‘火焰’生长明显受到抑制。
地被菊;干旱胁迫;生长;生理;光合生理
随着城镇绿地生态环境建设的快速发展,绿地面积和灌溉用水不断增加。在水资源短缺和季节性干旱地区,选择具有一定抗旱性、耐粗放管理的观赏地被植物作为绿化材料,对节约淡水资源和建设节约型园林具有重要意义。地被菊(ground-cover chrysanthemum)属菊科宿根草本植物,是20世纪80年代由陈俊愉等经过多年育种研究,选育出的菊花新品种群[1,2],具有开花繁密、群体花期长、抗逆性强、耐粗放管理等优良特性[3,4],适合在“三北”地区露地大面积应用,有较高的发展潜力和推广价值。‘火焰’为山西农业大学选育的优良地被菊品种,该品种株型紧凑、开花繁茂、花色艳丽,对其进行抗旱性研究,有助于‘火焰’的推广应用。
有关地被菊抗旱性研究已有部分报道,李锦馨[4,5]研究了在地栽控水条件下不同地被菊品种株高、冠幅、干物质含量、根冠比等的变化,认为地被菊抗旱性较强。时丽冉等[6,7]采用盆栽控水法模拟干旱,研究了地被菊品种‘落金钱’在不同程度干旱胁迫下光合生理特性和水分利用率以及膜脂过氧化和抗氧化酶活性的变化。目前国内研究多见于水分胁迫对地被菊生长、生理或光合某一方面的影响,未见综合地被菊生长形态、生理生化和光合特性对其抗旱机理进行研究,且不同地被菊品种抗旱性存在一定差异。为此,本文以地被菊品种‘火焰’为材料,测定其在不同干旱胁迫强度下生长、生理生化和光合生理的变化,探讨‘火焰’对干旱胁迫的响应机制,为‘火焰’在园林绿化中的合理应用提供科学依据。
1 材料与方法
1.1 材料和处理
供试材料‘火焰’,株高61 cm,冠幅90 cm;花序直径5~5.5 cm,舌状花6~7轮,舌状花正面为橙黄色,背面为红色;花期9月下旬至10月下旬。
试验于2013年5~9月在山西农业大学林学院温室进行,温度和湿度可控。2013年5月上旬于沙床扦插育苗,待插穗生根并展开10片叶后,挑选生长健壮、长势一致的植株于7月初定植于塑料盆中,每盆装基质(壤土∶河沙=2∶1)3 kg。缓苗2周后进行干旱胁迫处理。经预试验确定4个土壤水分梯度:
1)对照(CK),土壤含水量(16.66±0.2)%;
2)轻度干旱(LD),土壤含水量(13.33±0.2)%;
3)中度干旱(MD),土壤含水量(10±0.2)%;
4)重度干旱(SD),土壤含水量(6.66±0.2)%。
每处理3次重复,每次重复6株植株。为使各处理在同一天达到设定梯度,首先对重度胁迫处理停止供水,之后每隔2 d对其它干旱胁迫处理依次停止供水。每天18:00采用天平称重法保持土壤含水量恒定。干旱胁迫25 d时进行指标测定。
1.2 测定项目和方法
1.2.1 形态指标的测定
分别于处理前、处理后25 d对植株株高、叶片数进行测定。处理后25 d挑选植株顶端下第3~5节叶片,用叶面积仪测定叶面积。
1.2.2 生理生化指标的测定
取植株顶端下第3~5节叶片,测定叶片相对含水量(LRWC)、脯氨酸(Pro)含量、可溶性糖(Ss)含量、可溶性蛋白(Sp)含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、叶绿素(Chl)含量等生理指标。其中,LRWC采用烘干称重法[8];MDA含量采用硫代巴比妥酸比色法[9];Pro含量采用磺基水杨酸提取法[8];Ss含量采用蒽酮比色法[8];Sp含量采用考马斯亮蓝G-250染色法[8];SOD活性采用氮蓝四唑(NBT)光还原法[9];Chl含量用丙酮研磨法[8]。
1.2.3 光合指标的测定
取植株顶端下第3~5节叶片,用LCPro-SD便携式智能光合仪测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞CO2浓度(Ci)。测定时采用红蓝光源,光量子通量密度设定为1200 μmol·m-2·s-1,CO2浓度为(380±10)μmol· mol-1。
1.3 数据处理
应用SPSS 17.0统计软件对试验数据进行方差分析,采用Duncan多重比较法进行显著性分析,图表数据均为3次重复的平均值。
2 结果与分析
2.1 干旱胁迫对‘火焰’生长发育的影响
由表1可见,干旱胁迫显著降低了‘火焰’株高增量(plt;0.05)。株高增量随干旱胁迫强度加大呈下降趋势,在LD、MD、SD胁迫下分别比对照下降了22.7%、65.5%和94.3%;随干旱胁迫程度加深,‘火焰’叶片数增量逐渐降低。其中,叶片数增量在LD胁迫下与对照差异不显著(pgt;0.05),在MD、SD胁迫下与对照差异极显著(plt;0.01),分别比对照下降了39.6%、76.1%。
叶面积随干旱胁迫加重呈先升后降趋势(表1)。其中,LD胁迫与对照差异不显著(pgt;0.05),MD、SD胁迫极显著降低了叶面积(plt;0.01)。
2.2 干旱胁迫对‘火焰’叶生理生化指标的影响
2.2.1 干旱胁迫对LRWC的影响
随干旱胁迫程度加重,LRWC呈逐渐降低趋势(图1)。其中,LRWC在LD胁迫下与对照差异不显著(pgt; 0.05),MD和SD胁迫处理显著降低了LRWC(plt;0.05)。
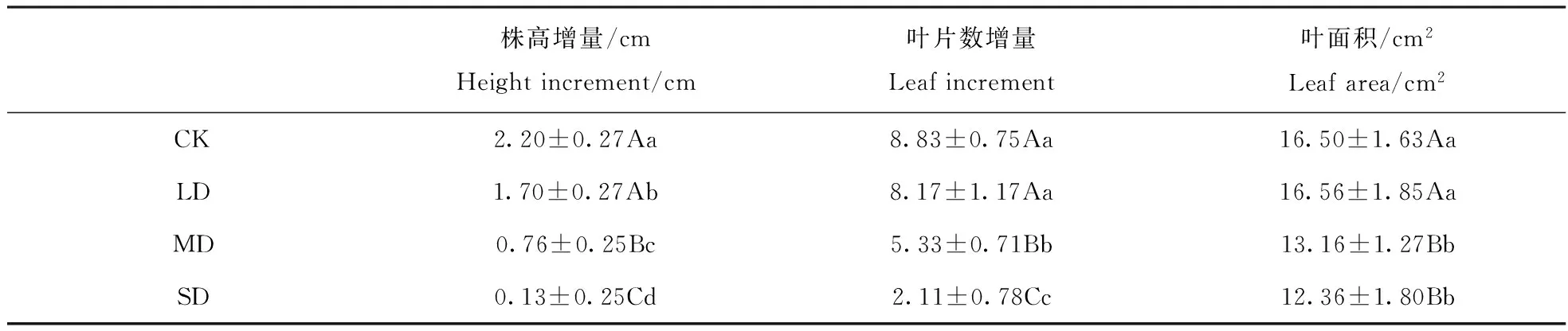
表1 干旱胁迫对‘火焰’生长发育的影响
注:不同大小写字母分别表示在plt;0.01和plt;0.05水平上差异显著。图1~图3同。
Note: Different uppercase letters and lowercase letters indicate significant difference atplt;0.01 andplt;0.05 levels, respectively. Similarly figure1 to figure 3.
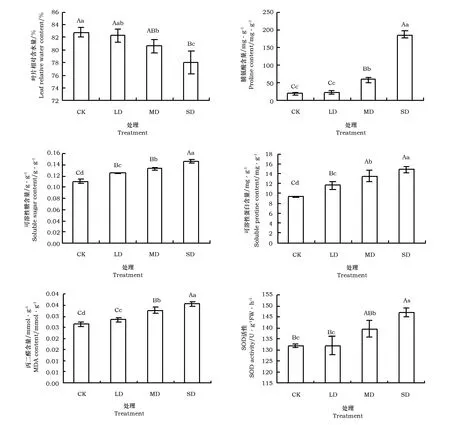
图1 干旱胁迫对叶片相对含水量、脯氨酸含量、可溶性糖含量、可溶性蛋白含量、丙二醛含量和超氧化物歧化酶(SOD)活性的影响
2.2.2 干旱胁迫对叶片渗透调节物质含量的影响
由图1可见,干旱胁迫极显著增加了叶片Pro含量(plt;0.01)。随干旱胁迫强度加大,‘火焰’叶片Pro含量持续上升。其中,Pro含量在MD、SD胁迫下增加幅度较大,分别是对照的2.9倍和9.3倍;叶片Ss含量随着干旱胁迫强度加剧逐渐增加,胁迫处理极显著地影响叶片Ss含量。在LD、MD、SD胁迫下叶片Ss含量分别比对照增加了13.4%、19.9%和32.5%;叶片Sp含量随干旱胁迫程度的加剧呈递增趋势,各处理间差异显著(plt;0.05)。
2.2.3 干旱胁迫对叶片MDA含量的影响
由图1可见,干旱胁迫显著增加了叶片MDA含量(plt;0.05)。随干旱胁迫程度加大,叶片MDA含量逐渐增加,在LD、MD、SD胁迫下分别比对照提高了7.8%、23.1%和34%。
2.2.4 干旱胁迫对叶片SOD活性的影响
随着干旱胁迫程度加剧,叶片SOD活性呈上升趋势(图1)。LD胁迫下,SOD活性与对照差异不显著(pgt;0.05),MD、SD胁迫处理显著提高了SOD活性(plt;0.05)。
2.3 干旱胁迫对‘火焰’叶光合响应特征的影响
2.3.1 干旱胁迫对‘火焰’Chl含量的影响
叶片Chl含量随干旱胁迫程度加重呈上升趋势(图2)。LD胁迫下,Chl含量与对照差异不显著(pgt;0.05),MD、SD胁迫极显著提高了Chl含量(plt;0.01)。
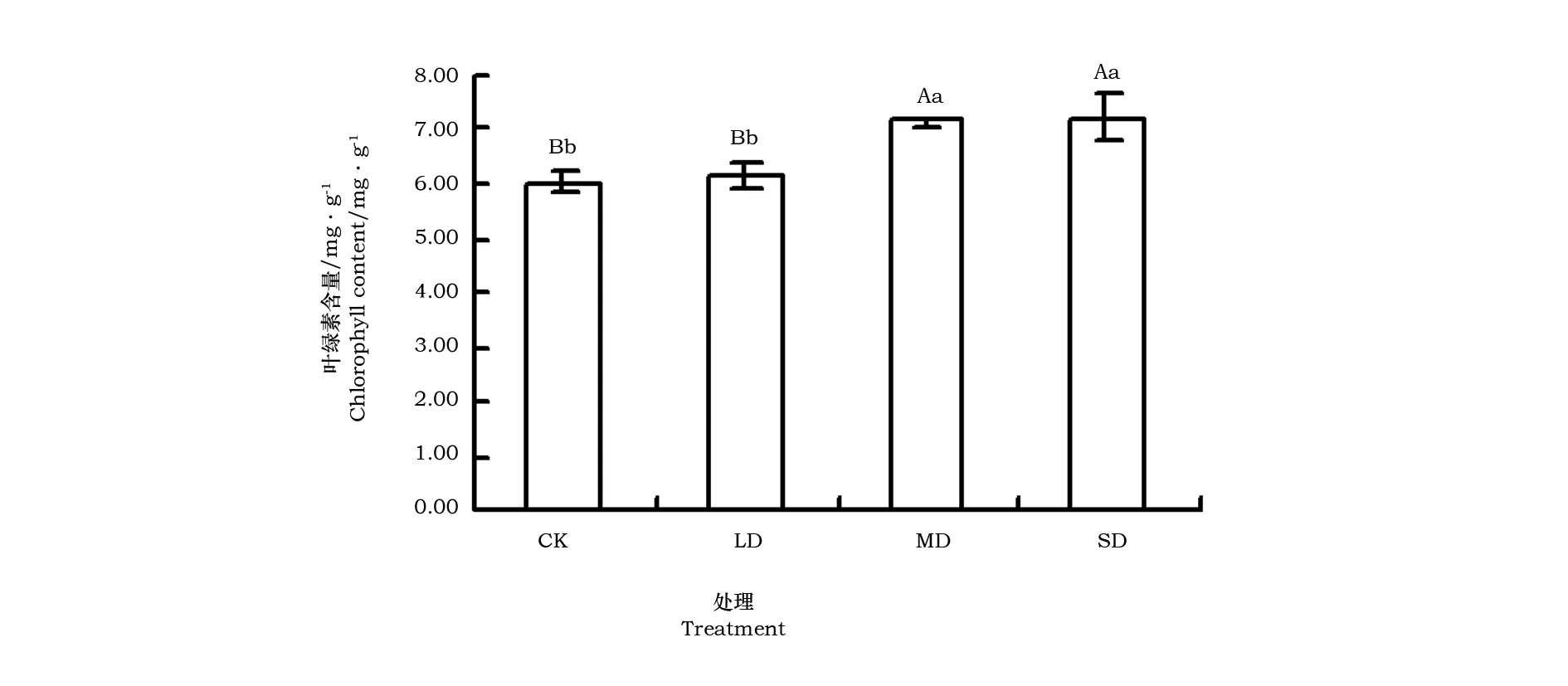
图2 干旱胁迫对叶绿素含量的影响
2.3.2 干旱胁迫对‘火焰’叶片Pn和Tr的影响
由图3可见,干旱胁迫极显著降低了叶片Pn(plt;0.01)。叶片Pn随干旱胁迫程度加大而下降,在LD、MD、SD干旱胁迫下分别比对照下降了16.2%、38%、61%。干旱胁迫极显著降低了叶片Tr(plt;0.01)。随着干旱胁迫强度加大,叶片Tr呈下降趋势,在LD、MD、SD胁迫下,Tr分别比对照下降了31.7%、53.6%和62.4%。
2.3.3 干旱胁迫对‘火焰’叶片Gs和Ci的影响
由图3可见,干旱胁迫极显著降低了‘火焰’Gs(plt;0.01)。Gs随干旱胁迫强度加大而下降,在LD、MD、SD胁迫下,Gs分别比对照下降了56.4%、71.8%和76.9%。干旱胁迫下Ci呈先降后升趋势。MD胁迫下,Ci降到最低,与对照差异显著(plt;0.05),SD胁迫下Ci开始上升,但与对照差异不显著(pgt;0.05)。
3 讨论
植物对干旱的反应不仅表现在生长发育、形态建成的适应性变化,同时体现在生理生化的适应性和调节性机制上[10,11]。水分调节能力是植物适应干旱环境的关键机制[12]。本研究中,轻度胁迫处理下,除株高增量与对照差异显著外,叶片数增量和叶面积均与对照差异不显著,说明轻度干旱对植株生长发育影响较小。在生理调节机制方面,‘火焰’Ss和Sp含量极显著增加,MDA显著增加,其它指标与对照差异不显著,表明在轻度干旱胁迫下,植物受到的伤害较轻。Ss和Sp含量增加对降低细胞水势、促进水分吸收和维持体内水分平衡有重要作用。在光合生理适应性方面,Pn、Gs和Ci显著下降,Tr呈下降趋势。许大全[13]认为,当Gs和Ci同时下降时,Pn下降是由气孔限制导致的,而判断光合作用非气孔限制的可靠依据是Ci升高和Gs降低。表明在轻度干旱条件下‘火焰’Pn降低主要是气孔限制引起的,气孔关闭的同时降低植株Tr,减少水分散失。
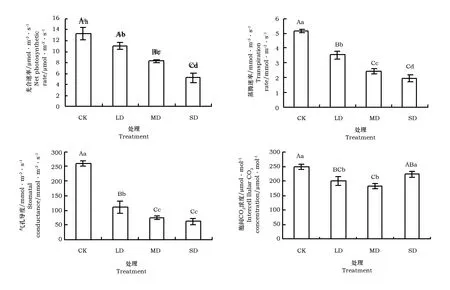
图3 干旱胁迫对光合速率、蒸腾速率、气孔导度和胞间CO2浓度的影响
水分胁迫抑制了细胞分裂和细胞伸长,阻碍了组织和器官的发育,使叶面积减少[14]。叶面积的减少使水分损失相应降低[14]。本研究中,中度干旱条件下,叶片数增量和叶面积极显著降低,植株通过减少叶片数和缩小叶面积来降低蒸腾。LRWC显著降低,MDA含量极显著上升,同时,渗透调节物质Pro、Ss和Sp含量持续增加,SOD活性升高,说明中度胁迫时植株体内水分亏缺已产生,细胞膜发生过氧化作用,且植株积极的做出适应性的调节反应。‘火焰’Ci降到最低,说明中度胁迫是Pn下降由气孔限制转变为非气孔限制的分界线。
重度胁迫时,植株株高生长基本停止,叶片数增量和叶面积进一步降低,植株生长受到抑制。王宇超等[15]研究表明当MDA含量大量增加时,细胞受到较严重的破坏。本研究中,重度干旱胁迫时LRWC极显著下降,MDA含量极显著升高,表明植株体内水分亏缺严重,细胞受到严重损伤。Pro、Ss和Sp含量持续上升。其中,Pro含量增加幅度远大于Ss和Sp,说明Pro在抵御重度干旱时起重要的渗透调节作用,与吴芹等[16,17]研究结果一致;樊卫国等[18]研究认为,重度胁迫时叶片Sp含量呈下降趋势,与本研究结果不一致,可能是重度干旱胁迫未超过植物所能忍耐的阈值,植物体内的可溶性蛋白合成能正常代谢[19]。‘火焰’SOD活性极显著升高。张仁和[20]认为,SOD活性在重度胁迫下呈降低趋势,与本研究结果不同,可能是‘火焰’对该逆境仍有一定的调节能力。Gs在重度胁迫时呈下降趋势,而Ci呈上升趋势,说明重度胁迫下‘火焰’Pn下降主要是非气孔限制引起的,但Ci与对照差异不显著,表明重度胁迫时光合系统未完全受到损伤,光合细胞还有一定的羧化能力。本研究中,在重度干旱条件下,‘火焰’Chl含量增加,与喻晓丽等[21]对火炬树的研究结果一致,可能与叶片含水量减小有关,这种变化对叶面积减小有补偿作用,是植株维持光合速率的生理适应机制[22]。
4 结论
(1)在轻度干旱条件下,植株通过调节Ss和Sp含量和气孔关闭来维持体内的水分平衡。
(2)中度干旱胁迫时,水分发生亏缺,植株受到伤害。一方面,植株通过增加Pro、Ss和Sp含量来降低细胞渗透势,促进水分吸收;另一方面植株通过调节气孔关闭和减缓生长来减少水分散失。
(3)重度干旱胁迫下,植株受伤害程度加重,生长受到抑制,但SOD活性和Pro、Ss和Sp含量呈上升趋势,且Ci与对照差异不显著,说明植株对SD胁迫有一定的适应能力。
(4)研究表明,土壤含水量为(13.33±0.2)%时对‘火焰’生长影响较小;土壤含水量为(10±0.2)%时,植株生长缓慢;土壤含水量为(6.66±0.2)%时‘火焰’生长明显受到抑制。
[1]崔娇鹏,陈俊愉.地被菊研究四十年及今后展望[C].中国菊花研究论文集(2002-2006),2002.
[2]王媛,崔雁汇,孔一昌,等.不同浓度赤霉素对地被菊‘紫重楼’开花特性的影响[J].中国农学通报,2012,28(19):164-167.
[3]吕晋慧,赵耀,王媛,等.地被菊花粉活力和储藏性研究[J].园艺学报,2012,39(12):2483-2490.
[4]李锦馨.地被菊地栽抗旱性试验研究[J].安徽农业科学,2008,36(36):15974-15976.
[5]李锦馨.地被菊在不同水分胁迫下生长状况研究[J].宁夏农林科技,2007(2):20-21.
[6]时丽冉,王玉平,刘国荣,等.干旱胁迫对地被菊光合生理特性及水分利用率的影响[J].河南农业科学,2011,40(3):119-121.
[7]时丽冉,陈红艳,崔兴国.干旱胁迫对地被菊膜脂过氧化和抗氧化酶活性的影响[J].北方园艺,2010(9):96-98.
[8]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[9]李合生,孙群,赵世杰,章文华.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[10]季孔庶,孙志勇,方彦.林木抗旱性研究进展[J].南京林业大学学报(自然科学版),2006,30(6):123-128.
[11]安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应[J].生态学报,2011,31(3):716-725.
[12]马成仓,高玉葆,李清芳,等.内蒙古高原锦鸡儿属植物的形态和生理生态适应性[J].生态学报,2011,31(21):6451-6459.
[13]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[14]李吉跃.植物耐旱性及其机理[J].北京林业大学学报,1991,13(3):92-100.
[15]王宇超,王得祥,彭少兵,等.干旱胁迫对木本滨藜生理特性的影响[J].林业科学,2010,46(1):61-67.
[16]吴芹,张光灿,裴斌,等.3个树种对不同程度土壤干旱的生理生化响应[J].生态学报,2013,33(12):3648-3656.
[17]刘旻霞,马建祖.6种植物在逆境胁迫下脯氨酸的累积特点研究[J].草业科学,2010,27(4):134-138.
[18]樊卫国,刘国琴,何嵩涛,等.刺梨对土壤干旱胁迫的生理响应[J].中国农业科学,2002,35(10):1243-1248.
[19]黄承玲,陈训,高贵龙.3种高山杜鹃对持续干旱的生理响应及抗旱性评价[J].林业科学,2011,47(6):48-55.
[20]张仁和,郑友军,马国胜,等.干旱胁迫对玉米苗期叶片光合作用和保护酶的影响[J].生态学报,2011,31(5):1303-1311.
[21]喻晓丽,邸雪颖,宋丽萍.水分胁迫对火炬树幼苗生长和生理特性的影响[J].林业科学,2007,43(11):57-61.
[22]李芳兰,包维楷,吴宁.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5406-5416.
ResponsestoDroughtStressinGround-coveringChrysanthemumcv'Huoyan'
Zhao Xialu1,Ren Lei1,Liang Libao1,Zhao Lijun1,Wang Jinjiang1,Wang Chen1,Zhao Jing1,Lv Jinhui1,Zhang Chunlai2
(1.CollegeofForestry,ShanxiAgriculturalUniversity,TaiguShanxi030801,China;2.InstituteforsorghumResearch,ShanxiAcademyofAgriculturalSciences/ShanxiProvincialEngineeringRDCentreforSorghum,YuciShanxi030600,China)
To provide optimized cultivation procedures for ground-covering chrysanthemum, this study investigated the growth and physiological responses of the chrysanthemum plant to drought stress. Using ground-covering chrysanthemum cultivar 'Huoyan' as material and pot-cultivation experiment with controlled irrigation, parameters of growth, stress physiology and biochemistry and photosynthetic physiology of 'Huoyan' plants under varied degree of drought stress were measured. The results showed that: drought stress inhibited plant growth, especially under moderate and severe drought stress conditions. Under increased degree of drought stress leaf relative water content (LRWC) was decreased, proline (Pro), contents of soluble sugar (Ss) and soluble protein (Sp) were increased, malonic aldehyde (MDA) content, the activity of superoxide dismutase (SOD) and chlorophyll content (Chl) were increased. Under drought stress, photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate (Tr) were significantly decreased while intercellular CO2concentration (Ci) was first increased and then decreased. When soil moisture content was 13.33%±0.2% little impact on plant growth was observed. When soil moisture dropped to 10%±0.2%, the plants grew slowly. When soil moisture reached 6.66%±0.2% plant growth was suppressed considerably.KeywordsGround-covering chrysanthemum; Drought stress; Plant growth; Physiological response; Photosynthetic physiology
2014-05-04
2014-06-11
赵夏陆(1989-),女(汉),山西离石人,在读硕士,研究方向:园林植物遗传育种。
吕晋慧,教授,硕士生导师。Tel:15935485943;E-mail: lujinhui11@126.com
“十二五”国家科技支撑计划课题(2013BAD01B07)
S682.1+1
A
1671-8151(2014)05-0458-06
(编辑:马荣博)
猜你喜欢
音乐天地(音乐创作版)(2022年1期)2022-04-26
学苑创造·A版(2021年2期)2021-03-11
世界科学(2020年1期)2020-02-11
中国生物医学工程学报(2019年5期)2019-07-16
动漫星空(兴趣百科)(2019年5期)2019-05-11
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
Coco薇(2017年5期)2017-06-05
学与玩(2017年6期)2017-02-16
电源技术(2016年9期)2016-02-27
