
草地生态系统对模拟大气增温的响应
2014-08-25 01:20黄文华王树彦焦志军韩国栋
草业科学 2014年11期
黄文华,王树彦,韩 冰,焦志军,韩国栋
(1.内蒙古农业大学生命科学学院,内蒙古 呼和浩特 010018; 2.内蒙古农业大学农学院,内蒙古 呼和浩特 010018)
草地生态系统对模拟大气增温的响应
黄文华1,王树彦2,韩 冰1,焦志军1,韩国栋1
(1.内蒙古农业大学生命科学学院,内蒙古 呼和浩特 010018; 2.内蒙古农业大学农学院,内蒙古 呼和浩特 010018)
气候变暖是全球气候变化的主要趋势,对生态系统将产生不可估量的影响。增温使植物的生长期延长,生物量有所增加,生长最适宜温度低的植物光合速率增加;而气孔导度变化没有规律,叶片中钠、钾、磷的含量显著下降。增温条件下不同草地类型植物群落的生物量的变化不一致,高寒草甸的矮嵩草(Kobresiahumilis)将会被其他替代,羊草草原会被大针茅(Stipagrandis)替代。被研究的草地动物主要是昆虫和鼠类,增温后昆虫世代数增加,并有向高海拔和高纬度地区扩散的趋势;增温样地鼠类的种群密度有所提高,性别比例和种群年龄结构未发生改变。增温使土壤呼吸和土壤酶活性增强,土壤细菌量增加,有机碳的变化不确定。本文总结了草地生态系统各组分对于增温的响应,但增温对草地生态系统和生物圈的影响有待于进一步深入持久的研究。
全球变暖;草地生态系统;植物;动物;土壤
全球变暖是21世纪全球气候变化的主要趋势[1]。监测和研究表明,由19世纪开始的温室效应正在继续和扩大,全球表面温度在过去的100年里上升了0.4~0.8 ℃,在高纬度和高海拔地区升幅更大[2]。根据政府间气候变化专门委员会(IPCC)第4次评估报告的预测,到2100年全球的平均气温将升高1.4~4.0 ℃[3]。
草地生态系统作为陆地生态系统的主体类型,占陆地总面积的1/4~1/3,对气候和环境的变化十分敏感[4]。中国草地面积约4亿hm2,约占国土总面积的42%,是我国最重要的自然资源。野外自然条件下生态系统增温试验是研究全球变暖与陆地生态系统关系的主要方法之一[5]。在模拟增温的条件下,我国学者对青藏高原及周缘地区的高寒草原、内蒙古典型草原、荒漠草原、东北草甸草原等不同草地生态系统的不同层面进行了研究。
生态系统包括生物部分和非生物部分。增温对植物的影响主要是通过改变植物的生长生理、物候特征,从而在群落层面改变生物量的生产、分配及生物的多样性等。增温对于草地生态系统动物的影响主要是对昆虫和鼠类的影响。土壤对增温的响应是从土壤呼吸、土壤酶、土壤有机碳、土壤养分的改变来实现的。本文综述草地生态系统对模拟大气增温的响应,并提出了在全球气候变化下模拟增温需要进一步深入研究的方向,为减少温室气体排放造成的气候变暖提供理论依据。
1 植物对大气增温的响应
1.1植物个体对大气增温的响应
植物是草地生态系统的主体,温度是影响植物生长和发育的重要生态因子之一。增温可能促进植物生长发育[6-7],也可能抑制其生长[8-9]或无显著影响[10](表1)。
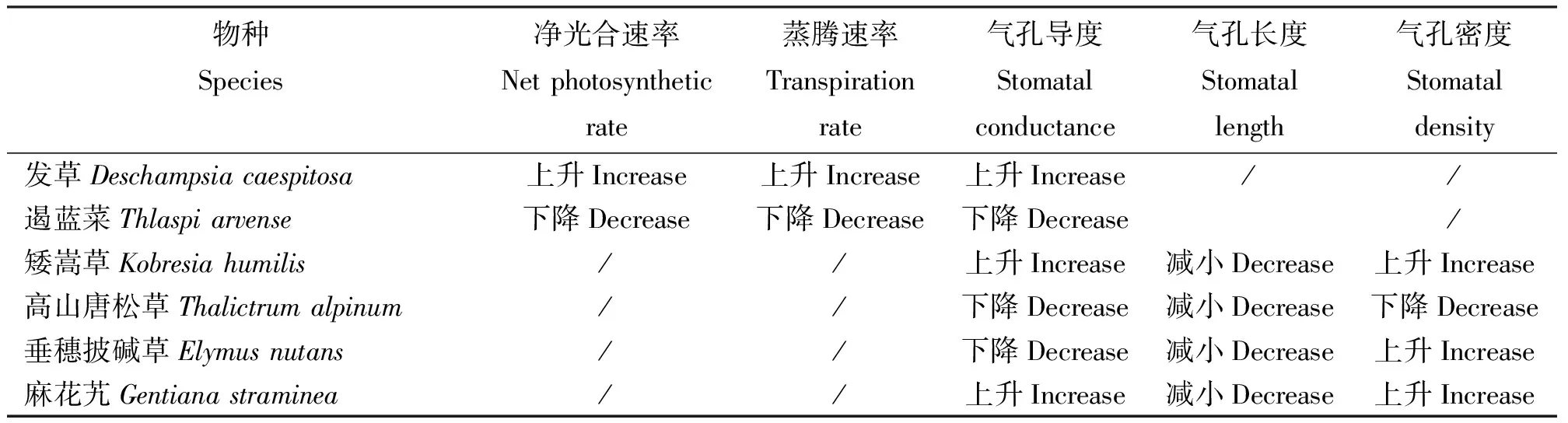
表1 植物个体生理指标对模拟增温的响应Table 1 The response of the plant individual physiological indexes of simulated warming
注:“/ ”表示目前没有研究结果,下同。
Note: “/” means no study result.The same below.
在野外自然条件下采用开顶式生长室模拟增温处理,发现单子叶草本植物发草(D.caespitosa)地上生物量增加,其光响应曲线明显高于对照处理,净光合速率、蒸腾速率、气孔导度、最大净光合速率等指标都明显上升;而双子叶植物遏蓝菜(T.arvense)的净光合速率、蒸腾速率和气孔导度在增温后明显下降[11]。赵建中等[12]对青藏高原矮嵩草草甸的莎草科黑褐苔草(Carexalrofusca)研究发现,其叶片数目随温度的升高而减少,分蘖数和生物量在一定的温度范围内达到最大值,温度过高或过低都不利于它们的生长和发育,认为莎草科植物存在最适宜生长温度。对高寒草甸重要植物矮嵩草(K.humilis)、高山唐松草(T.alpinum)、垂穗披碱草(E.nutans)和麻花艽(G.straminea)的研究发现,增温使矮嵩草、麻花艽和垂穗披碱草气孔密度上升,而高山唐松草下降;4种植物的气孔长度均减小;高山唐松草和垂穗披碱草气孔导度下降,矮嵩草和麻花艽上升[13]。一般来说,温度升高,减少高寒植物低温胁迫,促进有机质矿化[14],从而促进植物生长和CO2吸收增加,这时最大气孔导度增加更具适应意义。但是,对于光合最适温度较低的植物来说,温度增加后,达到光合最适温度的时间可能减少,不利于CO2吸收。这样,不同的植物对于增温的响应不同,其生理指标的变化也不同,但是各种变化总是朝着最适宜增温的方向进行。
一般情况下,增温能使植物物候进程加快,春季物候期提前,植物的秋季物候期推迟,植物生长季长度延长。矮嵩草草甸中的短穗兔耳草(Lagotisbrachystachya)在增温时营养生长被促进,生殖生长被抑制,同时表现出匍匐茎的数目随着增温而减少。增温能使矮嵩草草甸的各种群生长期延长,各物候的始期提前,而末期推迟[15]。Dunne等[16]利用辐射加热器改变环境温度研究美国科罗拉多亚高山草地,11种灌丛和草本花期对增温反应各有不同,有一些物种对增温的响应更为敏感,从而破坏种间竞争关系,引起群落优势种和组成发生改变。
羊草草原上,增温使羊草(Leymuschinensis)的生物量增加,分蘖和根的生物量增加,而鞘的生物量稍降低,对叶、根茎的影响较小。温差增大提高了分蘖和根的投资比例,而降低了其他器官的投资,表明夜间温度的降低将促进叶等源器官和鞘等暂储器官干物质的向外转移,增加对分蘖和根的投资,这是羊草对昼夜温差加大的适应性反应[17]。Regory等[18]通过转移积雪的方法提高土壤温度,研究增温对阿拉斯加草本植物的影响,结果表明增温没有使叶片大小和萌发数量发生变化。
对增温条件下松嫩草原羊草叶片矿质元素的影响进行研究,与对照相比,增温对叶片氮、钙离子、镁离子含量的影响差异不显著,却使磷含量显著降低了8.73%,钾离子含量显著降低了10.6%,钠离子含量显著降低了12.8%[19]。对羊草4大类12亚类植硅体的研究发现,增温后植硅体普遍增大,原因是增温促进了羊草的同化作用,吸收积累更多的硅元素。其中帽型植硅体中,尖顶帽型和平顶帽型的含量随着温度的升高而增加,刺帽型的含量下降。植硅体有规律的变化,说明其对外界的环境很敏感,能够反映环境因子的变化,作为古环境的代用指标是有效的[20]。由此可见,通过对植硅体的研究能够发现随着温度的升高,羊草对于硅元素的吸收是增加的,但是其他的矿质元素并没有特殊的规律。
温度低的区域,温度是植物生长的主要限制因子,温度高的区域,营养成分则是植物生长的主要限制因子。所以,温度作为重要的生态因子,对植物的生长和发育起着至关重要的作用,但是不同的草原类型,不同的增温装置、不同的植物种类对于增温的响应也不同。
1.2植物群落对增温的响应
温度变化影响了植物的生长发育,进而对植物的群落结构产生一定的影响。增温条件下对群落生物量的研究得到了不同的结果(表2)。
已有的研究表明,阿拉斯加苔原、英格兰北部草原在增温后生物量显著增加[21]。在未来温度升高2.6 ℃的条件下,美国矮草草原的生产力将增加[22]。Rebeeca等[23]在落基山脉用红外线设备照射控制环境温度,发现加热小区的三齿蒿(Artemisiatridentata)的地上生物量有所增加。对海北矮嵩草草甸连续5年的观测发现,植物群落地上部分生物量、平均高度、盖度均表现为逐渐上升的趋势[24]。徐振峰等[25]用开顶式生长室对川西北亚高山草地研究,建群种糙野青茅(Deyeuxiascabrescens)和牛尾蒿(A.roxburghiana)的地上生物量均显著增加,伴生种中华羊茅(Festucasinensis)的地上生物量却有所减少,地上总生物量增加。
温度升高造成生态适应性差的野古草(Arundinellahirta)生产力显著下降,致使整个测试群落生产力降低,温度升高对铁杆蒿(A.sacrorum)和黄背草(Themedatriandra)的影响较小[26]。在海北对矮嵩草草甸的模拟增温研究表明,短期的增温条件下群落的总生物量变化不明显,增温导致禾草生物量显著增加,而杂类草生物量显著减少[27]。分析可能原因是生长期延长,生物量增加,植物发育生长速率加快,杂类草成熟期提早,生长期缩短,加之玻璃纤维的存在使暖室内外温度交换减缓,减少了温度日变化,限制干物质积累,最终导致生物量减少。气候暖干化导致植被明显退化,造成高寒草甸禾本科牧草产量不同程度地减少[28]。由此可见,增温对植物的生物量的影响没有定论,有待进一步的研究。
增温对于群落结构及物种的多样性产生深刻的影响。气候变化影响着草原生态系统植物的种类和数量,受人类干扰的草原比天然草原更容易受气候变化的影响,对气候的阻抗小[29]。增温后,不同草原类型的群落结构也得到不同的结果。高寒草甸禾草和莎草(Cyperusrotundus)的盖度减少,杂类草的盖度增加;而沼泽草甸中禾草和莎草的盖度增加,杂类草盖度减少,且大幅度的增温促进了这种趋势[11]。对高寒草甸不同温度梯度上进行模拟增温的研究显示,随着温度的升高,形成明显的群落层片结构,上层以禾草为主,下层以莎草科和杂类草为主。禾草占据上层空间形成郁闭环境,因此矮嵩草等下层植物为了争取更多的阳光和生存空间,植株高度整体增加[30]。对中温型草原群落结构特征研究,预测草原群落高度将有所增加,分层结构也将向上层推移。在内蒙古羊草草原上,优势种羊草、卵穗苔草(Carexduriuscula)、艾蒿(A.vulgaris)在温度升高时初级生产力下降,而次优势种大针茅(S.grandis)、西伯利亚羽茅(Achnatherumsibiricum)、冰草(Agropyroncristatum)的重要值和初级生产力随温度升高而略有升高[31]。若这种趋势继续下去, 大针茅有可能代替羊草, 成为群落的最优建群种。在新疆天山巴音布鲁克草原上,随着温度的升高,物种丰富度增加[32]。相关研究也表明, 随着退化程度的加剧, 草地群落的物种组成、结构和群落多样性发生了重大改变[33],因此, 在未来全球增温的趋势下, 在海拔相对较低地区的矮嵩草草甸的优势种将会发生改变, 优势种矮嵩草有可能被其他植物所代替, 使得群落的结构和功能发生改变。在凋落物的研究中发现,凋落物分解速率加快,降低了凋落物中的氮含量,原因是凋落物中氮的释放加快从而加快了高寒草甸生态系统的氮循环,但是增温却增加凋落物中磷的固定,由此预测未来增温条件下有可能会导致生态系统中磷缺乏[34]。

表2 植物群落水平对模拟增温的响应Table 2 The response of plant community levels of simulated warming
增温后, 羊草草原群落禾本科种子雨及种子库含量减少,实生苗出生数显著增加,克隆苗出生数变化不大;杂类草种子雨及种子库含量增加,实生苗出生数减少,克隆出生数变化不大,羊草群落最终密度减少。植被最终的密度减少,地上植被、种子雨、种子库、实生苗之间的相似性系数与对照比无规律变化[35]。
总的来说,增温后植物群落的生物量、群落结构、种子雨和种子库等都发生了深刻的变化,最终导致群落演替的发生。
2 草地生态系统动物对增温的响应
动物作为草地生态系统中食物链的最顶端,更加容易受到全球气候变暖的影响。在草地生态系统中常见动物主要是昆虫、鼠类和大型食草动物。大多数昆虫需要越冬完成生活史,暖冬可以增加其越冬成活率,有可能造成第二年的大爆发。根据有效积温法则,气候变暖将增加昆虫发生季的有效积温,导致昆虫世代数增加[36]。昆虫飞行活动存在着最适宜温度和临界温度。当温度低于最适宜温度时,气候变暖增强昆虫移动;反之,则不利于昆虫移动。低温还会阻碍昆虫向取食地点的转移,导致昆虫因饥饿而死亡。综合气候数据显示,害虫的暴发与日均温有显著的正相关。许多昆虫的分布受到温度的限制,一些寒冷地区虽然有寄主植物,但昆虫无法完成整个生活史,这些地区在气候变暖条件下会增加昆虫定居的机会。气候变暖会导致昆虫的重新分布和生物入侵,受气候变暖的影响,昆虫倾向于向高纬度(两极方向) 或高海拔分布扩散[37]。气候变暖还会影响到种间关系,主要是指昆虫与天敌之间,不同的发生物候期有可能导致害虫大爆发。气候变化还通过影响植物的开花时间和传粉昆虫活动时间,改变它们之间的相互关系[38]。
鼠类是草地生态系统中最具有代表性的动物,对于鼠类在增温条件下的研究较少。利用开顶式生长小室对海北高寒草甸增温样地的根田鼠进行了研究,结果表明在冬季增温样地内根田鼠的种群密度显著高于对照,而其性别比、存活率、种群平均体重以及年龄结构无明显变化[39]。目前,增温条件下对于草地生态系统中大型食草动物的研究尚未见报道。
3 土壤对增温的响应
土壤作为生命系统支持体系的重要组成部分,对全球增温的响应更加敏感和广泛。土壤有机碳是土壤中较为活跃的部分,土壤有机碳的动态在土壤生产力和全球碳循环中起着十分重要的作用。但温度升高对土壤有机碳动态变化影响的研究仍存在着很大的不确定性。气候变暖对土壤碳库的影响主要表现在两个方面:一方面,温度升高,增加土壤有机质分解,土壤释放CO2和CH4量明显增加,导致土壤中碳的损失[40],并且土壤有机质的分解速率会随温度升高而加快。在气候变暖条件下,温度每升高1 ℃,土壤有机碳损失10%或更高[41];另一方面,空气中CO2浓度升高引起净光合效率提高,从而补偿了气候变暖导致的土壤有机碳的减少,因而土壤有机碳储量的变化可能不大。用红外辐射增温装置对川西北亚高山草原研究发现土壤的有机碳含量下降,而这种降低作用随增温时间的延长而减弱[42]。这是由于在增温初期,微生物活性增强,使得土壤活性有机碳库极易被分解释放出来;而随着增温时间延长,结构性有机碳由于结构稳定、复杂难以分解利用,土壤碳释放表现出对温度敏感性的降低。李娜[43]和潘新丽等[44]对荒漠草原和川西北高寒草甸的研究都证实了增温增加了土壤的有机碳的含量,其原因是增温改变了植物的生长速率,提高了碳固定能力,向土壤输入的碳相应的就增加。这种增温对土壤有机碳不确定性可以认为是不同生态系统的土壤对气候变暖的响应的强度和广度是不同的。
土壤呼吸是一个十分复杂的土壤生态学过程,主要包括土壤微生物呼吸、土壤动物呼吸和植物根系的呼吸。温度是影响总土壤呼吸速率最重要的因子之一。内蒙古荒漠草原土壤呼吸的季节变化和日变化均表现出与土壤温度变化趋势的一致性,土壤呼吸速率与土壤温度呈正相关[45]。相对短期增温后, 高寒草甸土壤呼吸速率显著提高;长期增温下的土壤呼吸与对照没有明显差别,对温度升高表现出适应性[46]。
土壤酶来源于动植物的分泌及其残体和微生物的分泌,是土壤生态系统代谢的一类重要动力。土壤酶参与土壤环境中生物化学过程,与有机物质分解、营养物质循环、能量转移、环境质量等密切相关,并且对环境条件的变化十分敏感,可作为土壤质量的生物指标[47]。温度是控制土壤酶活性的一个重要的因子。大多数研究都集中在脲酶、过氧化物酶、蔗糖酶(分别参与了尿素转化过程,土壤呼吸以及土壤腐殖质化过程)、磷酸酶和土壤蛋白酶等活力的测定。在高寒草甸土壤的研究中发现这3种酶的活力都和增温呈正相关关系,且随着土壤深度的增加,活力下降,并且不同的酶对于温度的敏感性不同。其原因是土壤酶大多数为胞外酶,属于土壤微生物的分泌产物,增温增加了土壤微生物的量,所以土壤酶的活性增加。
增温试验并没有使内蒙古荒漠草原土壤贫瘠化加速,反而能够增加土壤的有机质含量[48]。在地下经过复杂的微生物等综合作用,土壤有机质分解为其他营养元素,供植物生长吸收的同时也增加了地上、地下生物量,从而为土壤提供物质来源并为下一季度的有机质分解提供了物质基础。增温条件下高寒草甸土壤中全氮和有机碳都增加[49]。增温促进碳磷循环,但同时会减缓氮循环[50]。增温对土壤全氮、速效氮、土壤全磷、速效磷含量的影响不显著[51],全钾变化没规律,说明增温对土壤全钾的影响不大,原因是土壤中钾含量主要受土壤矿物质种类的影响,主要来源是矿物风化的部分,而从有机质分解供应的较少。对青藏高寒草甸的研究发现,增温显著增加了木质素含量及“木质素/氮”的比例。说明未来增温条件下,高寒草甸生态系统的碳、氮循环过程将可能加快。
增温处理显著增加了细菌的数量,对真菌和放线菌无显著影响,可能的原因是细菌的生长速度快,对温度变化比较敏感[52]。熊金波等[53]研究发现,短期的增温处理显著地改变了土壤细菌的群落结构而多样性和丰富度没有明显改变,解释原因为增温改变了土壤和植物的特征,间接地影响和作用于土壤微生物群落。
4 展望
在过去的二十多年中,对气候变暖对生态系统的影响进行了大量的研究。在今后的研究中应注意以下几个方面:
1)增温设备不同,不同的生态系统会做出特异性的响应[54]。所以,在今后的试验中可以在同一个研究对象选择不同的增温设备,比较差异,整合结果,完善模型的参数估计和验证。
2)全球变暖的效应是一个长期的效应。生态系统不同组分对温度的敏感性不同,短期的增温试验往往不能得出作为整体的生态系统响应与适应全球变暖的正确结论。所以,在以后的试验中注重原始数据的积累,综合不同生态系统的研究结果。
3)生物圈是全球最大生态系统,是物质流、能量流、信息流高度统一的整体。在以后的试验中不能单一地测定某些指标来衡量全球变暖的效应。另外,以前的研究都是集中在植物个体、群落结构等层次的研究,应增加在基因和蛋白质水平的适应机制研究。
4)全球变暖涉及到许多生态因子的变化,应增加如氮素沉积,降水格局的变化等因子的作用,才能加强对增温条件下生态系统的响应认识的全面性。
5)全球变暖的大背景下,对草地生态系统中植物-害虫-天敌之间相互作用关系的变化为研究重点,加强害虫风险的评估,对于因增温而引起的生物入侵更应该防范。
总之,全球变暖对草地生态系统的影响我们还是知之甚少,只有加强这方面的研究,才能正确地应对全球变暖这一世界性难题,促进生态可持续发展。
[1]IPCC.Climate Change,Impact,Adaptation,and Vulnerability[M].London:Cambridge University Press,2001.
[2]Houghton J T,Ding Y,Griggs D J.Climate Change 2001:The Scientific Basis[M].Cambridge:Cambridge Unversity Press,2001.
[3]IPCC.Climatic Change 2007:The Physical Science Basis[R].IPCC WGI Fourth Assessment Report,Geneva:Intergovernmental Panel on Climate Change,2007.
[4]林慧龙,王军,徐震,陈钟.草地农业生态系统中的碳循环研究动态[J].草业科学,2005,22(4):59-62.
[5]Shen K P,Harte J.Ecosystem climate manipulations[A].Sala O E,Jackson R B,Mooney H A,Howarth R W.Methods in Ecosystem Science[M].New York:Springer-Verlag Press,2000:353-369.
[6]徐光利,金泽新,何维明,王兴龙,车秀霞.不同增温处理对夏蜡梅光合特性和叶绿素荧光参数的影响[J].生态学报,2012,32(20):6343-6353.
[7]Chapin F S,Shaver G R,Giblin A E,Nadelhoffer K J,Laundre J A.Responses of arctic tundra to experimental and observed changes in climate[J].Ecology,1995,76(3):694-711.
[8]Chaves M M,Pereira J S,Maroco J,Rodrigues M L,Ricardo C P P,Osorio M L,Carvalho I,Faria T,Pinheiro C.How plants cope with water stress in the field Photosynthesis and Growth[J].Annals of Botany,2002,89:907-916.
[9]Teramura A H,Sulivan J H,Ziska L H.Interaction of elevated ultraviolet-B radiation and CO2on productivity and photosynthetic characteristics in wheat,rice and soybean[J].Plant Physiology,1990,94(2):470-475.
[10]Kudog,Suzulis.Warming effects on growth, production and vegetation structure of alpine shrubs:A five-year experiment in northern Japan[J].Oecologia,2003,135:280-287.
[11]石福孙,吴宁,吴彦,王乾.模拟增温对川西北两种植物生长以及光和特征的影响[J].应用与环境生物学报,2009,15(6):750-755.
[12]赵建中,刘伟,周华坤,张毓,于龙,许庆民.模拟增温效应对矮嵩草生长特征的影响[J].西北植物报,2006,26(12):2533-2539.
[13]张立荣,牛海山,汪诗平,李英年,赵新全.增温与放牧对矮嵩草草甸4种植物气孔密度和气孔长度的影响[J].生态学报,2010,30(24):6961-6969.
[14]Xu X F,Tian H Q,Wan S Q.Climate warming impacts on carbon cycling in terrestrial ecosystems[J].Journal of Plant Ecology,2007,31(2):175-188.
[15]周华坤,周兴民,赵新全.模拟增温效应对矮嵩草草甸影响的初步研究[J].植物生态学报,2000,24(5):547-553.
[16]Dunne J A,Harte J,Taylor K J.Subalpine meadow flowering phenology responses to climate change intergrating experimental and gradient methods[J].Ecological Monographs,2002,73(1):69-86.
[17]许振柱,周广胜.中国植物学会七十周年年会论文摘要汇编[C].成都:中国植物学会七十周年年会,2003.
[18]Regory S,Steven F O,Ericw P.Effects of lengthened growth season and soil warming on photology and physiology ofPolygonumbistorta[J].Global Change Biology,2000,6(3):357-364.
[19]侯文娟.增温和施氮对松嫩草原羊草叶片矿质元素的影响[D].长春:东北师范大学,2011.
[20]介冬梅,葛勇,郭继勋,刘红梅,石连旋.中国松嫩草原羊草植硅体对全球变暖和氮沉降模拟的响应研究[J].环境科学,2010,31(8):1708-1715.
[21]石福孙,吴宁,罗鹏.川西北亚高山草甸植物群落结构及生物量对温度升高的响应[J].生态学报2008,28(11):5287-5293.
[22]Morgan J A,Lecain D R,Mosier A R,Milchunas D G.Elevated CO2enhances water relations and productivity and affects gas exchange in C3and C4grasses of the Colorado shortgrass steppe[J].Global Change Biology,2001,7(4):251-466.
[23]Rebeeca S,Loik M E,Harte J.Gas change and water relations of two Rocky Mountain shrub species exposed to a climate change manipulation[J].Plant Ecology,2000,146(2):197-206.
[24]刘伟,王长庭,赵建中,许庆民,周立.矮嵩草草甸植物群落数量特征对模拟增温的响应[J].西北植物学报,2010,30(5):995-1003.
[25]徐振峰,胡庭兴,李小艳,张远彬,鲜骏仁,王开运.川西亚高山采伐迹地草坡群落对模拟增温的短期响应[J].生态学报,2009,29(6):2089-2095.
[26]杨永辉,王智平,佐仓保夫,唐常源,新藤静夫.全球变暖对太行山植被生产力及土壤水分的影响[J].应用生态学报,2002,13(6):667-671.
[27]李英年,赵亮,赵新全,周华坤.5年模拟增温后矮嵩草草甸群落结构及生产量的变化[J].草地学报,2004,12(3):236-239.[28]王谋,李勇,白宪洲,黄润秋.全球变暖对青藏高原腹地草地资源的影响[J].自然资源学报2004,19(3):331-335.
[29]Grime J P.Plant Strategies,Vegetation Processes,and Ecosystem Properties[M].2nd edition.New York:John Wiley and Sons,Ltd,2001.
[30]李娜,王根绪,杨燕,高永恒,柳林安,刘光生.短期增温对青藏高原高寒草甸植物群落结构和生物量的影响[J].生态学报2011,31(4):895-905.
[31]刘钦普,林震山.内蒙古草原羊草群落优势物种对气候变暖的响应[J].地理科学进展,2006,25(1):63-71.
[32]李凯辉,胡玉昆,王鑫,范永刚,吾买尔·吾守.不同海拔梯度高寒草地地上生物量与环境因子关系[J].应用生态学报,2007,18(9):2019-2024.
[33]祁彪,张德罡,丁玲玲,张力,汪玺,杨予海.退化高寒干旱草地植物群落多样性特征[J].甘肃农业大学学报,2005,40(5):626-631.
[34]林巧燕,徐广平,苏爱玲,段吉闯,张振华,罗彩云,汪诗平.增温对高寒草甸群落凋落物质量动态变化的影响[J].广西植物,2011,31(6):770-774.
[35]李胜平.羊草群落种子雨、种子库、幼苗再生对模拟增温和施氮的响应[D].长春:东北师范大学,2010.
[36]Thomson L J,Macfadyen S,Hoffmann A.Predicting the effects of climate change on natural enemies of agricultural pests[J].Biological Control,2010,52(3):296-306.
[37]Bale J S,Masters G J,Hodkinson I D,Awmack C,Bezemer T M,Brown V K,Butterfield J,Buse A,Coulson J C,Farrar J,Good J E G,Harrington R,Hartley S,Jones T H,Lindroth R L,Press M C,Symrnioudis I,Watt A D,Whittaker J B.Herb ivory in global climate change research:Direct effects of rising temperature on insect herbivores[J].Global Change Biology,2002,8(1):1-16.
[38]Wall M A,Timmerman-Erskine M,Boyd R S.Conservation impact of climatic variability on pollination of the federally endangered plant,Clematissocialis(Ranunculaceae)[J].Southeast Naturalist,2003,2(1):11-24.
[39]孙平,魏万红,赵亚军,徐世晓,赵同标,赵新全.局部环境增温对根田鼠冬季种群的影响[J].兽类学报,2005,23(5):261-268.
[40]McKane R B,Rastetter E B,Shaver G R,Nadelhoffer K J,GiblinA E,Laundre J A,Chapin F S.Climatic effects on tundra carbon storage inferred from experimental data and a model[J].Ecology,1997,78(4):1170-1187.
[41]Kirschbaum M U F.The temperature dependence of soil organic matter decomposition,and the effect of global warming on soil organic C storage[J].Soil Biology and Biochemistry,1995,27(6):753-760.
[42]陈智,尹华军,卫云燕,刘庆.夜间增温和施氮对川西亚高山针叶林土壤有效氮和微生物特性的短期影响[J].植物生态学报,2010,34(11):1254-1264.
[43]李娜.增温和施氮对荒漠草原生态系统土壤温室气体通量的影响[D].呼和浩特:内蒙古农业大学,2010.
[44]潘新丽,林波,刘庆.模拟增温对川西亚高山人工林土壤有机碳含量和土壤呼吸的影响[J].应用生态学报,2008,19(8):1637-1643.
[45]珊丹.控制性增温和施氮对荒漠草原植物群落和土壤的影响[D].呼和浩特:内蒙古农业大学,2008.
[46]张南翼.模拟增温及氮沉降对松嫩草原土壤氮素、磷素耦合作用的影响[D].长春:东北师范大学,2010.
[47]颜慧,钟文辉,李忠佩,蔡祖聪.长期施肥对红壤水稻土磷脂脂肪酸特性和酶活性的影响[J].应用生态学报,2008,19(1):71-75.
[48]包秀荣.控制性增温和施氮对荒漠草原土壤的影响[D].呼和浩特:内蒙古农业大学,2009,31-35.
[49]李娜,王根绪,高永恒.模拟增温对长江源区高寒草甸土壤养分状况和生物学特性的影响研究[J].土壤学报,2010,47(6):1214-1224.
[50]刘琳,朱霞,孙庚,罗鹏,王蓓.拟增温与施肥对高寒草甸土壤酶活性的影响[J].草业科学2011,28(8):1405-1410.
[51]马琳娜.松嫩草原土壤有机碳、氮素、磷素对模拟增温和施氮的响应[D].长春:东北师范大学,2009:33-34.
[52]谭成玉.松嫩草原微生物活性对模拟增温和施氮的响应[D].长春:东北师范大学,2010,29-33.
[53]熊金波,孙怀博,彭飞,林先贵,薛娴,褚海燕.青藏高原短期增温实验引起土壤细菌群落的快速响应[A].中国土壤学会第十二次全国会员代表大会暨第九届海峡两岸土壤肥料学术交流研讨会论文[C].成都:中国土壤学会,2012:945.
[54]牛书丽,韩兴国,马克平,万师强.全球变暖与陆地生态系统研究中的野外增温装置[J].植物生态学报,2007,31(2):262-271.
(责任编辑 武艳培)
Theresearchofgrasslandecosystemresponsetosimulatedatmosphericwarming
HUANG Wen-hua1, WANG Shu-yan2, HAN Bing1, JIAO Zhi-jun1,HAN Guo-dong1
(1.College of Life Science of Inner Mongolia Agricultural University, Hohhot 010018, China; =2.Agronomy College of Inner Mongolia Agricultural University, Hohhot 010018, China)
Global warming has brought great impacts on the ecological system including extending plants growth period, increasing biomass, increasing photosynthetic rate of the plants with lower optimum temperature without changing stomatal conductance, decreasing the content of sodium, potassium and phosphorus in the leaves.With the temperature increasing, the biomass variations of different types of grassland were inconsistent.Kobresiahumilisin alpine meadow would be substitute by other grass andEymuschinensiswould be substitute byStipagrandis.The insects and rodents were mainly studied with warming which showed that insects increased generations and had a tendency to spread to the high altitude and high latitudes and the rodent increased population density without changing of sex ratio and population age structure.Warming enhanced soil respiration, the activities of soil enzyme and the content of soil bacteria.The present paper summarized the response of each component in grassland ecological system which need further study the influence the grassland ecosystem and biosphere.
global warming; grassland ecological system; plant; animal; soil
HAN Bing E-mail:hb_nmg@163.com
2013-11-13 接受日期:2014-03-11
国家自然科学基金(31060057、31100330、31360113);中科院“西部之光”人才培养项目;内蒙古农业大学草业与草地资源教育部重点实验室
黄文华(1984-),男,甘肃天水人,在读硕士生,主要研究植物分子生物学。E-mail:huangwenhuags@163.com
韩冰(1970-),女(蒙古族),内蒙赤峰人,教授,博士,主要研究植物抗逆生物学。E-mail:hb_nmg@163.com
S812.29
:A
:1001-0629(2014)11-2069-07
10.11829j.issn.1001-0629.2013-0640
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
今日农业(2020年19期)2020-12-14
河南科学(2020年3期)2020-06-02
活力(2019年21期)2019-04-01
河北林业科技(2016年5期)2016-11-08
草业科学(2015年9期)2015-12-11
中国科技信息(2015年2期)2015-11-16
