
濒危植物长叶榧的光合生理生态特性
2014-08-10 12:26金则新郭水良王兴龙
生态学报 2014年22期
王 强,金则新,*,郭水良,管 铭,王兴龙
(1. 台州学院生态研究所, 临海 317000; 2. 上海师范大学生命与环境科学学院,上海 200030)
濒危植物长叶榧的光合生理生态特性
王 强1,金则新1,*,郭水良2,管 铭1,王兴龙1
(1. 台州学院生态研究所, 临海 317000; 2. 上海师范大学生命与环境科学学院,上海 200030)
利用Li- 6400 XT便携式光合作用仪对林窗、林缘、林下3种生境中的长叶榧(Torreyaackii)在春、夏、秋、冬4 个季节里的光合生理生态指标进行测定,探讨3种生境中长叶榧光合能力的季节变化及其对不同光环境的响应,分析其濒危机制,为长叶榧的迁地保护和种群的繁衍复壮提供理论依据。结果显示:(1)林窗、林缘的长叶榧净光合速率(Pn)日变化在夏季呈“双峰”曲线,其它3个季节均呈“单峰”曲线; 林下的Pn日变化在4个季节均呈“单峰”曲线。(2)3种生境中最大净光合速率(Pnmax)、光饱和点(LSP)、表观量子效率(AQY)、最大羧化速率(Vcmax)、最大电子传递速率(Jmax)、磷酸丙糖利用率(TPU)均为夏季最高,使得夏季有较强的光合作用能力,但夏季林窗、林缘的长叶榧出现光合“午休”现象,光补偿点(LCP)、暗呼吸速率(Rd)也较高,使得夏季林窗生境的长叶榧日均Pn明显低于于秋季,林缘的日均Pn与秋季差异不显著。(3)4个季节的日均Pn、Pnmax、LSP、Vcmax、Jma、TPU均为林窗最高,林窗与林下差异显著,表明长叶榧具有阳性植物的特点,适宜生长在光照较强的林窗生境。(4)在生长旺盛的夏、秋季,长叶榧的LSP比伴生物种低,LCP比伴生物种高,对光适应的生态幅度较窄;与伴生物种相比,长叶榧的Pn较低,光合能力较弱,在激烈的种间竞争中处于不利地位,可能成为其濒危的一个重要原因。
濒危植物;长叶榧;伴生植物种;光合生理生态特性
长叶榧(Torreyajackii)又名浙榧、加氏榧,隶属于红豆杉科(Taxaceae)榧树属(Torreya),为我国特有的新生代第三纪孑遗裸子植物,距今约有2亿年的历史,它在研究榧树属的分布、古植物区系以及第四纪冰期的气候等方面有重要意义。长叶榧多生长于山地丘陵地带的深沟峡谷,分布于北向阴坡的沟谷旁或水湿条件好的针阔叶混交林中,常处于群落的中下层[1]。长叶榧生长缓慢[2],分布范围十分狭窄,仅分布于浙江和福建,植株稀少,已列为国家2级保护植物[3]。目前对长叶榧的濒危机理研究较少,仅对其遗传多样性进行了分析,发现长叶榧物种水平遗传多样性较高,种群水平的遗传多样性较低,瓶颈效应、种群隔离和种群体间基因流低等因素使得长叶榧种群间出现了明显的遗传分化[4- 5]。长叶榧如此狭窄的地理分布必然与其自身的生理生态特性有关,而至今有关长叶榧生理生态特性的研究尚未见报道。
植物生理生态学能够把濒危植物生长的环境因子与其生理过程结合起来,有可能从一个侧面揭示植物濒危的原因和机制。 而光是植物生存和生长发育最重要的环境因子之一,不同生态习性的植物具有不同的光环境适应策略。如阳性植物具有较强的光合碳同化能力、高光补偿点和高光饱和点等[6];而阴性植物适应林下弱光环境,具有较低的光补偿点和较高的表观量子效率等[7]。近年来,通过光合生理生态研究来分析濒危植物日趋减少的原因及保护措施越来越受到人们的重视。如夏蜡梅(Sinocalycanthuschinensis)光合能力较弱,在种间竞争中处于不利地位,是导致其濒危的原因之一[8];七子花(Heptacodiummiconioides)[9]、银缕梅(Shaniodendronsubaequale)[10]、金花茶(Camellianitidissima)[11]光合能力处于中下水平,光补偿点较高,光饱和点较低,对光适应的生态幅度较窄,这些都从一个侧面反映了它们在群落中的竞争能力较弱, 在生存竞争中处于不利地位;四合木(Tetraenamongolica)的光合能力、水分利用效率弱于霸王(Zygophyllumxanthoxylon),说明四合木在生理生态学方面存在生存力、适应力差的特性,这些可能是四合木逐渐被霸王取代及濒危的重要内在原因之一[12];五桠果叶木姜子(Litseadilleniifolia)对强光的适应性较差,受森林持续片断化影响,强光胁迫加剧,导致种群衰退[13]。这些研究结果为揭示濒危植物的濒危机理和人工迁地保护等提供了科学依据。
为了有效保护长叶榧资源,必须了解其生理生态习性。通过对在林窗、林缘、林下3种生境中的长叶榧叶片在不同季节里的光合作用参数、叶绿素含量的季节性变化进行研究,分析不同生境、不同季节长叶榧光合生理指标的差异,阐明长叶榧的生理生态习性与适生生境。并与其伴生物种的光合生理指标相比较,探讨其可能与环境因子不协调的生理生态特性,进而揭示其濒危的生理生态学机制。本研究为长叶榧的迁地保护和种群的繁衍复壮提供理论依据。
1 样地与方法
1.1 样地概况
据野外调查,浙江省不仅是长叶榧的模式标本产地,也是长叶榧在全世界的分布中心,其在浙江省主要有两个分布区,一是千里岗地区,二是括苍山地区,其中桐庐、建德、富阳、浦江4县(市)交界的千里岗地区面积最大、株数最多,总面积约2196.3hm2,总株数约54.2万株。样地设置在浙江省桐庐县芦茨乡白云源森林公园,是千里岗地区的一部分[1- 2]。地理位置29°45′N,119°50′E,海拔高度410 m,属亚热带季风气候区,四季分明、气候温和、日照充足、雨量充沛。公园内长叶榧个体数较多,密度较大,具有一定的代表性,研究样地沿沟谷两侧呈带状分布,群落为针阔混交林,长叶榧为主要优势种,其它伴生物种有青冈栎(Cyclobalanopsisglauca)、薄叶润楠(Machilusleptophylla)、盐肤木(Rhuschinensis)、白栎(Quercusfabri)等。林窗(Gap)面积约400m2,林窗中央,近似全光环境,光照强度、气温高,湿度低。林下(Understory)生境郁闭度为70%,光照强度和气温较低,空气湿度较高。林缘(Edge)生境位于群落边缘,部分遮阴,光照强度、气温、空气湿度介于林窗和林下之间。
1.2 测定方法
1.2.1 光合参数测定
于2011年10月(秋季)、2012年1月(冬季)、4月(春季)、7月(夏季)上旬,选择晴朗天气,从每个生境中选取3棵株高1.5 m左右,长势基本一致的长叶榧小树,并在林窗生境中选择与长叶榧株高相近、环境条件基本一致的伴生物种青冈栎、薄叶润楠、盐肤木、白栎各3棵,采用Li- 6400 XT便携式光合作用仪(LI-COR,USA)测定植株顶部健康、成熟叶片的光合参数,测定选用自然光源,从6:00—18:00(冬季仅测8:00—16:00)每2 h测定1次,每次连续测定3 d,每个叶片重复记录3组数据,取平均值。测定时保持叶片自然着生角度和方向不变,采用开放气路,以空气中CO2浓度为气源,测定参数包括:胞间CO2浓度(Ci, μmol/mol)、气孔导度(Gs, mmol m-2s-1)等。
1.2.2 光响应测定
采用Li- 6400 XT便携式光合作用仪,用仪器自带的红蓝光源,在9:00—11:00设置光强梯度由高到低为2000、1500、1200、1000、800、600、400、200、150、100、50、20和0 μmol m-2s-1,测定长叶榧及其伴生物种光合速率。测定前用2000μmol m-2s-1的光强对所测叶片进行20 min诱导。光响应测定条件设置叶室温度为25℃,流速控制在500 μmol m-2s-1,相对湿度为60%。
1.2.3 CO2响应测定
选取与测定光响应相同部位的叶片,以接近叶片饱和光强为测定光强,控制条件与光响应相同,用液态CO2小钢瓶为CO2来源,测定时仪器自动匹配浓度,CO2浓度梯度从1500、1200、1000、800、600、400、200、150、120、100、80、50μmol/mol,每个CO2梯度最大等待时间为300s。
1.2.4 叶绿素含量的测定
选取与测定光合特性相同部位的叶片,装入内有冰袋的保鲜袋,迅速带回实验室,将叶片剪成2 mm正方小块后,称取1.00 g,放入研钵中,加少量CaCO3和25 mL蒸馏水,匀浆,取2.5 mL匀浆液,放入具塞比色管,加10 mL丙酮,提取30 min,过滤,取滤液约3 mL,采用紫外可见分光光度计(T6新世纪,北京)测OD663、OD645。计算如下:
Ca=12.7 OD663-2.69 OD645
Cb=22.9 OD645-4.68 OD663
1.3 数据处理
采用Excel和Origin7.5数据处理软件对所测数据进行整理分析和绘图,用Photosyn Assistant 1.1.2软件得到最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY或φ)和暗呼吸速率(Rd)。曲线拟合方程为[14]:
式中,Pn为净光合速率:Pnmax为最大净光合速率:PAR为光合有效幅射:φ为表观量子效率:Rd为暗呼吸速率:K为光合曲线的曲率,大小介于(0,1)之间。用Photosyn Assistant 1.1.2软件拟合CO2响应曲线,并计算得到最大羧化速率Vcmax、最大电子传递速率Jmax、磷酸丙糖利用率TPU,参照的是Long和Bernacchi[15]的方法进行计算。
所有数据采用SPSS软件进行分析,采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异(α=0.05),试验数据用平均值±标准误差表示。
2 结果与分析
2.1 不同季节环境因子变化
4个季节,林窗、林缘、林下3种生境中光合有效辐射(PAR)的变化规律基本一致,早上光合有效辐射较低,之后逐渐升高,在夏季林窗PAR最大值为1860μmol m-2s-1,4个季节中PAR都以林窗中最高,其次是林缘,林下最低。4个季节中大气温度的季节变化与PAR变化规律一致,在12:00时达到了最大值,夏季最高温度达38℃,4个季节中均以林窗最高,林缘次之,林下最低。大气相对湿度的变化与PAR变化相反,12:00时降到最低点,4个季节中林下大气相对湿度均高于林窗和林缘,夏季林下大气湿度最高,达到了73%。
2.2 不同生境长叶榧光合作用日进程
3 种生境中的长叶榧在春季、秋季、冬季3个季节里净光合速率(Pn)日变化均呈“单峰”曲线(图1),上午随着光强的增加, 叶片的Pn升高,12:00达到最大值,下午随着光照强度减弱Pn下降。夏季,林窗和林缘长叶榧Pn日变化均呈“双峰”曲线,第1峰出现在10:00,在12:00降幅较大,有明显的光合“午休”现象,第2峰则出现在16:00,比第1峰值小;林下的长叶榧Pn日变化比较平缓,呈“单峰”曲线。4个季节中日均Pn均为林窗>林缘>林下,差异显著(表1)。3种生境中的日均Pn为夏季、秋季>春季>冬季;林窗生境中秋季显著高于夏季;林缘生境中秋季与夏季差异不显著;林下生境中夏季显著高于秋季。

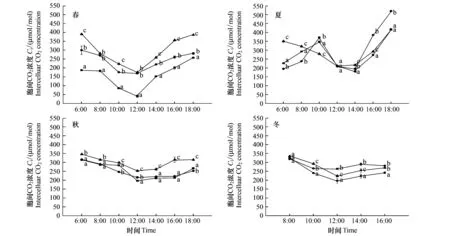
图1 不同生境长叶榧净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)日进程季节性变化Fig.1 Seasonal change in daily precess of net photosynthetic rate、stomatal conductance、intercellular CO2 concentration of Torreya jacki under different habitats数据为平均数±标准误差,不同字母表示差异显著(P<0.05)
表1 不同生境长叶榧日均净光合速率、日均胞间CO2浓度、日均气孔导度季节性变化
Table 1 Seasonal change in daily mean values of net photosynthetic rate、intercellular CO2concentration、stomatal conductance ofTorreyajackiileaves under different habitats

参数Parameters生境Habitats季节Seasons春Spring夏Summer秋Autumn冬Winter日均净光合速率(Pn)林窗2.72±0.231Aa4.03±0.272Ba4.76±0.103Ca1.77±0.147DaDailymeanvaluesofnetphotosynthetic林缘2.08±0.170Ab3.81±0.181Bb3.79±0.153Bb1.58±0.084Cbrate/(μmolm-2s-1)林下1.45±0.268Ac3.31±0.277Bc2.28±0.215Cc1.05±0.118Dc日均气孔导度(Gs)林窗0.06±0.001Aa0.18±0.001Ba0.08±0.001Ca0.03±0.001DaDailymeanvaluesofstomatalconductance/林缘0.04±0.003Ab0.12±0.004Bb0.07±0.002Ca0.03±0.001Da(molm-2s-1)林下0.04±0.001Ab0.08±0.002Bc0.04±0.001Ab0.02±0.001Cb日均胞间CO2浓度(Ci)林窗132.76±4.077Aa260.82±1.842Ba238.69±1.802Ca245.91±6.852DaDailymeanvaluesofintercellularCO2林缘219.74±2.746Ab279.90±2.735Bb245.01±1.279Cb263.07±3.163Dbconcentration/(μmol/mol)林下259.45±1.323Ac264.48±2.781Bc289.46±4.493Cc276.01±4.211Dc
数据为平均数±标准误差,不同小写字母表示同一列差异显著(P<0.05),不同大写字母表示同一行差异显著(P<0.05)
3 种生境中,春季、秋季、冬季长叶榧的气孔导度(Gs)日变化与Pn日变化规律一致,均为“单峰”曲线,峰值在12:00。夏季,林缘、林窗长叶榧的Gs随着光强的增大而增大,到10:00达到最大值,以后下降,到14:00达到最低值后逐渐上升;而林下的Gs为“单峰”曲线,峰值在12:00(图1)。4个季节中林窗、林缘日均Gs均以夏季>秋季>春季>冬季,差异显著(表1)。林下日均Gs也以夏季最高,与其它3个季节差异显著;春季、秋季显著高于冬季。林窗生境的日均Gs在4个季节均显著高于林下(表1);春季、夏季,林窗生境的日均Gs显著高于林缘;夏季、秋季、冬季,林缘生境的日均Gs显著高于林下。
3 种生境中,春季、秋季、冬季长叶榧胞间CO2浓度(Ci)日变化与Pn日变化规律相反,呈“V”字形。夏季,林窗、林缘长叶榧Ci随光强的增强而升高,到10:00达到最大值,以后随着光强的继续增强,Ci开始下降,到14:00达到最低值,后逐渐上升;林下Ci日变化呈“V”字形(图1)。4个季节中林窗和林缘日均Ci均为夏季>冬季>秋季>春季,差异显著(表1)。林下日均Ci以秋季>冬季>夏季>春季,差异显著。春季、秋季、冬季的日均Ci均为林下>林缘>林窗,差异显著;夏季为林缘>林下>林窗,差异显著。
2.3 不同生境长叶榧光合参数变化
由表2可知,3种生境中的长叶榧最大净光合速率(Pnmax)、光饱和点(LSP)、表观量子效率(AQY)均为夏季>秋季>春季>冬季,Pnmax、AQY 4个季节均差异显著;LSP除林窗生境中春季与冬季、林下生境中夏季与秋季无显著差异外,其它均差异显著。4个季节的Pnmax、LSP为林窗>林缘>林下,Pnmax除春季及夏季的林窗与林缘、夏季的林缘与林下无显著差异外,其它均差异显著;LSP 除秋季的林窗与林缘差异不显著外,其余均差异显著。夏季、秋季的AQY为林下>林缘>林窗,差异显著;春季、冬季的AQY林下显著高于林缘、林窗。林窗中的光补偿点(LCP)为夏季显著高于其它3个季节,秋季显著高于冬季、春季;林缘的LCP为夏季>秋季>冬季>春季,差异显著;林下的LCP为夏季、秋季显著高于冬季、春季,冬季显著高于春季。春季、夏季的LCP均为林窗>林缘>林下,差异显著;秋季、冬季为林窗、林缘显著高于林下。林窗中的暗呼吸速率(Rd)为冬季显著低于其它季节;林缘为夏季>春季>秋季>冬季,差异显著;林下为夏季显著高于其它季节。夏季、秋季的Rd均为林窗>林缘>林下,差异显著;春季、冬季的Rd为林窗、林缘显著高于林下。

表2 不同生境长叶榧光合参数季节性变化
数据为平均数±标准误差,不同小写字母表示同一列差异显著(P<0.05),不同大写字母表示同一行差异显著(P<0.05)
3种生境中的最大羧化速率(Vcmax)、最大电子传递效率(Jmax)、磷酸丙糖利用率(TPU)均为夏季>秋季>春季>冬季,Vcmax除林缘生境的春季与冬季无显著差异外,其余均差异显著;Jmax4个季节均差异显著;TPU除林窗生境的春季与秋季差异不显著外,其余均差异显著。春季、秋季、冬季的Vcmax为林窗>林缘>林下,夏季为林缘>林窗>林下,差异显著。春季、秋季、冬季的Jmax为林窗>林缘>林下,夏季为林缘>林窗>林下,差异显著。4个季节的TPU均为林窗>林缘>林下,除春季的林缘与林下、冬季的林窗与林缘差异不显著外,其余均差异显著。
2.4 不同生境长叶榧叶片叶绿素含量变化
叶绿素在光合作用中起着吸收光能的作用,其含量的高低直接影响到植物光合作用的强弱[16]。由表3可知,3 种生境中长叶榧叶绿素a、叶绿素b、叶绿素a+b含量均以冬季最低,除林窗生境的叶绿素b外,冬季与其它季节均差异显著。春、夏、秋3个季节的叶绿素a、叶绿素b、叶绿素a+b含量均为林下大于林窗,除夏季、秋季的叶绿素b含量林下与林窗差异不显著外,其它均差异显著。而冬季则是林窗显著高于林下。
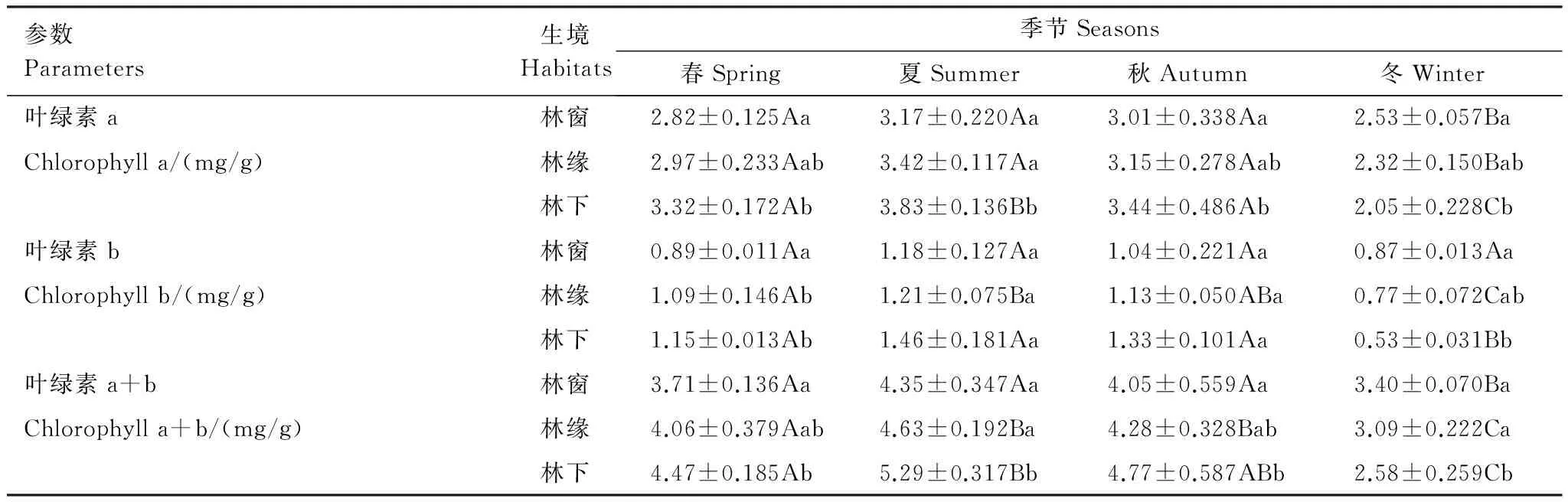
表3 不同生境长叶榧叶绿素含量季节性变化
数据为平均数±标准误差,不同小写字母表示同一列差异显著(P<0.05),不同大写字母表示同一行差异显著(P<0.05)
2.5 长叶榧及其伴生植物光合作用日进程
林窗长叶榧及其伴生植物Pn日变化如图2所示,春季、秋季、冬季3个季节Pn日变化呈“单峰”曲线,峰值均出现在12:00。春季峰值最高的为盐肤木,最低的为薄叶润楠;秋季峰值最高的为青冈栎,最低的为薄叶润楠;冬季峰值最高的为青冈栎,最低的为长叶榧。夏季林窗长叶榧及其伴生植物Pn日变化均呈“双峰”曲线,第1峰出现在10:00,峰值较大的为盐肤木、白栎、青冈栎,分别比长叶榧高48.1%、29.9%、26.7%,差异显著,第2峰则出现在16:00,比第1峰值小,峰值较高的也为盐肤木、白栎、青冈栎,分别比长叶榧高46.4%、48.1%、50.5%,差异显著。
4个季节的长叶榧及伴生物种日均Pn见表4,春季除薄叶润楠外,其它伴生物种日均Pn均显著高于长叶榧;夏季各伴生物种日均Pn均显著高于长叶榧;秋季青冈栎、白栎的日均Pn显著高于长叶榧;冬季常绿树种青冈栎、薄叶润楠的日均Pn均显著高于长叶榧。春季、夏季、秋季3个季节月平均Pn大小为青冈栎>白栎>盐肤木>长叶榧>薄叶润楠,差异显著,长叶榧在生长季节内与其伴生物种相比Pn处于中下水平,表明长叶榧的光合能力较弱,在激烈的种间竞争中将处于不利地位。
2.6 长叶榧及其伴生植物光饱和点(LSP)和光补偿点(LCP)
从表5可以看出,长叶榧及其伴生物种LSP季节变化规律相同,春季较低,在夏季达到最大值,以后LSP不断降低,在冬季降到最低值。在春季、夏季长叶榧LSP显著低于其伴生物种;秋季长叶榧LSP除显著低于青冈栎外,显著高于另外3种伴生物种。冬季长叶榧LSP也显著低于其常绿伴生植物。春季长叶榧LCP显著低于其4种伴生物种,夏季长叶榧LCP显著高于其4种伴生物种,秋季也显著高于其常绿伴生植物。

图2 长叶榧及其伴生种Pn日进程季节性变化Fig.2 Seasonal change in daily precess of net photosynthetic rate of Torreya jacki and its accompanying species under different habitats数据为平均数±标准误差,不同字母表示差异显著(P<0.05)
Table 4 Seasonal change in daily mean values of net photosynthetic rate ofTorreyajackiileaves and its accompanying species under different habitats

参数Parameters生境Habitats季节Seasons春Spring夏Summer秋Autumn冬Winter日均净光合速率(Pn)长叶榧Torreyajackii2.72±0.233Aa4.03±0.271Ba4.76±0.107Ca1.77±0.146DaDailymeanvaluesofnet青冈栎Cyclobalanopsisglau-ca2.97±0.187Ab8.75±0.231Bb7.30±0.294Cb2.26±0.057Dbphotosyntheticrate/薄叶润楠Machiluslepto-phylla2.33±0.132Ac4.86±0.115Bc3.88±0.259Cc2.07±0.510Db(μmolm-2s-1)白栎Quercusfabri2.91±0.042Ab7.28±0.307Bd6.60±0.255Cd盐肤木Rhuschinensis3.25±0.343Ad7.82±0.046Be1.91±0.041Ce
数据为平均数±标准误差,不同小写字母表示同一列差异显著(P<0.05),不同大写字母表示同一行差异显著(P<0.05)
3 讨论
光合作用属于酶促反应,光合能力的差异与其所处的环境有关。长叶榧在夏季的林窗和林缘生境中,长叶榧净光合速率(Pn)日变化呈“双峰”曲线,出现明显的光合“午休”现象。这是因为夏季林窗、林缘生境的气温和光照强度都很高,特别是中午12:00左右,林窗生境最大PAR达1860 μmol m-2s-1、最高温度达38℃,林缘生境PAR达1680 μmol m-2s-1、温度达35℃,而此时叶片蒸腾速率大,为了控制水分的散失,Gs逐渐降低,与外界气体交换受阻,导致Ci大幅度降低,进入叶肉细胞内用于光合作用的原料CO2供应不足,使Pn降低。据Farquhar等的观点,如果Pn和Ci变化方向相同,并且Gs降低时Pn的降低是气孔因素造成的;如果Pn和Ci变化方向相反,并且Gs增大,则Pn的下降主要是由非气孔限制,即叶肉细胞同化能力的降低引起的[17]。可见夏季林窗、林缘生境中的长叶榧光合“午休”现象是气孔因素造成的。
表5 长叶榧及其伴生种光饱和点和光补偿点季节变化
Table 5 Seasonal change in Light saturation point and Light compensation point ofTorreyajackiileaves and its accompanying species under different habitats

参数Parameters季节Seasons物种Species长叶榧Torreyajackii薄叶润楠Machilusleptophylla青冈栎Cyclobalanopsisglauca白栎Quercusfabri盐肤木Rhuschinensis光饱和点(LSP)春462±16.819Aa628±25.637Ba752±24.494Ca564±23.025Da530±22.861DaLightsaturationpoint/夏927±34.456Ab940±38.375Bb1288±49.499Cb1206±48.826Cb1108±45.233Db(μmolm-2s-1)秋752±30.700Ac604±20.575Ba808±28.903Cc700±28.577Dc160±6.531Ec冬428±17.473Aa467±6.153Bc544±9.961Cd光补偿点(LCP)春10±0.408Aa18±1.306Ba13±1.469Ca18±1.143Ba18±2.776BaLightcompensation夏20±0.816Ab10±0.463Bb4±0.372Cb4±0.165Cb4±0.489Cbpoint/(μmolm-2s-1)秋14±0.571Ac8±0.489Bb8±0.177Bc25±0.255Cc30±0.326Dc冬10±0.408Aa8±0.326Ab8±0.189Ac
数据为平均数±标准误差,不同小写字母表示同一列差异显著(P<0.05),不同大写字母表示同一行差异显著(P<0.05)
对光能利用能力的高低是植物光合速率存在差异的重要原因,LSP和LCP反应了植物对光照条件的要求,分别体现了对强光和弱光的利用能力。夏季,3种生境中的长叶榧LSP和LCP均较高,反映出长叶榧在夏季对强光的利用能力增强,而对弱光的利用能力减弱。AQY是植物光合作用对光的利用效率[18],3种生境中长叶榧AQY均以夏季最高,表明其在夏季对光能的利用效率高。而夏季3 种生境中的长叶榧Rd均有所提高,呼吸消耗有所增大。此外,Vcmax的大小主要受到Rubisco蛋白酶的活性、数量的影响,Rubisco蛋白酶是植物叶片光合作用过程中催化RuBP固定CO2的关键性酶[19];Jmax代表RuBP再生能力,低的电子传递速率会限制光合磷酸化作用和NADPH的再生,从而使叶片的Vcmax受到一定限制[20];磷酸丙糖(TP)是光合作用的最初产物,TPU利用率较低会引起TP积累,会对光合作用产生负的反馈调节引起光合速率的降低。长叶榧叶片Vcmax、Jmax、TPU都以夏季最高,说明夏季长叶榧在光合作用过程中催化RuBP固定CO2的Rubisco的活性和数量的增加,较高的电子传递速率促进了光合磷酸化作用和NADPH的合成,提高了RuBP再生能力,这些特性使它们充分利用光能进行碳同化,进而促使光合作用能力的提高。由此可看出,在夏季强光、高温的条件下,虽然3种生境中的长叶榧对弱光的利用率有所降低、暗呼吸作用有所增大,但LSP、AQY、Vcmax、Jmax、TPU明显增高,因而在夏季表现出较强的光合作用能力,Pnmax也是夏季最大。但夏季林窗、林缘的长叶榧出现光合“午休”现象,使得夏季林窗长叶榧的日均Pn明显低于秋季、林缘的日均Pn与秋季差异不显著。
光是影响植物光合作用的重要因子,不同生境下光强的变化对长叶榧光合特性有较大的影响。林窗生境近似全光环境,PAR最大;林缘生境光照振幅较大;林下生境光照变化相对平缓,PAR约为全光的17%。在4个季节中长叶榧叶片的LSP、Vcmax、Jmax、TPU均为林窗最高,林窗与林下差异显著。说明林窗生境的长叶榧通过提高对强光的利用能力,提高最大羧化速率、最大电子传递效率、磷酸丙糖利用率,来提高光合作用能力。表现在林窗的日均Pn均显著高于林缘、林缘显著高于林下,这与银杉(Cathayaargyrophylla)随着PAR的降低Pn显著降低[21]的结果一致。4个季节的Pnmax也是林窗最高,林窗与林下差异显著,可看出长叶榧适宜生长在光照强的林窗生境。而林下光强较弱,低的LCP能更好的利用弱光进行光合作用;高的AQY对光能的利用效率提高;低的Rd也有助于减少呼吸消耗,在光合速率较低时积累较多有机物,增强了对弱光环境的适应,表明长叶榧的光合特性对光强的变化有一定的可塑性[22]。枫香(Liquidambarformosana)和四川大头茶(Gordoniaacuminate)也反映出类似的结果,随着PAR的降低两种幼苗的LSP、LCP、Rd都有明显的下降趋势[23]。通常耐荫树种随着PAR的降低,Pnmax仅有略微的降低[24],光合能力一般较弱,在生理上的可塑性较差[25]。而长叶榧的光合作用能力较强,随着PAR的降低,Pnmax下降明显,光合生理具有一定的可塑性,可看出长叶榧是阳性树种。
叶绿素可以将捕获的光能转化为化学能,是光合作用的光敏催化剂,其含量和比例是植物适应和利用环境因子的重要指标[26]。林下长叶榧叶片叶绿素总量要高于林窗,这可能是因为林下PAR弱,植物为适应弱光生境,通过增加叶绿素的量来弥补光合作用中“质”的不足,更好的捕获光能。已有研究表明虽然弱光环境中叶绿素含量上升,但由于光照不足,其光合速率却呈下降趋势[27]。3种生境中,不同季节叶绿素含量以夏季最高,表明夏季强光、高温的环境下,促进了长叶榧叶绿素的生物合成和积累,使其含量上升,增强了夏季长叶榧光能的吸收和转化能力,这也使得夏季长叶榧光合能力高于其它3个季节的原因之一。
长叶榧分布范围狭窄,种群日趋缩小,造成其濒危的原因是多方面的,物种生物学特性是导致物种濒危的主要原因之一。Pn的大小决定着植物体物质积累能力的高低,会影响植物生长的快慢。从长叶榧与其伴生植物光合能力的比较可以看出,4个季节中长叶榧日均Pn均处于中下水平,并且由于夏季强光和高温限制了长叶榧光合能力,Pn午间下降持续时间最长,年总光合量较小,有机物积累能力低,生长速度慢,竞争能力弱,因而水热条件优越的地段,往往被竞争能力强的树种所占据,不得不退到坡度大,裸岩多,土壤较为贫瘠的下坡和山麓,阳光充足的南坡和山脊较为少见[1],这可能是造成长叶榧分布范围日趋缩小,成为濒危物种的原因之一。此外,生长旺盛的夏季、秋季长叶榧LCP显著高于其4种伴生植物,表明长叶榧不耐荫,而其常分布于各类群落的中下层,由于群落郁闭度较高[1],使长叶榧幼苗的生长处于不利地位,常因光照不足而饥饿死亡,天然更新能力弱[28],这也可能导致长叶榧成为濒危物种的另一原因。而LSP显著低于其4种伴生植物,说明长叶榧也不不耐强光,对光适应的生态幅度窄。
综上所述,长叶榧Pn日变化除夏季的林窗、林缘生境为“双峰”曲线,出现光合“午休”现象外,其它季节、其它生境均为“单峰”曲线,长叶榧的光合“午休”现象是气孔因素造成的。长叶榧在夏季表现出较强的光合作用能力,Pnmax也是夏季最大,但夏季林窗、林缘的长叶榧出现光合“午休”现象,使得夏季林窗的日均Pn明显低于秋季、林缘的日均Pn与秋季差异不显著。4个季节中长叶榧的日均Pn、Pnmax均以林窗最高,林窗的光合能力显著高于林下,可看出长叶榧适宜生长在光线较好的林窗生境中。长叶榧的光合作用能力较强,光合生理具有一定的可塑性,表明长叶榧是阳性树种。造成长叶榧濒危的原因很多,人类过度樵采是一方面因素,而另一方面,在自然状态下长叶榧光合能力较弱,植株生长缓慢,对光适应的生态幅度要较窄,这些都反应出长叶榧在群落中的竞争能力较弱,在生存竞争中处于不利地位,因此急需对长叶榧进行有效的保护。
[1] Wang C T. Present situation of wildTorreyajackiiresource and its protecting measure in Zhejiang province. Journal of Anhui Agriculture Science, 2005, 33(3): 432- 432, 450- 450.
[2] Wang C T. The biological characteristics of wildTorreyajackiiand protection research. Practical Forestry Technology, 2005, (10): 6- 7.
[3] Mistry of Environmental Protection, Institute of Botany, The Chinese Academy of Sciences. China Rare and Endangered Plants List: Volume 1. Beijing: Science Press, 1987.
[4] Li J M, Jin Z X. Genetic variation and differentiation inTorreyajackiiChun an endangered plant endemic to China. Plant Science, 2007, 172(5): 1048- 1053.
[5] Li J H, Jin Z X, Li J M. Genetic diversity of endangered plantTorreyajackii: A study with RAPD markers. Chinese Journal of Applied Ecology, 2007, 18(12): 2661- 2667.
[6] Scholes J D, Press M C, Zipperlen S W. Differences in light energy utilization and dissipation between dipteroearp rain forest tree seedlings. Oecologia, 1997, 109(1): 41- 48.
[7] Senevirathna A M W K, Stirling C M, Rodrigo V H L. Growth, photosynthetic performance and shade adaptation of rubber (Heveabrasiliensis) grown in natural shade. Tree Physiology, 2003, 23(10): 705- 712.
[8] Jin Z X, Li J M, Ma J E. Photosynthesis traits of the endangered plant speciesSinocalycanthuschinensis. Journal of Zhejiang University: Science Edition, 2011, 38(6): 682- 688.
[9] Jin Z X, Ke S X. The photosynthetic characteristics of the main species of theHeptacodiummiconioidescommunity in Tiantai Mountain of Zhejiang Province, China. Acta Ecologica Sinica, 2002, 22(10): 1645- 1652.
[10] Zhu T J, Yue C L, Jin S H. Ecophysiological trait comparison ofShaniodendronsubaequaleand accompanying species. Journal of Zhejiang Forestry College, 2008, 25(2): 176- 180.
[11] Shi S L, Wang Y C, Zhou H B, Zhou J H. Comparative analysis of water related parameters and photosynthetic characteristics in the endangered plantTetraenamongolicaMaxim. and the closely relatedZygophyllumxanthoxylon(Bunge) Maxim. Acta Ecologica Sinica, 2012, 32(4): 1163- 1173.
[12] Xiong Z C, Luo W H, Wang M L, Wang D R, Wen X Y. Comparative study on photosynthetic characteristics ofCamellianitidissimaand its accompanying species. Guangxi Science, 2012, 19(2): 201- 204.
[13] Guo X R, Cao K F, Xu Z F. Response of photosynthesis and antioxygenic enzymes in seedlings of three tropical forest tree species to different light environments. Chinese Journal of Applied Ecology, 2004, 15(3): 377- 381.
[14] Zhu H, Ma R J. Photosynthetic characteristics comparison between an invasive plant,LantanacamaraL., and associated species. Acta Ecologica Sinica, 2009, 29(5): 2701- 2709.
[15] Long S P, Bernacchi C J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. Journal of Experimental Botany, 2003, 54(392): 2393- 2401.
[16] He W M, Dong M. Growth and physiological features ofSalixmatsudanaon the Mu Us Sandland in response to shading. Chinese Journal of Applied Ecology, 2003, 14(2): 175- 178.
[17] Farquhar S P, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33(1): 317- 345.
[18] Farquhar G D, Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species. Planta, 1980, 149(1): 78- 90.
[19] Hartman F C, Harpel M R. Structure, function, regulation, and assembly of ribulose- 1, 5-Bisphosphate carboxylase/oxygenase. Annual Review of Biochemistry, 1994, 63(1): 197- 234.
[20] Harley P C, Thomas R B, Reynolds J F, Strain B R. Modelling photosynthesis of cotton grown in elevated CO2. Plant Cell and Environment, 1992, 15(3): 271- 282.
[21] Zhang W F, Fan D Y, Xie Z Q, Jiang X H. The seasonal photosynthetic responses of seedlings of the endangered plantCathayaargyrophyllato different growth light environments. Biodiversity Science, 2005, 13(5): 387- 397.
[22] Strauss-Debenedetti S, Bazzaz F A. Plasticity and acclimation to light in tropical Moraceae of different succession positions. Oecologia, 1991, 87(3): 377- 387.
[23] Wang R. The Response and Acclimation of Two Different Plant Functional Groups to Different Light Regimes in Subtropical Evergreen Broad-leaved Forest [D]. Beijing: Chinese Academy of Forestry, 2007.
[24] Reich P B, Walters M B, Tjoelker M G, Vanderklein D, Buschena C. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate. Functional Ecology, 1998, 12(3): 395- 405.
[25] Lovelock C E, Jebb M, Osmond C B. Photoinhibition and recovery in tropical plant species: response to disturbance. Oecologia, 1994, 97(3): 297- 307.
[26] Liu Y Q, Sun X Y, Wang Y, Liu Y. Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters ofUrticadioica. Acta Ecologica Sinica, 2007, 27(8): 3457- 3464.
[27] Chi W, Wang R F, Zhang C L. Changes of photosynthetic characteristics of strawberry leaf under shading. Chinese Journal of Applied Ecology, 2011, 12(4): 566- 568.
[28] Gao Z W. Study on the biological character and conservation of endemic speciesTorreyajackii. Chinese Biodiversity, 1997, 5(3): 206- 209.
参考文献:
[1] 王昌腾. 浙江省野生长叶榧资源现状及保护对策. 安徽农业科学, 2005, 33(3): 432- 432, 450- 450.
[2] 王昌腾. 野生长叶榧生物学特性与保护研究. 林业实用技术, 2005, (10): 6- 7.
[3] 国家环境保护局, 中国科学院植物研究所. 中国珍稀濒危保护植物名录: 第1册. 北京: 科学出版社, 1987.
[5] 李建辉, 金则新, 李钧敏. 濒危植物长叶榧群体遗传多样性的RAPD分析. 应用生态学报, 2007, 18(12): 2661- 2667.
[8] 金则新, 李钧敏, 马金娥. 濒危植物夏蜡梅光合生理生态特性. 浙江大学学报: 理学版, 2011, 38(6): 682- 688.
[9] 金则新, 柯世省. 浙江天台山七子花群落主要植物种类的光合特性. 生态学报, 2002, 22(10): 1645- 1653.
[10] 朱汤军, 岳春雷, 金水虎. 银缕梅和伴生植物光合生理生态特性比较. 浙江林学院学报, 2008, 25(2): 176- 180.
[11] 石松利, 王迎春, 周红兵, 周健华. 濒危种四合木与其近缘种霸王水分关系参数和光合特性的比较. 生态学报, 2012, 32(4): 1163- 1173.
[12] 熊忠臣, 骆文华, 王满莲, 王代荣, 文香英. 金花茶与其伴生植物光合特性的比较研究. 广西科学, 2012, 19(2): 201- 204.
[13] 郭晓荣, 曹坤芳, 许再富. 热带雨林不同生态习性树种幼苗光合作用和抗氧化酶对生长光环境的反应. 应用生态学报, 2004, 15(3): 377- 381.
[14] 朱慧, 马瑞君. 入侵植物马缨丹(Lantanacamara)及其伴生种的光合特性. 生态学报, 2009, 29(5): 2701- 2709.
[16] 何维明, 董鸣. 毛乌素沙地旱柳生长和生理特征对遮荫的反应. 应用生态学报, 2003, 14(2): 175- 178.
[21] 张旺锋, 樊大勇, 谢宗强, 蒋晓晖. 濒危植物银杉幼树对生长光强的季节性光合响应. 生物多样性, 2005, 13(5): 387- 397.
[23] 王荣. 亚热带常绿阔叶林两个不同功能组树种幼苗对不同光环境的响应与适应 [D]. 北京: 中国林业科学研究院, 2007.
[26] 刘悦秋, 孙向阳, 王勇, 刘音. 遮荫对异株荨麻光合特性和荧光参数的影响. 生态学报, 2007, 27(8): 3457- 3464.
[27] 迟伟, 王荣富, 张成林. 遮荫条件下草莓的光合特性变化. 应用生态学报, 2011, 12(4): 566- 568.
[28] 高兆蔚. 我国特有树种长叶榧树的生物学特性与保护问题研究. 生物多样性, 1997, 5(3): 206- 209.
Photosynthetic traits of the endangered plant speciesTorreyajackii
WANG Qiang1, JIN Zexin1,*, GUO Shuiliang2, GUAN Ming1, WANG Xinglong1
1InstituteofEcology,TaizhouUniversity,Linhai317000,China2SchoolofLifeandEnvironmentScience,ShanghaiNormalUniversity,Shanghai200030,China
A portable Li- 6400 XT photosynthesis measuring system (LI-COR Biosciences, Lincoln, NE, USA) was used to explore howTorreyajackii, an endangered species endemic to China, adapts to the environment and to illustrate and determine various photosynthetic characteristics ofT.jackiiin three natural habitats with different light conditions, specifically gap, edge and understory habitats. Also, the physiological mechanisms that have resulted in the endangered status of this species were explored to provide baseline reference data in support of off-site conservation and population rejuvenation ofT.jackii. The results forT.jackiigrowing in forest gap and edge habitats show that when the diurnal variation in the photosynthetic rates in the summer is graphed, typical curves with double peaks appear; in addition, photosynthetic rates in the other seasons show curves with a single peak as do photosynthetic rates in understory habitat based on data in all four seasons. The net photosynthetic rate (Pnmax), light saturation point (LSP), apparent quantum yield (AQY), maximum rate of carboxylation (Vcmax), maximum rate of electron transport (Jmax), and triose phosphate use rate (TPU) were highest in summer in all habitats; this indicates the photosynthetic capacity ofT.jackiiis significantly higher in summer than in other seasons. The graphs indicate a midday depression of photosynthesis occurs in summer in gap and edge habitats when the dark respiration rate (Rd) and light compensation point (LCP) are also higher. As a result, the daily mean values ofPnin gap habitat are significantly lower than in autumn while those ofPnin edge habitat are similar to the daily mean values observed in autumn. In all four seasons the daily mean values ofPn,Pnmax, LSP,Vcmax,Jma, and TPU in gap habitat are the highest with significant differences between those in gap and understory habitat. These findings indicate that the photosynthetic characteristics ofT.jackiiare typical for a plant growing in sunny habitats, meaningT.jackiigrows more vigorously in gap habitat with adequate light. In summer and autumn, the LSP ofT.jackiiis lower than those of other plants typically associated with its habitats, while the LCP is higher than those of the associated plants, which indicates thatT.jackiihas a narrow ecological range of adaptability to light conditions. ThePnofT.jackiiis lower than those of the associated plants in the community, indicating thatT.jackiiis at a disadvantage during interspecific competition with its poor ability to adapt photosynthetically to a wide variety of light conditions; this might be an important factor contributing to the endangered status ofT.jackii.
endangered plant;Torreyajackii; associations plant species; physiological characteristics of photosynthesis
浙江省自然科学基金项目(Y505331)
2013- 02- 26; 网络出版日期:2014- 03- 14
10.5846/stxb201302260307
*通讯作者Corresponding author.E-mail: jzx@tzc.edu.cn
王强,金则新,郭水良,管铭,王兴龙.濒危植物长叶榧的光合生理生态特性.生态学报,2014,34(22):6460- 6470.
Wang Q, Jin Z X, Guo S L, Guan M, Wang X L.Photosynthetic traits of the endangered plant speciesTorreyajackii.Acta Ecologica Sinica,2014,34(22):6460- 6470.
猜你喜欢
科学(2022年4期)2022-10-25
资源导刊(2021年9期)2021-10-02
落叶果树(2021年6期)2021-02-12
现代农业科技(2019年21期)2019-12-25
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
体育科技(2016年2期)2016-02-28
少儿科学周刊·儿童版(2015年10期)2015-11-07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
