
线粒体转录因子A参与软骨细胞退变逆转的机制
2014-09-13 03:17梁月屏廖壮文吕浩然谢楚海
中国老年学杂志 2014年20期
黄 彦 梁月屏 廖壮文 吕浩然 谢楚海
(广州医科大学附属第二医院骨外科,广东 广州 510260)
骨关节炎(OA)是一种以关节软骨的变性、破坏及骨质增生为特征的慢性关节病〔1〕。关节软骨细胞的过度凋亡是OA发病的重要病理机制,而软骨细胞的凋亡与线粒体关系密切,线粒体转录因子A(TFAM)是线粒体转录和复制的关键激活子,与细胞凋亡过程相逆〔2〕。本研究分析膝关节骨关节炎患者、正常关节软骨细胞的线粒体DNA特征和TFAM表达的差异情况,以期阐明TFAM与OA发生、发展的关系。
1 资料与方法
1.1病例资料 选择我院2011年10月至2012年10月的膝关节OA患者20例,入选OA组,同时设立因关节矫形等手术剩余的正常关节软骨组织细胞20例。诊断标准:参照中华医学会骨科学分会制定的OA诊治指南(2007年版)的诊断标准〔3〕。纳入标准:①女性患者,绝经1 年以上, 年龄55~65岁; ②符合上述的诊断标准; ③能够收集到较为齐全的相关资料者并签署知情同意书;④研究前患者均签订知情同意书。排除标准:①继发性膝骨关节炎患者;②6个月内服用过影响骨代谢药物的患者;③合并严重心、肝、肾以及其他系统疾病者,精神病及肿瘤患者;④不同意和配合进行研究的患者。
1.2收集样本和建立软骨细胞的体外培养体系
1.2.1关节软骨的收集 取膝OA关节置换手术中取下关节软骨。取下的软骨组织置于含青霉素钠/链霉素双抗PBS液的无菌培养皿中,漂洗去除血细胞和关节滑液,清除滑膜组织。最后在含双抗液的PBS液的无菌培养皿中用眼科剪将软骨组织剪成1~3 mm大小的组织碎块,注意使标本保持湿润状态。
1.2.2软骨细胞的培养 无菌条件下取得标本后立即置入盛有PBS的无菌培养皿中,无菌生物柜中取材术中切除的关节全层软骨片(去除周围软组织),无菌手术刀片切块,剪碎,加入PBS液冲洗,将软骨移入离心管中,加入Ⅱ型胶原酶,37℃培养箱中消化,其中2 h后每隔20 min晃动离心管一次,至悬液变混浊确定为软骨块基本被消化。之后离心,弃上清液。加入完全培养基,充分吹打混匀后可获得含有软骨细胞的悬液。倒置显微镜观察软骨细胞培养各个时期的细胞形态,大小,分布。
1.2.3MTT比色法测细胞增殖 取对数生长期的两种软骨细胞,接种于96孔板,分别于培养24 h、48 h、72 h、96 h、120 h后,加入20 μl MTT孵育4 h后,再加入100 μl DMSO溶解形成的甲瓒晶体,振摇10 min,用酶标仪测定570 nm处的吸光度。
1.2.4流式细胞术检测细胞凋亡 采用PI/Annexin V法,参照BD公司凋亡检测试剂盒的说明书操作,将待检测的两种软骨细胞在染色液中(10 ml PI, 1 ml Annexin V-FITC, 10 ml上样缓冲液和79 ml H2O)室温孵育15 min后,应用用流式细胞仪在488 nm波长氩离子激光下以Elite软件分析细胞凋亡率。
1.3线粒体DNA特征与线粒体转录因子A表达
1.3.1PCR引物及TaqMan探针 本实验中所有引物由上海英潍捷基贸易有限公司合成、标记。应用荧光检测物质有报告基因FAM 和熄灭基因TAMRA。mtDNA D-loop HV1(Hipervariable region 1, HV1)上游引物5′-TTGCACGGTACCATAAATACTTGAC-3′,下游引物5′-GAGTTGCAGTTGATGTGTGATAGTTG-3′,TaqMan探针5′-CTCCCCATGCTTACAAGCAAGTACAGCAAT-3′,片段长度128 bp。核β-globin基因上游引物5′-ACACAACTGTGTTCACTAGC-3′,下游引物 5′-CAACTTCATCCACGTTCACC-3′,TaqMan探针5′-CTCCTGAG GAGA AGTCTGCT-3′,片段长度110 bp。TFAM上游引物5′-GGAATGTGGAGCGTGCTAAAA-3′,下游引物5′-TGCTGGAAAAACACTTCGGAATA-3′,片段长度118 bp。β-actin上游引物5′-TTCGAGCAAGAGATGGCCA-3′,下游引物 5′-TACATGGTGGTGCCGCC-3′,片段长度270 bp。
1.3.2细胞线粒体DNA的提取 加RNase(200 μg/ml)于37℃、2~4 h消化RNA。之后,用酚、酚氯仿、异戊醇(体积比为25∶24∶1) DNA以去除蛋白质、脂类等其他物质。纯化后的线粒体DNA量较少,线粒体DNA溶液加入0.6倍体积异丙醇或沉淀缓冲液,置于-20℃下放置30 min,进行离心即得纯净的线粒体DNA。
1.3.3线粒体DNA拷贝数的实时荧光定量PCR检测 以线粒体DNA HV1和核单拷贝基因β-globin分别作为线粒体和核DNA拷贝数的标记。设计HV1和β-globin基因的引物及探针,对该基因片段进行PCR扩增,在绘制标准曲线的基础之上,分别对待测样本线粒体HV1和β-globin基因进行实时荧光定量PCR检测。线粒体DNA的拷贝数计算公式:相对拷贝数/二倍体核基因组=2×HV1/β-globin。
1.3.4线粒体转录因子A表达的测定 RNA抽提及逆转录cDNA:按照RNA提取试剂盒说明书提取组织RNA。置于紫外分光光度计上波长为260 nm和280 nm处测量吸光值,计算两者的比值,选取比值在1.7~2.0者用于逆转录反应。cDNA合成严格按试剂盒说明书进行,合成的cDNA储存在-20℃用于PCR反应。定量PCR反应:加入上下游引物各,在7500 Real Time PCR仪监测TFAM的表达差异。产物鉴定:取RT-PCR产物加入buffer,经电泳后,溴乙锭染色。
1.4统计学处理 用SPSS16.0软件行方差分析及χ2检验。
2 结 果
2.1倒置显微镜观察 分离原代软骨细胞呈球形,大小均一。正常组原代软骨细胞呈圆形或椭圆形,贴壁较快,6 d后长满瓶壁,传代后增殖加快,随着传代次数增加,细胞由椭圆形变为梭形细胞。而OA组原代细胞贴壁晚,单层集落样,细胞形态多不规则,传代后细胞以梭形为主,贴壁缓慢,4 d左右长满瓶壁。
2.2MTT试验 正常组增殖在24、48、72、96、120 h均较OA组明显旺盛(P<0.01),而OA细胞组增殖较慢,到72 h后趋向平台期。见表1。
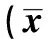
表1 实验各组MTT试验结果图±s)
2.3流式细胞术检测细胞凋亡 OA组的凋亡率高于正常细胞组(P<0.01),正常细胞凋亡率为1.65%,OA细胞凋亡率为7.11 %。2 μg/L IL-1β作用72 h后,正常细胞凋亡率为13.79%,OA细胞凋亡率为35.13 %。2 μg/L浓度的IL-1β对OA软骨细胞的凋亡诱导强于正常组(P<0.01)。
2.4线粒体DNA拷贝数和TFMA mRNA表达量的变化 HV1和β-globin基因定量PCR标准曲线的相关系数分别为0. 996和0. 998,斜率分别为-3.148和-3.369。OA组线粒体DNA的平均拷贝数为(389±117),而正常细胞为(756±185),前者显著低于后者(P<0.01)。
2.5TFMA mRNA表达量的变化及与线粒体DNA拷贝数的关系 定量RT-PCR结果显示,OA组TFMA mRNA的表达量(TFMA/β-actin)为0.556±0.021,而正常细胞表达量为1.053±0.024,前者低于后者(P<0.05)。见图1。表达量和线粒体DNA拷贝数之间有相关性(r=0.924,P<0.01);在OA组中,线粒体DNA拷贝数和TFMA mRNA的表达水平也有相关性(r=0.906,P<0.05)。

图1 各组TFMA mRNA表达
3 讨 论
软骨细胞、蛋白多糖和Ⅱ型胶原相互作用,共同维持软骨的生理性状,其中任何改变都可以引起软骨的退变〔4〕。大量研究发现软骨细胞凋亡的增加是OA病理改变的重要特征〔5〕,Johnson等〔6〕研究证实,无论是活体还是体外培养,OA软骨中存在过度的软骨细胞凋亡。Leong等〔7〕认为相比正常的软骨细胞,OA的软骨细胞所分泌的蛋白质发生改变,同时合成代谢活动减少、炎性细胞因子的增加和基质降解酶活跃,导致关节软骨的损伤。Musumeci等〔8〕研究发现,OA患者不仅软骨细胞的结构改变,较正常软骨细胞凋亡的增加,而且基质钙化和蛋白多糖减少。因此,在本研究选择软骨细胞作为研究OA的作用点,体外培养观察表明原代正常组细胞附壁生长和增殖活性良好,具有较好的软骨细胞生物学特性,而OA组软骨细胞的贴壁和增殖较慢,传代久后生物学特性基本消失,凋亡明显增多,对相同浓度IL-1β的诱导反应敏感,提示分离的原代OA软骨细胞符合软骨细胞退变,为其研究提供了较好的实验对象。
Musumeci等〔9〕的研究认为,线粒体的功能障碍是破坏了软骨细胞在关节内微环境,从而导致了骨关节炎的发生〔10〕。OA的软骨细胞线粒体功能障碍可能来自体细胞中的线粒体DNA突变或炎症介质的直接影响,使软骨基质钙化和软骨细胞凋亡的增加。线粒体的复制和转录过程需要核编码的一些转录因子进行调控,在哺乳动物中起主要调节作用的因子是TFAM〔11〕。TFAM是由596 bp核苷酸编码的25 kD蛋白,具有两个短串联的高亲和力的高迁移率族蛋白(HMG)盒域,在结构上包含一个N端头结构域、两个HMG box结构域和一个C末端尾结构, 两个HMG box之间还存在一个由27个氨基酸残基组成的短连接区域。Rebelo等〔12〕的研究认为,TFAM与线粒体DNA在线粒体中结合成复合体,从而保持线粒体DNA结构的稳定性。本研究结果发现,OA软骨细胞TFMA mRNA的表达量低于正常软骨细胞,说明OA软骨细胞中线粒体DNA拷贝数下降,调控其转录、复制的主要调节因子TFMA的表达也相应下调。Nakanishi等〔13〕研究后认为TFAM可以改善线粒体DNA的损伤和减少,由此产生的氧化还原调节炎症反应;Woo等〔14〕研究了敲除线粒体TFAM基因的小鼠,发现其线粒体DNA出现缺失,同时出现线粒体的氧化损伤。
在软骨细胞中线粒体转录因子A在线粒体DNA转录、复制、结构的维持和损伤修复及细胞凋亡中均起着重要作用。线粒体DNA自身遗传结构改变和线粒体转录因子A等因素共同作用是导致OA软骨细胞线粒体DNA拷贝数改变的原因。
4 参考文献
1Sinusas K. Osteoarthritis: diagnosis and treatment〔J〕. Am Fam Phys,2012; 85(1):49-56.
2Iliopoulos D, Gkretsi V, Tsezou A. Proteomics of osteoarthritic chondrocytes and cartilage〔J〕. Expert Rev Proteomics, 2010;7(5):749-60.
3中华医学会骨科学分会. 骨关节炎诊治指南(2007年版)〔J〕.中国临床医生,2008; 1: 28-30.
4Danisovic L, Varga I, Zamborsky R,etal. The tissue engineering of articular cartilage: cells, scaffolds and stimulating factors〔J〕. Exp Biol Med (Maywood), 2012; 237(1):10-7.
5Zamli Z, Sharif M. Chondrocyte apoptosis: a cause or consequence of osteoarthritis〔J〕?Int J Rheum Dis, 2011;14(2):159-66.
6Johnson EO, Charchandi A, Babis GC,etal. Apoptosis in osteoarthritis: morphology, mechanisms, and potential means for therapeutic intervention〔J〕. J Surg Orthop Adv, 2008;17(3):147-52.
7Leong DJ, Sun HB. Events in articular chondrocytes with aging〔J〕. Curr Osteoporos Rep, 2011; 9(4):196-201.
8Musumeci G, Loreto C, Carnazza ML,etal. Characterization of apoptosis in articular cartilage derived from the knee joints of patients with osteoarthritis〔J〕. Knee Surg Sports Traumatol Arthrosc, 2011; 19(2):307-13.
9Musumeci G, Loreto C, Carnazza ML,etal. OA cartilage derived chondrocytes encapsulated in poly(ethylene glycol) diacrylate (PEGDA) for the evaluation of cartilage restoration and apoptosis in an in vitro model〔J〕. Histol Histopathol, 2011; 26(10):1265-78.
10Blanco FJ, Rego I, Ruiz-Romero C. The role of mitochondria in osteoarthritis〔J〕. Nat Rev Rheumatol, 2011;7(3):161-9.
11Canugovi C, Maynard S, Bayne AC, The mitochondrial transcription factor A functions in mitochondrial base excision repair〔J〕. DNA Repair (Amst), 2010; 9(10):1080-9.
12Rebelo AP, Williams SL, Moraes CT. In vivo methylation of mtDNA reveals the dynamics of protein-mtDNA interactions〔J〕. Nucleic Acids Res, 2009;37(20):6701-15.
13Nakanishi H, Hayashi Y, Wu Z. The role of microglial mtDNA damage in age-dependent prolonged LPS-induced sickness behavior〔J〕. Neuron Glia Biol, 2011;28(1):1-7.
14Woo DK, Green PD, Santos JH,etal. Mitochondrial genome instability and ROS enhance intestinal tumorigenesis in APC(Min/+) mice〔J〕. Am J Pathol, 2012;180(1):24-31.
猜你喜欢
右江民族医学院学报(2022年2期)2022-05-19
中老年保健(2021年5期)2021-12-02
河北医学(2021年10期)2021-10-27
今日农业(2020年19期)2020-12-14
医学新知(2019年4期)2020-01-02
中国临床医学影像杂志(2019年6期)2019-08-27
中成药(2018年9期)2018-10-09
中国实验动物学报(2017年2期)2017-05-18
中国病理生理杂志(2017年2期)2017-01-17
中华实验和临床病毒学杂志(2016年3期)2016-08-09
