
莳萝蒿适应盐渍环境的Na+区域化方式和生理特征
2014-08-08 02:15邱念伟杨翠翠卢正珂李遵宁岳贤军程秀秀许莹莹
生态学报 2014年21期
邱念伟,杨翠翠, 卢正珂, 李遵宁, 岳贤军, 程秀秀, 许莹莹, 周 峰
(1. 曲阜师范大学生命科学学院, 曲阜 273165; 2. 南京晓庄学院生物化工与环境工程学院, 南京 211171)
莳萝蒿适应盐渍环境的Na+区域化方式和生理特征
邱念伟1,杨翠翠1, 卢正珂1, 李遵宁1, 岳贤军1, 程秀秀1, 许莹莹1, 周 峰2,*
(1. 曲阜师范大学生命科学学院, 曲阜 273165; 2. 南京晓庄学院生物化工与环境工程学院, 南京 211171)
莳萝蒿是广泛分布在我国北方的一种特殊类型的菊科盐生植物,阐明莳萝蒿特殊的耐盐机制和生理特征有助于丰富植物抗盐性研究的内容。用0、100、200、300、400 mmol/L NaCl处理莳萝蒿7 d后,比较莳萝蒿盐处理植株与对照植株在生长和生理方面的差异,并详细分析了Na+在莳萝蒿体内的积累水平和区域化方式。结果显示:莳萝蒿虽然能够耐受400 mmol/L NaCl,但盐处理显著抑制了莳萝蒿的生长,整株鲜重随着盐处理浓度的升高逐渐减小。在水分生理方面,随着盐处理浓度的升高,莳萝蒿叶片细胞的渗透调节能力逐渐增强,其叶片肉质化程度却呈逐渐降低的趋势。分析盐处理对光合作用的影响发现,盐处理后莳萝蒿叶片光合速率与气孔导度显著下降,而其PSⅡ光化学活性并未受到抑制,叶绿素含量甚至逐渐增大,说明盐处理后莳萝蒿叶片光合速率的降低主要是由于气孔因素造成的,而不是由于光合结构被破坏。莳萝蒿体内的Na+含量随着盐处理浓度的升高显著增加,400 mmol/L NaCl条件下叶、茎、根中的Na+含量分别高达321.4、242.1和182.3 μmol/g鲜重;莳萝蒿体内的Na+70%以上积累在叶片内,而叶片内98%左右的Na+积累在叶片原生质体中,叶片原生质体中的Na+平均浓度是质外体1.2—1.8倍,推测其叶片细胞内存在着有效的Na+区域化机制。盐处理后莳萝蒿叶片液泡膜V-H+-ATPase的质子泵活性比对照增加了30%—50%,液泡膜Na+/H+逆向转运活性则增加至对照的4—7倍,进一步证实莳萝蒿叶片具有较强的液泡Na+区域化能力。随着盐处理浓度的升高,Na+在叶片中的分布比例相对减少,V-H+-ATPase的质子泵活性和Na+/H+逆向转运活性增幅也减缓。这种Na+区域化能力使莳萝蒿获得了较强的耐盐性,有效保护了其光系统,降低了细胞汁液渗透势。但是盐处理后这种耐盐方式并不能阻止莳萝蒿叶片肉质化程度和光合活性下降,莳萝蒿生长仍然受盐抑制,说明Na+区域化是莳萝蒿适应盐渍环境的必要条件而非充分条件。
莳萝蒿;Na+区域化;生长;光合;适应
莳萝蒿(ArtemisiaanethifoliaMattf)广泛分布在我国北方盐碱地区,是一种特殊的菊科盐生植物,它既不具备真盐生植物的快速生长及稀盐能力,又不具备泌盐植物的泌盐结构,也不具备拒盐植物有效的拒盐方式[1],却能够在高盐条件下生存并完成生活史。因此莳萝蒿的耐盐机制具有特殊性,但其耐盐机制还未见报道。在盐渍条件下莳萝蒿吸收了大量的Na+并能够使细胞质中的代谢活动不受伤害,说明其具有非常有效的Na+区域化方式。本文拟从个体、组织和细胞3个水平上分析莳萝蒿在盐渍条件下的离子区域化方式和生理特征,阐述其耐盐机制。
在前人提出的众多耐盐机制中,最受关注的是Na+/H+逆向转运蛋白,它广泛存在于生物膜上,具有调节细胞pH值和Na+平衡的作用[2- 5]。通过转Na+/H+逆向转运蛋白已经使近百种盐敏感植物获得了耐盐性,充分证实了Na+/H+逆向转运蛋白在植物耐盐性中的作用[6- 9]。但是这些通过转Na+/H+逆向转运蛋白基因获得耐盐性的植物虽然耐盐能力提高,但其仍然不能真正“适应”盐渍环境。“适应”是指植物处于不适宜的环境时,能够通过自身的形态或生理变化,将环境的不利影响降到最低,并最大限度地利用环境的有益因素,使植物达到最佳的生活状态[10]。能够真正“适应”盐渍环境的植物叫做真盐生植物,真盐生植物主要分布在藜科植物中,如碱蓬属、滨藜属、猪毛菜属和盐穗木属等。这类植物的特征是“喜盐”,适当的盐度能够促进其生长,如碱蓬最适盐浓度是200—300 mmol/L NaCl[11]。但是,包括莳萝蒿、芦苇、二色补血草在内的很多盐生植物在盐渍环境下生长均受到抑制,因此它们均不是真盐生植物,通过转基因获得耐盐性的植物也均没有达到“适盐性”目标[6- 9,12]。盐渍环境对非真盐生植物来讲仍然是一种胁迫,而对真盐生植物来讲则是一种适宜环境,Na+是真盐生植物重要的渗透调节物质[11,13]。以莳萝蒿为代表植物,分析其Na+区域化机制和生长特征,有助于了解这类非适盐植物耐盐但不能“适盐”的主要原因,为植物耐盐性研究提供新内容。
1 材料与方法
1.1 材料培养
莳萝蒿(Artemisiaanethifolia)的种子采自黄河三角洲。挑选籽粒饱满的种子播种于装有干净细砂的塑料盆内,当幼苗长到4—5个叶片时,选取生长一致幼苗进行液体通气培养。水培幼苗适应2 d后进行NaCl处理,方法是用含有NaCl的完全Hoagland营养液(pH值5.9)每天递增100 mmol/L NaCl,同一天达到处理终浓度(100、200、300、400 mmol/L NaCl),以防止盐激效应。以Hoagland营养液为对照(实际含有约2 mmol/L Na+)。为保持NaCl浓度恒定,每2d更换1次营养液。培养温室的昼夜温度为(30±2)℃/(23±2)℃,光周期15/9 h,白天最大光照强度约为1500 μmol m-2s-1,相对湿度40%—60%。NaCl处理7 d后,测定有关指标。
1.2 测定方法
1.2.1 整株鲜重和叶绿素含量的测定
到达终浓度的当天将整株植物从液体培养盆内取出,轻轻吸去根部表面的水分,分别称量叶片、茎和根的鲜重(FW)。整株鲜重=叶鲜重+茎鲜重+根鲜重,每个处理做10个重复。
选取莳萝蒿最幼功能叶测定叶绿素含量[14]。由于莳萝蒿是羽状深裂叶,因此以鲜重为单位表示其叶绿素含量。
1.2.2 叶片肉质化程度和细胞汁液渗透势的测定
用叶片肉质化程度表示叶片的含水量:首先称量叶片鲜重(FW),然后将鲜材料放入105 ℃的烘箱杀青10 min后,80 ℃烘干至恒重,称量叶片干重(DW)。
叶片肉质化程度=FW/DW×100%
每个处理做5个重复。
叶片液氮速冻后,放入注射器内室温下溶冰,然后挤出细胞汁液。细胞汁液渗透势用蒸汽压渗透压计(Vapor Pressure Osmometer 5520,Hansatech Instrument Ltd., UK)测定。每个处理做5个重复。
1.2.3 叶片光合参数的测定
莳萝蒿幼苗在20:00充分暗适应后,在黑暗条件下用HandyPEA (Plant Efficiency Analyser;Hansatech Instrument Ltd., UK)测定其最大光化学效率(maximal efficiency of PSⅡ photochemistry,Fv/Fm)。每个处理做5个重复。
光合速率(Pn)和气孔导度(Gs)采用英国PP system公司的Ciras- 1型光合测定系统测定。测定条件为:CO2浓度360 μmol/mol、相对湿度40%,光照1000 μmol m-2s-1,温度25 ℃。叶室中莳萝蒿羽状叶叶面积用Li- 3000A叶面积仪(美国LI-COR)测定。每个处理做5个重复。
1.2.4 根、茎、叶Na+含量的测定
将每株植物分成根、茎、叶3部分,分别烘干、称重。然后取0.1 g 干重的材料用马弗炉在500 ℃充分灰化,用浓硝酸溶解,然后蒸馏水稀释定容。Na+浓度用原子吸收光谱仪(Z- 8000型, Hitachi, Japan)测定。各器官中Na+含量用μmol/g 鲜重表示:
各器官中Na+总量=Na+含量×器官鲜重每个处理做5个重复。
1.2.5 叶片质外体和原生质体Na+含量的测定
质外体液体的收集参照Tetlow的方法稍加改动[15]。取一定数量的完整叶片,将叶片茎基端向下置于容积为40 mL的注射器中,注射器下端出液口套一容积为0.5 mL的Eppendorff管,放入超速冷冻离心机中4 ℃、400g离心10 min,避免细胞破裂。质外体液体顺着注射器收集到Eppendorf管中,收集到的质外体(apoplast)液体稀释后测定其Na+浓度(Ca)。方法1.2.2中得到的细胞汁液[其中含有质外体溶液和原生质体(protoplast)溶液两部分] 4000g离心10 min,取上清液用于Na+浓度的测定(CT)。参照Flowers的方法,假定质外体与原生质体体积比约为3∶97,可推算原生质体溶液的平均Na+浓度(Cp)[16]:
Cp=(CT×1-Ca×Va)/Vp
式中,Va= 1×3%,Vp=1× 97%。每个处理做5个重复。
Na+在质外体和原生质体中的分布比例计算公式分别为:
Pa=Ca×Va/(Ca×Va+Cp×Vp)
Pp=Cp×Vp/(Ca×Va+Cp×Vp)
1.2.6 液泡膜微囊的制备
液泡膜微囊的提取参考Ballesteros 等人的方法[17]。以不同盐浓度处理的莳萝蒿叶片为材料,用预冷重蒸水冲洗2—3次,用吸水纸吸去表面水分后,按材料∶匀浆缓冲液=1∶2加入预冷匀浆缓冲液,该缓冲液含有50 mmol/L Tricine-Tris (pH 7.5),3 mmol/L EGTA, 3 mmol/L MgSO4,0.5% PVP,2 mmol/L DTT,0.2 mmol/L PMSF,5% 甘油(Glycerol),并用甘露醇调至与叶片细胞汁液等渗。用匀浆机匀浆2次,四层纱布过滤。取滤液10000 r/min (Beckman, L- 80XP)离心20 min,一并除去线粒体及没有破碎的细胞和细胞壁碎片。取离心后的上清液小心铺在18%∶24%∶30%∶45%(质量分数)不连续蔗糖梯度溶液上(含5 mmol/L Hepes-Tris pH值7.5,1 mmol/L DTT),100000g离心2h,小心收集24% (质量分数) 界面上的膜微囊,来自24%界面的微囊富含液泡膜。用稀释液(含有3 mmol/L MgSO4, 50 mmol/L Hepes-Tris pH值7.5, 1 mmol/L PMSF,1 mmol/L DTT)稀释2—4倍,100,000×g 再离心30 min。离心后的沉淀用尖头细毛笔小心悬浮在贮藏液[含有40% 甘油 (glycerin),2 mmol/L DTT, 10 mmol/L Hepes-Tris (pH值=7.5)]中,用匀浆器混匀后分装于Eppendorf管中,液氮中冷冻后,-80 ℃超低温冰箱保存备用。以上操作均在0—4 ℃条件下完成。用考马斯亮蓝法测定提取液中蛋白质含量。
1.2.7 液泡膜H+-ATPase泵活性、Na+/H+逆向转运活性的测定
H+-ATPase泵活性和Na+/H+逆向转运活性测定均参照Ballesteros 等人的方法[17]。质子泵活性的测定的反应体系为2 mL,其中含有33 mmol/L Hepes-tris(pH值=7.5),50 mmol/L KCl,0.1 mmol/L (NH4)2MoO4,166 mmol/L 甘露醇,0.05%牛血清蛋白,3 mmol/L DTT,1.5 mmol/L ATP-Na2,膜微囊制剂蛋白量100 μg。25 ℃保温5 min,再加入浓度为1 mol/L的MgCl210 μL启动反应。用acridine orange作荧光染料,反应体系中acridine orange的终浓度为5 μmol/L。测定荧光猝灭的初速度(激发波长为495 nm,发散波长为525 nm)。以单位时间内荧光猝灭值占总荧光量的百分数(%quench/min)表示。
荧光猝灭达到最大且稳定时,加入15 μmol/L bafilomycin 20 μL终止V-H+-ATPase 活性,然后加入Na+(NaCl),使体系中Na+浓度为100 mmol/L,测定荧光恢复的初速度,以单位时间内荧光恢复值占总荧光量的百分数(%quench/min)表示Na+/H+逆向转运活性。每个处理的两种酶活性均做5个重复。
2 结果与分析
2.1 盐处理对莳萝蒿生长状况的影响
盐处理显著抑制了莳萝蒿的生长,随着盐处理浓度的升高,莳萝蒿整株鲜重显著下降。100、200、300、400 mmol/L NaCl条件下的整株鲜重分别为对照的87.9%、79.0%、72.3%、65.7% (图1)。地上部鲜重尤其是叶片鲜重下降最为显著,分别为对照的84.5%、73.0%、64.0%、51.9%。盐处理后,莳萝蒿的叶片数和叶片大小均显著小于对照,在300—400 mmol/L NaCl下,老叶有发黄枯萎现象;但功能叶片的叶绿素含量显著高于对照(图1),这是叶片生长受抑制后的常见表现。
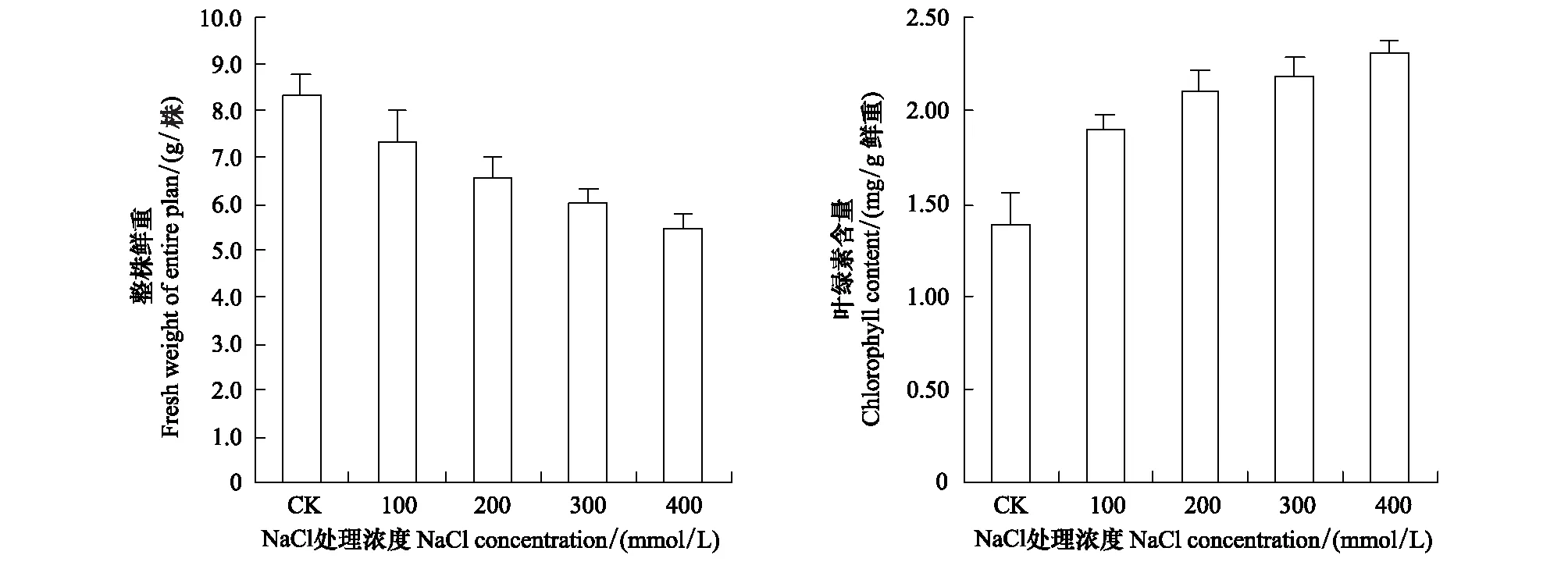
图1 盐处理对莳萝蒿整株鲜重(a)和叶绿素含量(b)的影响Fig.1 Effect of salt-treatment on fresh weight of individual Artemisia anethifolia plants (a) and chlorophyll content (b)
2.2 盐处理对莳萝蒿水分状况的影响
莳萝蒿地上部分主要为叶片,叶片的水分状况可以反映其生活状态。为了适应盐渍条件下的低渗环境,其叶片的细胞汁液渗透势也显著下降,以提高其吸水能力(图2)。400 mmol/L NaCl条件下莳萝蒿叶片细胞汁液渗透势达到对照的2.07倍。莳萝蒿叶片的肉质化程度却显著下降(图2)。400 mmol/L NaCl条件下,莳萝蒿叶片肉质化程度仅为对照的69.1%。

图2 盐处理对莳萝蒿叶片肉质化程度和细胞汁液渗透势的影响Fig.2 Effect of salt-treatment on leaf succulence degree and osmotic potential of leaf cell sap of Artemisia anethifolia plants
2.3 盐处理对莳萝蒿叶片光合功能的影响
盐处理后莳萝蒿叶片的光合功能显著下降(图3),100、200、300和400 mmol/L NaCl条件下的净光合速率分别为对照的70.6%、48.7%、40.5%和28.1%。气孔导度的下降趋势与净光合速率类似,而莳萝蒿叶片的PSⅡ最大光化学效率(Fv/Fm)不受盐处理影响(图3)。说明盐处理抑制莳萝蒿的光合功能与气孔因素有密切关系。
2.4 不同浓度盐处理条件下莳萝蒿各器官中Na+的含量与分布
盐处理后,莳萝蒿幼苗的各个器官中的Na+含量均显著增加,但地上部的Na+含量均显著高于根部,其中叶片的Na+含量最高(表1),说明莳萝蒿并不阻止Na+运往地上部。随着盐处理浓度的增加,叶片Na+含量增加幅度显著小于根和茎,如400 mmol/L NaCl条件下叶、茎、根中Na+含量分别为100 mmol/L NaCl条件下的2.04、3.16和2.66倍。因此,在高盐浓度下,莳萝蒿相对减缓了叶片Na+积累幅度。各器官中的Na+总量结果则显示(表2),在不同盐渍条件下,叶片均是莳萝蒿Na+分布的主要器官,分布比例在70%以上。但随着盐浓度的升高,叶片Na+总量所占的比例逐渐降低,而根和茎中Na+总量所占的比例逐渐升高。

表1 不同盐处理条件下莳萝蒿各器官中Na+含量(μmol/g鲜重)
表2 不同盐处理条件下莳萝蒿各器官Na+总量(μmol)与分布比例
Table 2 Total Na+content (μmol) and distribution ratio in different organs ofArtemisiaanethifoliaplants under different concentrations of NaCl

器官OrganNaCl处理浓度/NaClconcentrations(mmol/L)CK100200300400叶Leaf154.5±16.8(88.9)940.8±30.7(82.8)1019.5±43.6(76.3)1147.0±53.8(75.0)1260.9±53.6(72.9)茎Stem2.7±0.4(1.7)28.4±2.5(2.5)45.0±4.3(3.4)57.9±5.6(3.8)71.9±8.1(4.8)根Root16.6±1.0(9.4)167.3±16.7(14.7)270.4±23.9(20.3)323.5±21.6(21.2)331.1±28.9(22.3)

图3 盐处理对莳萝蒿叶片光合功能的影响Fig.3 Effect of salt-treatment on photosynthetic function of Artemisia anethifolia leaf
2.5 不同浓度盐处理条件下莳萝蒿叶片质外体和原生质体Na+浓度与分布
由于叶片是莳萝蒿积累Na+的主要器官,因此重点分析了叶片组织质外体和原生质体Na+浓度与分布(图4)。随着盐处理浓度的增加,质外体和原生质体中的Na+浓度均显著上升。但无论对照还是盐处理条件下,原生质体中的Na+平均浓度均显著高于质外体,CK、100、200、300和400 mmol/L NaCl条件下原生质体Na+浓度分别为质外体的1.8、1.7、1.5、1.3和1.2倍(图4),说明叶肉细胞可以主动吸收和积累Na+,但随着盐处理浓度的升高,原生质体中的Na+相对浓度逐渐减小。
叶片积累的Na+主要分布在原生质体中,在不同盐浓度处理下,原生质体的Na+总量均占叶片组织Na+总量的98%左右。CK、100、200、300和400 mmol/L NaCl条件下质外体的Na+总量分别占叶片组织Na+总量1.7%、1.8%、2.1%、2.3%和2.4%(图4),从这组数据也可以看出在高盐浓度下,质外体积累Na+的数量相对增加。
2.6 不同浓度盐处理条件下莳萝蒿叶片液泡膜V-H+-ATPase与Na+/H+逆向转运活性
盐处理莳萝蒿叶片的液泡膜V-H+-ATPase质子泵活性均显著大于对照,100、200、300和400 mmol/L NaCl处理的V-H+-ATPase质子泵活性分别为对照的1.3、1.3、1.4和1.5倍。但随着盐处理浓度的升高,V-H+-ATPase质子泵活性的增加幅度放缓。如200、300、400 mmol/L NaCl条件下V-H+-ATPase质子泵活性分别为100 mmol/L NaCl的1.07、1.15、1.17倍。300—400 mmol/L NaCl条件下,V-H+-ATPase质子泵活性几乎不再增加(图5)。
莳萝蒿叶片的液泡膜Na+/H+逆向转运活性的特点是在对照条件下活性较低,盐处理后活性迅速增大,约为对照的4—7倍。同样,随着盐处理浓度的升高,Na+/H+逆向转运活性的增加幅度放缓。如200、300、400 mmol/L NaCl条件下的Na+/H+逆向转运活性分别为100 mmol/L NaCl的1.22、1.34、1.38倍。300—400 mmol/L NaCl条件下,Na+/H+逆向转运活性也几乎不再增加(图5)。

图4 不同盐处理条件下莳萝蒿叶片质外体和原生质体平均Na+浓度(μmol/mL)与分布比例Fig.4 Na+ concentration and distribution percentage in apoplast and protoplasts of Artemisia anethifolia leaf tissue under different concentrations of NaCl

图5 不同盐处理条件下莳萝蒿液泡膜V-H+-ATPase质子泵活性和Na+/H+逆向转运活性的变化Fig.5 Changes in V-H+-ATPase proton pump activity and Na+/H+ antiporter activity of Artemisia anethifolia leaves under different concentrations of NaCl
3 讨论
莳萝蒿具有较强的耐盐能力,能够在400 mmol/L NaCl条件下存活并生长。但其生长量随着盐处理浓度的升高而逐渐减小(图1),其主要原因可能与叶片的光合速率迅速下降有关。由于植物光合系统中所有的酶均对Na+非常敏感[8,18],而莳萝蒿叶片的PSⅡ光化学活性未受到高盐伤害(图3),叶绿素含量还显著增加(图1),说明莳萝蒿能够对其光系统进行有效地保护。伴随着光合速率的下降,莳萝蒿叶片的气孔导度也显著下降(图3),因此推测盐处理抑制莳萝蒿的净光合速率主要是由于气孔因素引起的。
盐生植物保护细胞质免受伤害的共同机制是减少Na+在细胞质中的积累,避免Na+毒害[19- 20]。避免细胞质Na+毒害的方式主要有3种:拒盐、泌盐和稀盐。莳萝蒿既没有把盐分阻隔在地下部,也不能通过大量吸水和快速生长稀释盐分。因此,莳萝蒿既不是拒盐植物,也不是喜盐植物,也没有泌盐结构,它的耐盐机制还未见报道。从本文结果可以看出,莳萝蒿的耐盐机制是将吸收到体内的盐分主要积累在地上部尤其是叶片中,叶片中的Na+含量占整株植物的70%以上(表2),也是Na+含量最高的器官(表1)。叶片中的盐分则97%以上Na+均积累在原生质体中(图4),由于细胞质中的酶对Na+敏感,所以细胞质中的Na+浓度一般不超过10 mmol/L[5,20],因此莳萝蒿叶片中的Na+应该主要积累在叶片细胞的液泡中[19,21]。莳萝蒿原生质体中的Na+浓度高于质外体也证明莳萝蒿区域化Na+的方式不是由细胞质外排到质外体,而是区域化到液泡内。
Na+区域化是植物耐盐的共同机制,其中Na+在液泡中的区域化最受关注[21]。液泡膜上两种重要的酶参与了Na+在液泡内的区域化:V-H+-ATPase和Na+/H+逆向转运蛋白,其中V-H+-ATPase为Na+逆向转运建立跨液泡膜的质子能量梯度,Na+/H+逆向转运蛋白是Na+区域化到液泡的载体[2- 5]。经测定,盐处理后二者的活性协同上调(图5),说明盐处理后莳萝蒿增强了Na+区域化到液泡的能力,两种酶活性的增加可能是由于蛋白质的表达增加引起的[22- 23]。不过,当盐处理浓度达到300—400 mmol/L NaCl时,两种酶的活性均不再增加,说明此时莳萝蒿的Na+区域化能力已经达到了极限。在此浓度下,莳萝蒿的基部老叶出现明显枯萎现象,是由于盐分优先积累在老叶,保护幼叶所致。图5的结果显示莳萝蒿的Na+/H+逆向转运活性均是组成性表达的,这一特征与真盐生植物类似[24],耐盐甜土植物的Na+/H+逆向转运活性往往是盐诱导表达的[21]。
由于Na+是通过蒸腾流进入植物体的,莳萝蒿为了减少Na+的吸收所采取的方式是关闭气孔(图3),降低蒸腾。但这一方式不可避免地影响了植物光合作用的气体交换,从而抑制了自身的生长速度。降低蒸腾是植物在盐渍渗透胁迫条件下的普遍反应。但真盐生植物如盐地碱蓬(SuaedasalsaLinn.)在盐渍条件下能够利用Na+降低细胞汁液渗透势[11],且能大量吸收水分,快速生长而稀释盐分[13]。莳萝蒿也能利用Na+降低细胞汁液渗透势(图2)。但其叶片含水量却随着盐处理浓度的升高而下降,这可能是莳萝蒿与真盐生植物的主要区别。有文献认为真盐生植物的吸水快速生长机制与盐诱导增加水孔蛋白表达有关[25]。莳萝蒿叶片盐处理后含水量下降很可能是因为其缺乏诱导质膜水孔蛋白大量表达的机制。分子生物学实验证明水孔蛋白活性增加可以提高植物光合能力和生物产量[26],因此水孔蛋白可能在真盐生植物的耐盐性及快速生长机制中具有重要作用。
总之,众多实验已经证明Na+区域化可以赋予植物耐盐性,是耐盐植物的共同特征。但是Na+区域化并不能使莳萝蒿避免盐抑制生长现象,并未具备类似真盐生植物适应盐渍环境而快速生长的能力。所以Na+/H+逆向转运活性是植物适应盐渍环境的必需条件而不是充分条件。
[1] Zhao K F, Li F Z. The halophytes in China. Beijing: Science Publishing House, 1999: 289- 292.
[2] Maeshima M. Tonoplast transporters: Organization and function. Annual Review of Plant Physiology and Plant Molecular Biology, 2001, 52(1): 469- 497.
[3] Mansour M M F, Salama K H A, Al-Mutawa M M. Transport proteins and salt tolerance in plants. Plant Science, 2003, 164(6): 891- 900.
[4] Hunte C, Screpanti E, Venturi M, Rimon A, Padan E, Michel H. Structure of a Na+/H+antiporter and insights into mechanism of action and regulation by pH. Nature, 2005, 435(7046): 1197- 1202.
[5] Sun J, Chen S L, Dai S X, Wang R G, Li N Y, Shen X, Zhou X Y, Lu C F, Zheng X J, Hu Z M, Zhang Z K, Song J, Xu Y. Ion flux profiles and plant ion homeostasis control under salt stress. Plant Signaling Behavior, 2009, 4(4): 261- 264.
[6] Ohta M, Hayashi Y, Nakashima A, Hamada A, Tanaka A, Nakamura T, Hayakawa T. Introduction of a Na+/H+antiporter gene fromAtriplexgmeliniconfers salt tolerance to rice. FEBS Letters, 2002, 532(3): 279- 282.
[7] Chinnusamy V, Jagendorf A, Zhu J K. Understanding and improving salt tolerance in plants. Crop Science, 2005, 45(2): 437- 448.
[8] Yang Q, Chen Z, Zhou X, Yin H, Li X, Xin X, Hong X, Zhu J K, Gong Z. Overexpression of SOS (Salt Overly Sensitive) genes increases salt tolerance in transgenic Arabidopsis. Molecular Plant, 2009, 2(1): 22- 31.
[9] Oh D H, Lee S Y, Bressan R A, Yun D J, Bohnert H J. Intracellular consequences of SOS1 deficiency during salt stress. Journal of Experimental Botany, 2010, 61(4): 1205- 1213.
[10] Daines R J, Gould A R. The cellular basis of salt tolerance studied with tissue cultures of the halophytic grassDistichlisspicata. Journal of Plant Physiology, 1985, 119(3): 269- 280.
[11] Qiu N W, Chen M, Guo J R, Bao H Y, Ma X L, Wang B S. Coordinate up-regulation of V-H+-ATPase and vacuolar Na+/H+antiporter as a response to NaCl treatment in a C3halophyteSuaedasalsa. Plant Science, 2007, 172(6): 1218- 1225.
[12] Zhao F Y, Guo S L, Zhang H, Zhao Y X. Expression of yeastSOD2 in transgenic rice results in increased salt tolerance. Plant Science, 2006, 170(2): 216- 224.
[13] Li WQ, Liu X J, Zhao K F, Liu H L. Growth, development and ions distribution of three halophytes under salt stress. Chinese Journal of Eco-Agriculture, 2006, 14(2): 49- 52.
[14] Zhang Q D. Several methods of determination of chlorophyll. Chinese Bulletin of Botany, 1985, 3(5): 60- 64.
[15] Tetlow I J, Farrar J F. Apoplastic sugar concentration and pH in barley leaves infected with brown rust. Journal of Experimental Botany, 1993, 44(5): 929- 936.
[16] Flowers T J, Yeo A R. Ion relations of plants under drought and salinity. Australian Journal of Plant Physiology, 1986, 13(1): 75- 91.
[17] Ballesteros E, Blumwald E, Donaire J P, Belver A. Na+/H+antiport activity in yonoplast vesicles isolated from sunflower Roots induced by NaCl stress. Physiologia Plantarum, 1997, 99(2): 328- 334.
[18] Zhang L R, Xing D. Rapid determination of the damage to photosynthesis caused by salt and osmotic stresses using delayed fluorescence of chloroplasts. Photochemistry and Photobiology Science, 2008, 7(3): 352- 360.
[19] Munns R, Tester M. Mechanisms of salinity tolerance. Annual Review of Plant Biology, 2008, 59(1): 651- 681.
[20] Zhu J K. Regulation of ion homeostasis under salt stress. Current Opinion in Plant Biology, 2003, 6(5): 441- 445.
[21] Qiu N W, Yang H B, Wang B S. The Na+/H+antiporter and its relation to salt tolerance in plants. Plant Physiology Communications, 2001, 37(3): 260- 264.
[22] Han N, Shao Q, Lu C M, Wang B S. The leaf tonoplast V-H+-ATPase activity of a C3halophyteSuaedasalsais enhanced by salt stress in a Ca-Dependent mode. Journal of Plant Physiology, 2005, 162(3): 267- 274.
[23] Queirós F, Fontes N, Silva P, Almeida D, Maeshima M, Gerós H, Fidalgo F. Activity of tonoplast proton pumps and Na+/H+exchange in potato cell cultures is modulated by salt. The Journal of Experimental Botany, 2009, 60(4): 1363- 1374.
[24] Hasegawa P M, Bressan R A, Zhu J K, Bohnert H J. Plant cellular molecular responses to high salinity. Annual Review of Plant Physiology and Plant Molecular Biology, 2000, 51(1): 463- 499.
[25] Qi C H, Chen M, Song J, Wang B S. Increase in aquaporin activity is involved in leaf succulence of the euhalophyteSuaedasalsaunder salinity. Plant Science, 2009, 176(2): 200- 205.
[26] Uehlein N, Lovisolo C, Siefritz F, Kalenhoff R. The tobacco aquaporin NtAQP1 is a membrane CO2pore with physiological functions. Nature, 2003, 425(6959): 734- 737.
参考文献:
[1] 赵可夫, 李法曾. 中国盐生植物. 北京: 科学出版社, 1999: 289- 292.
[13] 李伟强, 刘小京, 赵可夫, 刘海亮. NaCl 胁迫下3 种盐生植物生长发育及离子在不同器官分布特性研究. 中国生态农业学报, 2006, 14(2): 49- 52.
[14] 张其德. 测定叶绿素的几种方法. 植物学通报, 1985, 3(5): 60- 64.
[21] 邱念伟, 杨洪兵, 王宝山. Na+/H+逆向转运蛋白及其与植物耐盐性的关系. 植物生理学通讯, 2001, 37(3): 260- 264.
Na+compartmentation and physiological characteristics ofArtemisiaanethifoliain adaptation to saline environment
QIU Nianwei1, YANG Cuicui1, LU Zhengke1, LI Zunning1, YUE Xianjun1, CHENG Xiuxiu1, XU Yingying1, ZHOU Feng2,*
1CollegeofLifeSciences,QufuNormalUniversity,Qufu,Shandong273165,China2SchoolofBiochemicalandEnvironmentalEngineering,NanjingXiaozhuangUniversity,Nanjing,Jiangsu211171,China
Artemisiaanethifolia(Compositae) is a halophyte that is widely distributed in northern areas of China. Studying the mechanisms by which this plant adapts to high levels of salt will increase our understanding of salt adaptation in vascular plants.A.anethifoliaplants were treated with 0, 100, 200, 300, and 400 mmol/L NaCl for 7 d, respectively. Then, the differences in growth and physiology were compared between salt-treated and controlA.anethifoliaplants. In particular, the Na+accumulation levels and Na+compartmentation patterns inA.anethifoliaplants were analyzed in detail. Although theA.anethifoliaplants were able to survive under 400 mmol/L NaCl, salt-treated plants showed lower fresh weight as the NaCl concentration increased, indicating that plant growth was inhibited by salt. The leaves ofA.anethifoliashowed an increased capacity for osmotic adjustment but a decreased degree of leaf succulence with increasing NaCl concentrations. Photosynthetic analyses showed that there was a gradual decline in net photosynthetic rate and stomatal conductance of salt-treated leaves with increasing NaCl concentrations. However, the maximal efficiency of photosystem II photochemistry (Fv/Fm) inA.anethifolialeaves was not inhibited by salt, and the chlorophyll content even increased with increasing salt concentrations. These observations suggested that the decreased photosynthetic rate was due to stomatal factors, rather than damage to the components of the photosynthetic machinery. The Na+content inA.anethifoliaplants tended to increase with increasing salt concentrations. The Na+content in leaves, stems, and roots ofA.anethifoliawas 321.4, 242.1, and 182.3 μmol/g FW, respectively, in the 400 mmol/L NaCl treatment. More than 70% of the Na+absorbed by salt-treatedA.anethifoliaplants accumulated in their leaves, and approximately 98% of the Na+that accumulated in leaves was localized in leaf protoplasts. The average Na+concentration in protoplasts ofA.anethifolialeaf tissue was 1.2—1.8 times that in the apoplast. These results indicate that efficient Na+compartmentation occurred inA.anethifolialeaf cells. The V-H+-ATPase proton pump activity of salt-treated leaves was 30%—50% higher than that of control leaves, and the tonoplast Na+/H+antiporter activity of salt-treated leaves was 4—7 times that in leaves of control plants. These findings suggested thatA.anethifoliaplants have a strong ability to compartmentalize Na+in the vacuole. As the concentrations of salt increased, the Na+distribution ratio in leaves decreased. Likewise, the range of increased V-H+-ATPase proton pump activity and tonoplast Na+/H+antiporter activity decreased as the salt concentration increased. Na+compartmentation plays an important role in the salt tolerance ofA.anethifoliaplants, because it protects their photosystems and results in lower osmotic potential in the leaf cells. However, Na+compartmentation could not prevent the decrease in the degree of leaf succulence and the photosynthetic activity ofA.anethifolialeaves under highly saline conditions, and so their growth was inhibited by salt. These results suggest that Na+compartmentation is necessary but not sufficient for adaptation ofA.anethifoliato a saline environment.
Artemisiaanethifolia; Na+compartmentation; growth; photosynthesis; adaption
江苏省自然科学基金青年基金项目(BK2012073); 国家重点实验室开放课题(2011KF07)
2013- 01- 31; 网络出版日期:2014- 03- 13
10.5846/stxb201301310204
*通讯作者Corresponding author.E-mail: zfibcas@163.com
邱念伟,杨翠翠, 卢正珂, 李遵宁, 岳贤军, 程秀秀, 许莹莹, 周峰.莳萝蒿适应盐渍环境的Na+区域化方式和生理特征.生态学报,2014,34(21):6147- 6155.
Qiu N W, Yang C C, Lu Z K, Li Z N, Yue X J, Cheng X X, Xu Y Y, Zhou F.Na+compartmentation and physiological characteristics ofArtemisiaanethifoliain adaptation to saline environment.Acta Ecologica Sinica,2014,34(21):6147- 6155.
猜你喜欢
江苏安全生产(2022年1期)2022-03-09
煤气与热力(2021年11期)2021-12-21
矿产勘查(2020年5期)2020-12-25
中国农业科技导报(2020年1期)2020-03-15
中国真菌学杂志(2019年1期)2019-01-23
食用菌(2017年5期)2017-10-19
中国劳动关系学院学报(2016年1期)2016-09-26
大连工业大学学报(2015年4期)2015-12-11
特产研究(2015年1期)2015-04-12
农业科技与装备(2014年6期)2014-11-28
