
辽宁仙人洞种子植物区系及与其他山地的相似性分析
2014-08-02 03:54:17黄国辉杨允菲
东北师大学报(自然科学版) 2014年4期
刘 利,黄国辉,张 梅,杨允菲
(1.辽东学院农学院,辽宁 丹东 118003;2.东北师范大学草地科学研究所,植被生态科学教育部重点实验室,吉林 长春 130024)
辽宁仙人洞种子植物区系及与其他山地的相似性分析
刘 利1,黄国辉1,张 梅1,杨允菲2
(1.辽东学院农学院,辽宁 丹东 118003;2.东北师范大学草地科学研究所,植被生态科学教育部重点实验室,吉林 长春 130024)
基于植物地理学理论,运用聚类模型、排序分析和最小生成树等数量方法,对辽宁仙人洞自然保护区植物区系地理组成、与其他山系植物区系的相似性进行了分析.结果表明,仙人洞自然保护区野生种子植物物种组成丰富,共有96科362属764种.被子植物占物种总数量的97.64%,其中双子叶植物占82.32%;属的地理成分以温带成分为主,共有236属,占总属数的74.67%,处于中国-日本森林植物亚区暖温带—温带的过渡范围内,划归于华北植物区系平原山地亚地区.在与之进行比对研究的16个山体中,仙人洞自然保护区种子植物属的分布区类型与凤凰山、千山、太行山、五台山、小五台山和东灵山有着较大的相似性,而纬度是影响植物地理分布的重要因素.
种子植物;区系谱;聚类分析;相似关系;辽东半岛
物种分布格局受水热条件及一些潜在生态因子的共同控制,同时也是物种演化历史、地理结构及环境变化等许多因素共同作用的结果[1],因此,植物区系的分布格局以及相互关系一直是生态学者关注的热点问题[2-5].仙人洞保护区位于辽东半岛的南部地区,南临黄海、北靠千山,物种组成具有东北向华北过渡交汇的特点.保护区内山势险峻、峰峦起伏,分布有大面积的前震旦系假岩溶地貌景观,在物种和地质、地貌等方面都有特殊的保护和科研价值.以往的研究多关注于生物多样性、生态系统功能等方面[6-8],而对物种区系方面的研究尚未见报道.本文在实地调查和参考相关文献[9-15]的基础上,对仙人洞保护区种子植物区系组成、属的分布区类型结构及与其他山体的相似性进行了分析,以揭示辽东半岛和山东半岛及华北植物区系的关系,为自然保护区的科学管理及当地生物资源的可持续开发利用提供科学支撑和参考依据.
1 研究区自然概况
辽宁仙人洞国家级自然保护区(124°53′24″~124°3′30″E,39°54′00″~40°03′00″N)位于辽东半岛庄河市境内的北部山区,地质构造属于中朝板块北东部南缘,地貌地形简单,大部分属于中切割侵蚀构造尖顶状低山,局部沿水系侵蚀堆积为山间峡谷区.保护区景色奇特,洞壁耸生,怪石嶙峋,河流迂回曲折.由于地处暖温带湿润季风气候区,夏季炎热降水多,冬季寒冷降雪少,春秋两季气候凉爽,雨热同期,具有一定的海洋性气候特点,年平均气温8.7℃,无霜期181.9 d,年平均日照时数约2 383.6 h;保护区内森林茂密,年降水量约1 000 mm,年均相对湿度69%.保护区地处东北和华北植物区系的交错地带,物种组成丰富.东北区系代表种有核桃楸(Juglansmandshurica)、黄檗(Phellodendronamurense)等;华北区系代表种有赤松(Pinusdensiflora)、麻栎(Quercusacutissima)、油松(Pinustabulaeformis)、玉铃花(Styraxobassis)等,赤松-麻栎林是最有代表的植物群落,适应性强,比较稳定.另有海州常山(Clerodendrumtrichotomum)、天女木兰(Magnoliasieboldii)等10余种珍稀植物物种,形成了独特的植物群落.
2 研究方法
2.1 区系成分及其分布区类型的确定与统计
对调查和收集的种子植物名录[9-15],以门为单位分别统计裸子植物和被子植物的科、属、种,再将被子植物以纲为单位分别统计双子叶植物和单子叶植物的科、属、种.由于同属物种在系统发育上具有较为一致的起源和进化趋势,同时在进化过程中,随着地理环境的变化发生分异,因而有较明显的地区差异.本研究依据吴征镒的种子植物分布类型的划分标准[16],逐一确定并统计其分布区类型.
2.2 数据处理方法
各区系成分所占比率(F)的计算公式[17]:
F=(Fi/T)×100%.
式中:Fi为某区系第i个区系成分的分类群数(如属或种)(i=1,2,3,…,n);T为某区系各种地理成分的分类群总数.即
聚类模型以属的地理成分为指标[18],求出分布区类型间距离矩阵,设有n个m维样本数据xij:i=1,2,…,n;j=1,2,…,m.计算各区系样本间标准欧氏距离(dik):
把每个分布区类型作为一类,将距离最近的两类归并为一个新类,采用系统聚类中组间均联法作图,谱系图通过SPSS18.0软件获得;以主坐标法(PCO)对各分布区类型间相似关系进行的排序分析,以及侧重于揭示相邻最小生成树(MST)的排序分析均通过PAST3.0统计软件获得.
3 结果与分析
3.1 区系组成
仙人洞保护区共分布有野生种子植物96科362属764种,其中裸子植物5科、12属、18种,双子叶植物81科、287属、629种,单子叶植物10科、63属、117种.可见,仙人洞植物物种数量较为丰富,其中被子植物居绝对优势,占本区种子植物总种数的97.64%;双子叶植物629种,单子叶植物117种,分别占总种数的82.32%和15.31%;而裸子植物比较匮乏,仅占总种数的2.36%.
3.2 属的分布区类型
以属为地理成分分析的基本单位,仙人洞种子植物可划归15个分布区类型(见表1).从表1可见,热带、亚热带分布属共有75属,占总属数的23.75%,其中泛热带分布属居首位,共有44属,无严格分布于热带的属,都是由热带向亚热带扩展并进一步向温带延伸到达本区的属;其次是热带亚洲至热带非洲分布属,共9属;旧世界热带分布属,本区有8属,多为草本植物和灌木,如香茶菜属(Rabdosia)、天门冬属(Asparagus)、八角枫属(Alangium)等;热带亚洲成分共有7属,有山胡椒属(Lindera)、葛属(Pueraria)和蛇莓属(Duchesnea)等.温带性质分布属共有236属,占总属数的74.67%,其中北温带成分居首位,共有116属,其绝对数量和相对数量均居该区系的第一位,许多是灌木和草本种类;木本属也比较丰富,是该地区植被的优势种和表征种,如赤松、千金榆(Carpinuscordata)、黑桦(Betuladahurica)、核桃楸;东亚分布属共有30属,其中典型的东亚成分有17属,如猕猴桃属(Actinidia)、溲疏属(Deutzia)、五加属(Acanthopanax)、紫苏属(Perilla)等.该分布型的变型中,中国-日本变型占有绝大多数,表明该区系与华东、日本植物区系的关系更密切;东亚-北美间断分布属有28属,如皂荚(Gleditsiajaponica)、五味子(Schisandrachinensis)、大丁草(Gerberaanandria)等,该分布类型中有部分古老属或原始科的代表,也有很多是少型或二型属.中国特有成分较为贫乏,仅有5属,都为残遗的古特有属,这与其远离中国特有属分布中心的地理位置是一致的.从以上分析可以看出,仙人洞种子植物区系以温带性质属为主,热带性质属也占有一定比例,应置于中国-日本森林植物亚区从暖温带向温带过渡的范围内.
3.3 不同山地种子植物属的分布区类型相似性
地理成分组成不仅是植物区系的一个重要特征,也是比较不同植物区系间关系的一个重要指标.植物分布区类型谱可以反映不同地理成分在该区系中占有的比率或对其区系总体的贡献[10],也可进一步揭示各区系间的相似关系,并消除调查面积对比较结果的影响.表1列出了仙人洞等16个山地种子植物属的分布区类型谱[12-18],在此基础上计算出欧式距离矩阵(见表2),然后进一步进行聚类和排序及相似性分析.

表1 16个山地种子植物属的分布区类型谱 %
注:F2—F15分别代表泛热带分布,热带亚洲及北美洲间断分布,旧世界热带分布,热带亚洲及热带澳大利亚分布,热带亚洲及热带非洲分布,热带亚洲分布,北温带分布,东亚及北美洲间断分布,旧世界温带分布,温带亚洲分布,地中海、西亚至中亚分布,中亚分布,东亚分布,中国特有分布.世界分布属不计,下同.

表2 16个山地种子植物属的分布区类型成分的欧式距离矩阵
3.3.1 聚类分析
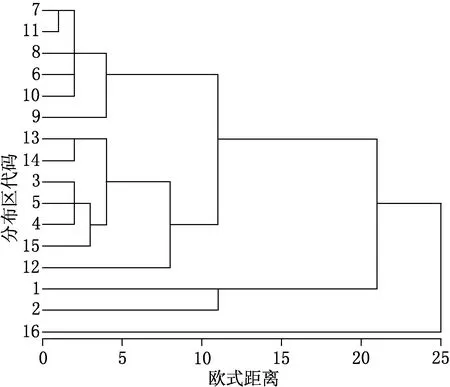
图1 16个山地种子植物属的分布区类型成分的聚类谱系
基于欧式距离矩阵,应用系统聚类中的组间均联法得到表征16个山地种子植物属的分布区类型关系的聚类谱系(见图1).图1可以直观地反映不同山地植物属分布区类型之间的相似性关系,在其相似性约为12的水平上,分为三类:第一类包括白石砬子和长白山;第二类包括凤凰山、千山、仙人洞、太行山、五台山、小五台山和东灵山,它们先聚合一起,再与徂徕山、蒙山、昆嵛山、泰山、崂山及艾山聚为一类;第三类为庐山单独.
3.3.2 排序分析
应用排序分析中的主坐标法得到表征16个山地种子植物属分布区类型关系的排序散点图和最小生成树(见图2).主坐标分析中坐标轴的特征根和方差贡献率是选择主坐标的依据.通过计算,第一主坐标的方差贡献率为82.141%,第二主坐标的方差贡献率为10.329%,前两个主坐标的累积方差贡献率为92.47%(见表3),表示前两个主坐标可表达全部信息的92.47%,意味着图2中只有7.53%的信息丢失.所以,选择前两个主坐标作为排序因子进行排序,基本显示了不同山体植物区系的相似性关系,大体可以把16个山地种子植物属的分布区类型划分为3组(见图2A),这和聚类分组情况大致匹配.主坐标结合最小生成树分析则清楚表明了不同区系的相似性及表达过程(见图2B).
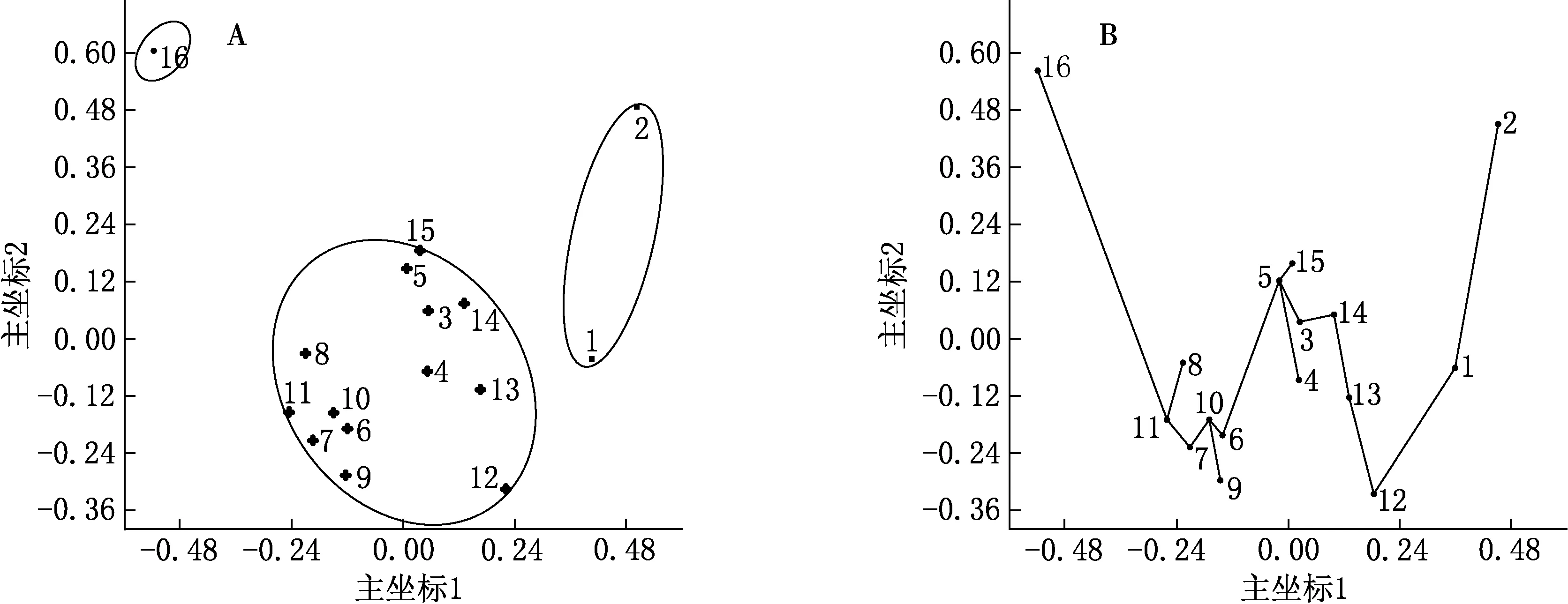
图2 16个山地种子植物属分布区类型成分的主坐标排序(A)及其最小生成树(B)
从排序散点图 (见图2A) 可见,沿着第一坐标轴,区系的纬度有从低到高递增的趋势,显示出第一坐标轴与纬度有关.随着纬度升高,区系的宏观气候条件发生规律性变化,区系内寒温带和温带性质的属逐渐增加,而亚热带和热带性质的属逐渐减少,表现出由亚热带向暖温带、温带逐渐过渡的趋势.沿着第二坐标轴,区系随经度也呈现一定的变化趋势,但相关性不明显,较为复杂,这可能与地形条件的影响有关.可见,纬度是影响植物区系分布组成的主要因子,而经度、地形条件对区系组成也有一定的影响.

表3 区系谱前两个主坐标的方差统计率和累计方差贡献率
3.3.3 相似性分析
基于各山地属分布区类型间的关系,通过聚类分析、排序分析,结合最小生成树分析,仙人洞等16个山地植物区系大体上可划分为3组:
(1) 长白山组.包括白石砬子和长白山,欧式距离为10.965,是中国-日本森林植物亚区的东北植物区系的主要组成成分,是东北地区生物多样性的中心,区系以温带区系成分为主.
(2) 千山泰山组.包括辽东半岛亚地区的千山、凤凰山和仙人洞,欧式距离为3.382~7.570,属暖温带湿润季风气候区;华北平原山地亚地区的五台山、小五台山和东灵山,属暖温带大陆季风性气候区,四季分明,冷热干湿界限明显,区系以温带区系成分为主;还有山东半岛亚地区的泰山、徂徕山、昆嵛山、崂山和艾山,欧式距离为3.011~6.362,属暖温带湿润季风气候区.
(3) 庐山组.以庐山为代表,地处亚热带东部湿润季风区,气候温和,四季分明,属热带和暖温带过渡类型,植物以亚热带区系成分为主,隶属于华东植物区系,具有由暖温带落叶阔叶林向亚热带常绿阔叶林的过渡特征.
4 结论与讨论
中国东北部地区是东亚地区植物多样性较丰富的地区,是探索东亚与中国-日本森林植物区系起源与分化的关键区域之一[19].辽宁仙人洞保护区地处东北地区南部,植物种类较为丰富,共分布有野生种子植物764种,隶属于96科362属,这与该区所处地理位置、地形及气候、降水等有密切的关系.从属的地理分布型看,仙人洞种子植物地理成分复杂,温带性质成分最多,占总属数的74.67%;热带性质成分次之,占总属数的23.75%.温带成分中有较多的属分布到亚热带和热带,无局限于热带地区分布的典型热带属,大多为热带地理成分中能够向北延伸分布的类群[20];植物以温带成分为主体,但兼有热带植物成分,其区系组成具有交汇过渡性的特点,是在旧的热带植物区系的基础上演变而成的暖温带山地植物区系,应划归中国-日本森林植物亚区从暖温带向温带过渡范围内的华北平原山地亚地区[21].
研究大尺度植物地理分布格局的重要方法是根据植物的自然属性,特别是分布的亲缘关系,把植物类群分成多种地理分布型[22].通过构建聚类模型和主坐标分析,仙人洞与其他不同山地种子植物区系的相似关系呈现出较为一致的组别划分,仙人洞与千山、凤凰山共同划归华北平原山地亚地区,支持了前人的结果[23].区系分组大体上是以地域为特征,其中纬度是影响区系类别和相似关系的重要因素之一.植物的分布区域和环境条件(如:能量、水资源可用性及生境异质性等)密切相关.不同山地植物分布类型,随着环境及水热条件的纬向变化也体现出一定的规律性.
本研究采用多种方法探究了多个山地植物区系的相似性.其中,聚类分析能够量化体现不同山地植物区系间的差异;主坐标分析则是一个从多变量中选出新的综合变量来反映原来多个变量主要信息的多元统计分析方法,用较少的主坐标对矩阵中的群体进行排序,从整体上较准确地了解各变量的综合贡献和各对象之间的关系;而最小生成树则清晰地表明了相邻区系的相似性.多种分析方法综合起来更能较准确地刻画山地区系间的相似性,可以相互补充、相互验证,充分反映出大区域尺度上不同山地植物区系间的相似性.
[1] PETERSON A T,LIEBERMAN B S. Species’ geographic distributions through time:playing catch-up with changing climates[J]. Evolution:Education and Outreach,2012,5(4):569-581.
[2] KIERG,MUTKE J,DINERSTEIN E,et al. Global patterns of plant diversity and floristic knowledge[J]. Journal of Biogeography,2005,32(7):1107-1116.
[3] ZHU H,MA Y X,YAN L C,et al. The relationship between geography and climate in the generic-level patterns of Chinese seed plants[J]. Acta Phytotaxonomica Sinica,2007,45(2):134-166.
[4] CULMSEE H,PITOPANG R,MANGOPO H. et al. Tree diversity and phytogeographical patterns of tropical high mountain rain forests in Central Sulawesi,Indonesia[J]. Biodiversity and Conservation,2011,20:1103-1123.
[5] OLMSTEAD R G. Phylogeny and biogeography in Solanaceae,Verbenaceae and Bignoniaceae:a comparison of continental and intercontinental diversification patterns[J]. Botanical Journal of the Linnean Society,2013,171(1):80-102.
[6] 张晓明,王月婷,唐丽丽,等. 辽宁仙人洞国家级自然保护区药用维管束植物多样性研究[J]. 吉林师范大学学报:自然科学版,2013(1):20-27.
[7] 张恒庆,高嵩,靖晶,等. 庄河仙人洞红松人工林遗传多样性的ISSR分析[J].辽宁师范大学学报:自然科学版,2009,32(3):348-350.
[8] 刘承江. 辽宁仙人洞国家级自然保护区森林生态系统服务功能价值评估[D].大连:辽宁师范大学,2009.
[9] 闫双喜,杨秋生,王鹏飞,等.中国部分地区种子植物区系亲缘关系的研究[J].武汉植物学研究,2004,22(3):226-230.
[10] 张宪强,张治国,张淑萍,等. 山东昆嵛山植物区系初步研究[J]. 植物研究,2003,23(4):492-499.
[11] 许崇梅,张颖颖,樊守金,等. 山东艾山植物区系研究[J]. 西北植物学报,2004,24(10):2083-2088.
[12] 茹文明,张峰. 山西五台山种子植物区系分析[J]. 木本植物研究,2000,20(1):36-47.
[13] 王士泉,贾泽峰,李法曾. 山东崂山植物区系研究[J]. 植物科学学报,2001,19(6):467-474.
[14] 赵遵田,王锡华,李京东,等. 山东省蒙山种子植物区系研究[J]. 山东科学,2005,18(4):42-51.
[15] 王振杰. 河北山地高等植物区系研究[D]. 石家庄:河北师范大学,2006.
[16] 吴征镒,孙航,周浙昆,等. 中国种子植物区系地理[M]. 北京:科学出版社,2010:120-314.
[17] 马克平,高贤明,于顺利.东灵山地区植物区系的基本特征与若干山区植物区系的关系[J].植物研究,1995,15(4):501-515.
[18] 刘利,张梅,康峰峰. 辽宁白石砬子植物区系及与邻近区系的关系[J].西北植物学报,2013(3):607-612.
[19] 曹伟.长白植物区维管束植物多样性及分布格局[D]. 沈阳:中国科学院沈阳应用生态研究所,2004.
[20] 涂业苟,余能富,吴能良,等. 江西铜钹山自然保护区种子植物区系初步研究[J].江西农业大学学报,2012,34(4):754-761.
[21] 郝朝运,刘鹏. 浙江磐安蕨类植物地理成分及区系相似性分析方法的研究[J].地理科学,2005,25(3):379-384.
[22] QIAN H,SONG J S,KRESTOV P,et al. Large-scale phytogeographical patterns in East Asia in relation to latitudinal and climatic gradients[J]. Journal of Biogeography,2003,30(1):129-141.
[23] 鲁法军,杜晓艳,曲畅游,等. 再论山东半岛和辽东半岛植物区系在区划中的地位[J]. 植物生态学报,2005,29(3):516-518.
(责任编辑:方 林)
Floristic compositions of spermatophyte and the similarity with other mountain land in Xianren Cave Natural Reserve in Liaoning Province
LIU Li1,HUANG Guo-hui2,ZHANG Mei2,YANG Yun-fei2
(1.College of Agriculture,Eastern Liaoning University,Dandong 118003,China;2.Key Laboratory of Vegetation Ecology,Ministry of Education,Institute of Grassland Science,Northeast Normal University,Changchun 130024,China)
Understanding the distribution of taxon in mountains is an important aim in ecology. In this paper,the floristic relationships between Xianren Cave National Natural Reserve(NNR) and other adjacent flora were analysed. Based on Plot survey,floristic spectrum,cluster analysis and principal coordinates analysis(PCO),the elements of seed plants in Xianren Cave and the similarity of adjacent flora were investigated. The results showed that:it is abundant in wild species composition of Xianren Cave NNR,the types of distribution are various and complex. 764 species in 96 families and 362 genera were identified;angiosperm accounted for 97.64% of the total of seed plants,and dicotyledon accounted for 82.32%. A total of 236 genera that accounted for 74.67% of seed plants were grouped into 15 geographical elements. The types of geographical distributing composition are various and complex,temperate floral elements are dominant,which have something in common with tropical flora. The present dates indicate Xianren Cave should belong to North China flora and the region should belong to the East Asia zone representing the feature of transition from warm temperate to temperate area-type. The study showed the similarities were obvious between Xianren Cave and Fenghuang moutain,Qian moutain,Taihang moutain,Wutai moutain,Xiaowutai moutain and Dongling moutain of the 16 moutains in this study,and the latitude is an important determining factor in geographical distribution composition.
spermatophyte;floristic spectrum;cluster analysis;similarity;Liaotung peninsula
1000-1832(2014)04-0116-06
10.11672/dbsdzk2014-04-022
2014-01-08
辽宁省科技攻关项目(2011204001);辽东学院博士科研启动资助项目.
刘利(1970—),男,博士,副教授,主要从事生态学及植物资源研究;通讯作者:杨允菲(1956—),女,教授,博士研究生导师,主要从事植物种群生态学研究.
Q 948.5 [学科代码] 180·5165
A
猜你喜欢
野生动物学报(2024年4期)2024-11-28 00:00:00
金沙江文艺(2021年12期)2021-12-22 02:14:10
安徽农业科学(2021年14期)2021-09-03 09:55:59
景德镇陶瓷(2021年1期)2021-03-24 05:47:22
炎黄地理(2019年5期)2019-10-20 01:59:19
世界热带农业信息(2019年11期)2019-01-05 12:18:59
作文周刊·小学二年级版(2018年40期)2018-04-09 08:18:16
贵阳文史(2017年3期)2017-11-24 13:00:58
华东师范大学学报(自然科学版)(2017年1期)2017-02-27 13:41:16
安徽农业科学(2016年9期)2016-10-14 22:12:23
