
植物生长调节剂混用对水稻同化物及三种保护酶的影响
2014-08-06 03:29易书佳孔祥清徐宇付迪
黑龙江八一农垦大学学报 2014年6期
易书佳,孔祥清,徐宇,付迪
(黑龙江八一农垦大学农学院,大庆 163319)
黑龙江省位于中国北部地区,地处寒温带地区,属我国低温冷害高发地区,通过李文亮[1]等对黑龙江1971年到2005年温度分析得出,全省各地均频繁发生不同程度低温冷害,南部地区低温冷害发生频率高于北部地区。逆境胁迫能够影响植物生理系统,影响植株生长,严重时造成植株死亡。植物生长调节剂是一类能够调控植物生长、发育和衰老进程的活性物质[2-4],由于植物生长调节剂可以直接对植物激素系统调控,并具有用量小、速度快、效益高、残毒少、可对植物的外部性状与内部生理过程进行双调控等多种作用,使之在近代植物生理学及农业科学具有十分重要的意义。孔祥森研究发现,植物外施DTA-6能够有效提高水稻幼苗体内保护酶活性,降低有害物质积累[5]。甲哌钅翁是一种温和、吸收快抑制植物茎叶疯长的新型植物生长调节剂,能显著提高植物幼苗叶片中SOD、CAT 的活性提高植物叶片中可溶性糖含量增加植物抗寒性,并有效抑制幼苗的顶端优势,延缓植株地上部分的纵向生长[6]。细胞分裂素有助于促进侧芽分化,解除顶端优势,加快腋芽发育,延缓叶片衰老,提高叶片中保护酶活性,提高植物抗逆性[7-8]。烯效唑作为一种能够使水稻矮化增产,促进植株根系生长,提高作物光合效率等作用的植物生长调节剂而被广泛使用[9]。实验根据甲哌钅翁、细胞分裂素及烯效唑其作用机理以及作用效果各不相同,研究三种植物生长调节剂混配在不同时期喷施后对水稻生理指标的影响,为水稻提高抗逆性提供依据。
98%甲哌钅翁(南通施壮化工有限公司)
5%烯效唑乳油(青岛百禾丰华生物工程有限公司)
1 材料与方法
1.1 实验材料
供试品种:龙庆1 号
实验地点:大庆市黑龙江八一农垦大学试验地
供试药剂:
98%细胞分裂素(河南新力源农业科技有限公司)
1.2 试验方法
采用双因素裂区设计,小区对比的方法,以不同药剂及剂量处理,设不同生长调节剂及清水对照5个水平;分别在水稻分蘖期(A)和水稻拔节期(B)喷施,每个处理3 次重复,共计10 各小区,每小区6 行,行长15 m,小区面积为30 m2,插秧规格为30 cm×13 cm 如表1。施肥,灌水等其他情况均同于正常管理。

表1 试验设计Table 1 The treatment design
1.3 测定项目与方法
分别于施药后每间隔15 d 取样一次,每小区五点取样,每样点取样三穴。
1.3.1 保护酶活性测定
超氧化物歧化酶(SOD)活性测定采用氮蓝四唑法[10-11];
过氧化物酶(POD)活性测定采用愈创木酚显色法[12-13];
过氧化氢酶(CAT)活性测定采用紫外吸收法[14-15]。
1.3.2 总糖及还原糖含量测定
总糖含量测定采用蒽酮比色法[16],还原糖含量测定采用3,5-二硝基水杨酸法[17]。
1.4 数据统计分析
数据利用Micosoft Excel 2010 进行图表制作;用DPS 软件进行数据显著性分析。
2 结果与分析
2.1 不同时期各处理对水稻叶片还原糖、总糖含量的影响
2.1.1 对叶片还原糖含量影响
不同时期喷施不同植物生长调节剂水稻叶片还原糖含量影响如表2 所示,在水稻分蘖期喷施调节剂的各时期各处理还原糖含量均高于对照组,喷药15 d 后,处理A1,A2,A3,A4,CK 叶片中还原糖含量A1>A4>A2>A3>CK,处理A1、A4 极显著高于对照。喷药30 d 后,处理A1,A2,A3,A4,CK 叶片中还原糖含量A2>A4>A3>A1>CK,处理A2、A4、A3 极显著高于对照。喷药45 d 后,处理A1,A2,A3,A4,CK叶片中还原糖含量A3>A2>A1>A4>CK,均极显著高于对照。
在水稻拔节期喷施调节剂的各时期各处理还原糖含量均高于对照组,喷药15 d 后,处理B1,B2,B3,B4 叶片中还原糖含量B2>B1>B4>B3>CK,处理B1、B2 极显著高于对照。喷药30 d 后,处理B1,B2,B3,B4,CK 叶片中还原糖含量B1>B2>B3>B4>CK,处理B1 极显著高于对照。
2.1.2 叶片总糖含量变化
不同时期喷施不同植物生长调节剂对水稻叶片中总糖含量影响如表3 所示在水稻分蘖期喷施调节剂的各时期各处理总糖含量均显著高于对照组,处理A1,A2,A4 在喷药后15 d 呈现出明显增高,之后逐渐下降。在水稻拔节期喷施调节剂的各处理在喷药后15 d 总糖含量极显著高于对照,B1>B2>B4>B3>CK。喷药后30 d,B4>B1>B2>CK>B3,处理B4 总糖含量低于对照。
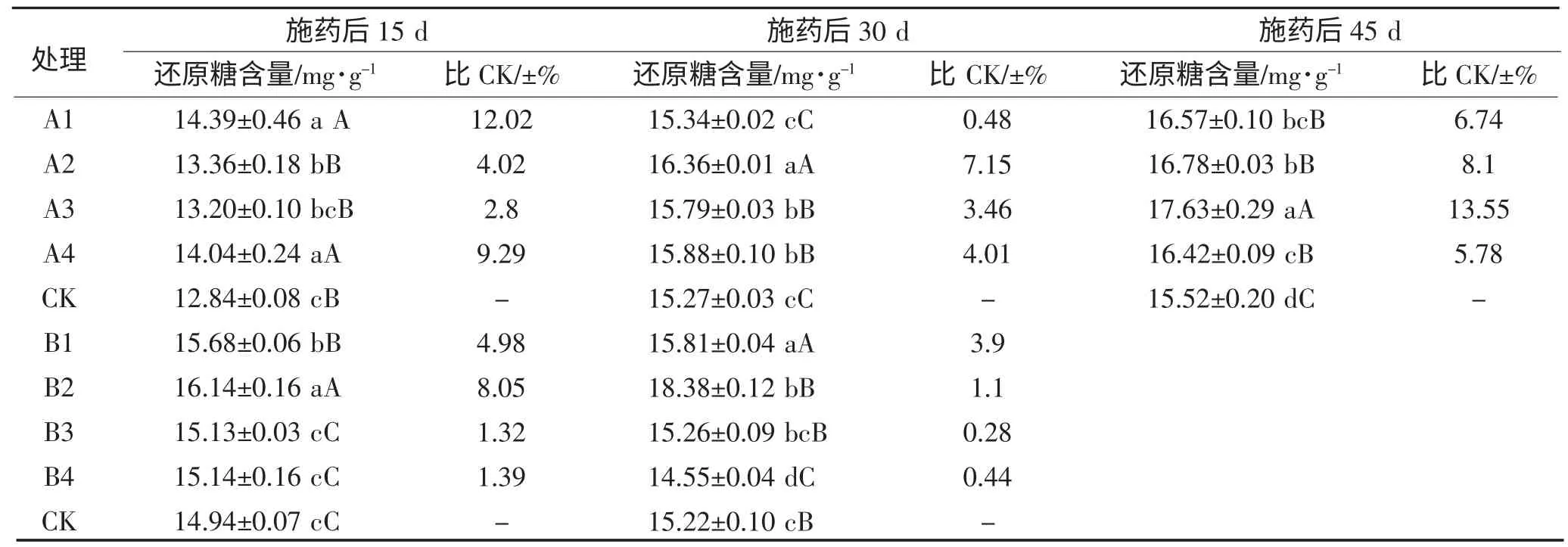
表2 各处理水稻叶片还原糖含量影响Table 2 The effects of soluble sugar of rice leaf under different treatment

表3 各处理对水稻叶片总糖含量影响Table 3 The effects of total sugar of rice leaf under different treatment
2.2 不同时期各处理水稻叶片SOD、POD、CAT 活性含量的变化
2.2.1 叶片中SOD 活性含量变化
水稻叶片在不同时期喷施不同植物生长调节剂后叶片中SOD 活性变化如表4 所示在水稻分蘖期喷施调节剂的各时期各处理SOD 活性均高于对照组,处理A1,A2,A4 在各个时期的活性均以相同增长率高于对照,而处理A3 在处理后并未立即体显出显著增长,但在整个时期呈现快速增长的态势。在水稻拔节期喷施调节剂的各时期各处理SOD 活性均高于对照组,喷药后15 d,各处理SOD 活性差异较小,喷药后30 d,B2,B4 对叶片SOD 活性的影响较大其SOD活性极显著高于对照和其他处理。
2.2.2 叶片中POD 活性含量变化
水稻叶片在不同时期喷施不同植物生长调节剂后叶片中POD 活性变化如表5 所示,在水稻分蘖期喷施后15 d 处理A1,A2 的POD 活性显著高于其他处理及对照,而处理A3,A4 在喷药15 d 后POD 活性略低于对照,但在整个时期呈现快速增长的态势,处理A3 在喷药后45 d POD 活性提高最为显著,而处理A4 则在喷药后30 d POD 活性提高最为显著。在水稻拔节期喷施调节剂,喷药后15 d,各处理POD活性均极显著高于对照,处理B1,B4 的POD 活性极显著高于对照和其他处理,喷药后30 d,B1,B2 对叶片POD 活性的影响较大,其POD 活性极显著高于对照和其他处理。
2.2.3 叶片中CAT 活性含量变化
水稻叶片在不同时期喷施不同植物生长调节剂后叶片中CAT 活性变化如图5 所示在水稻分蘖期喷施调节剂的各时期各处理SOD 活性均高于对照组,在喷药15 d 后,各处理SOD 活性差异较小,处理A1,A2,A3,A4 在各个时期的活性均极显著高于对照。在水稻拔节期喷施调节剂的各时期各处理CAT活性均高于对照组,喷药后15 d 处理B2>B1>B3>B4>CK,喷药后30 d 处理B3>B1>B2>B4>CK。

表4 各处理水稻叶片SOD 活性影响Table 4 The effects of SOD activity of rice leaf under different treatment

表5 不同时期植物生长调节剂对水稻叶片POD 活性影响Table 5 The effects of POD activity of rice leaf under different treatment

表6 不同时期植物生长调节剂对水稻叶片CAT 活性影响Table 6 The effects of CAT activity of rice leaf under different treatment
3 结论与讨论
通过不同时期施用不同植物生长调节剂,使水稻叶片中总糖、还原糖含量以及保护酶活性发生明显变化。处理后水稻叶片中各项指标均高于对照。在水稻分蘖期喷施效果明显优于水稻拔节期喷施药剂。
叶片中可溶性糖增加可降低细胞质凝固点,增加植物抗寒性。水稻还原糖含量呈上升趋势认为水稻喷施98%细胞分裂素15.10 g·hm-2+5%烯效唑乳油150 mL·hm-2后生长旺盛,新陈代谢快。
在水稻叶片中主要光合产物以可溶性糖的形式存,并在叶片中暂时贮藏。在水稻生育后期叶片中的光合产物向籽粒库运输进而合成淀粉[18]。喷施98%细胞分裂素15.15 g·hm-2+98%甲哌钅翁30.30 g·hm-2、98%细胞分裂素30.30 g·hm-2+98%甲哌钅翁60.60 g·hm-2、98%细胞分裂素15.15 g·hm-2+5%烯效唑乳油300 mL·hm-2能够使水稻叶片可溶性总糖含量在短时间内迅速升高加强水稻光合产物。
SOD、POD、CAT 在植物体内通过互相协调配合,清除植物体内活性氧,从而阻止植物体内有害物质积累,达到保护作用,保护酶含量增加有助于增强植物抗逆性。水稻茎秆强度受木质素含量影响,POD 酶参与木质素形成,POD 酶活性影响植物抗倒伏能力[19]。水稻叶片喷施药剂后,叶片中SOD、POD、CAT 的活性呈现持续升高的趋势。在分蘖期喷药后30~45 d 叶片中POD、CAT 含量显著增高,98%细胞分裂素15.15 g·hm-2+ 98%甲哌钅翁30.30 g·hm-2和98%细胞分裂素30.30 g·hm-2+5%烯效唑乳油150 mL·hm-2能够提高水稻叶片中保护酶活性从而增强水稻抗逆性及耐受性[20],增加水稻木质化。
[1]李文亮,张冬有,张丽娟.黑龙江省低温冷害发生规律及预测研究[J].灾害学,2008,23(4):30-35.
[2]杨建昌,王志琴,朱庆森.外源植物激素对水稻光合能力与产量的影响[J].江苏农业研究,1995,16(1):27-31.
[3]翟丙年,郑险峰,杨岩荣,等.植物生长调节物质的研究进展[J].西北植物学报,2003,23(6):1069-1075.
[4]王嘉宇,徐正进.外源赤霉素对不同穗型水稻穗部性状的影响[J].沈阳农业大学学报,2005,36(1):18-20.
[5]孔祥森.DTA-6 浸种对水稻种子发芽及幼苗生长的影响[J].黑龙江八一农垦大学学报,2008,20(4):9-11.
[6]罗立津,徐福乐,洪淑珠.甲哌钅翁对甜椒幼苗抗寒性的诱导作用研究[J].农药学学报,2010,12(2):142-148.
[7]王三根.细胞分裂素在植物抗逆和延衰中的作用[J].植物学通报,2000,17(2):121-126.
[8]赵小辉,朱乾华,贺薇,等.D-酪氨酸荧光猝灭法同时测定6-苄基腺嘌呤和糠氨基嘌呤[J].农药,2013,52(3):198-201.
[9]王熹,俞美玉,陶龙兴.烯效哇对稻苗的生理影响[J].中国水稻科学,1994,8(1):15-20.
[10]王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[11]李玲.植物生理学模块实验指导[M].北京:科学出版社,2009.
[12]张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2004.
[13]李玲.植物生理学模块实验指导[M].北京:科学出版社,2009.
[14]王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[15]刘萍,李明军.植物生理学实验技术[M].北京:科学出版社,2007.
[16]张志良.植物生理学实验指导[M].北京:高等教育出版社,2001.
[17]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[18]王书丽,郭天财,王晨阳,等.两种筋力型小麦叶、粒可溶性糖含量及与籽粒淀粉积累的关系[J].河南农业科学,2005(4):12-15.
[19]严重玲,洪业汤,林鹏,等.酸雨胁迫下稀土元素对菠菜膜保护系统的作用[J].生态学报,1999,19(4):543-545.
[20]欧阳西荣,林彰文.种子处理对玉米幼苗抗低温能力的影响[J].种子,2004,23(6):26-29.
猜你喜欢
信息记录材料(2022年3期)2022-05-17
今日农业(2021年13期)2021-11-26
农家参谋(2020年6期)2020-07-29
现代园艺(2018年3期)2018-02-10
科技与创新(2016年22期)2017-03-30
科技视界(2016年20期)2016-09-29
特产研究(2016年3期)2016-04-12
中国继续医学教育(2015年5期)2016-01-07
中国合理用药探索(2011年3期)2011-03-20
祝您健康(1999年5期)1999-12-25
