
茅台酒厂生态功能区4种优势树种光合特征比较
2014-07-25 11:29张习敏张宇斌
西南林业大学学报 2014年5期
张习敏 申 刚 陈 玲 张宇斌 乙 引
(1.贵州师范大学生命科学学院,贵州 贵阳 550001;2.贵州省植物生理与发育调控重点实验室,贵州 贵阳 550001)
茅台酒厂生态功能区4种优势树种光合特征比较
张习敏1,2申 刚1,2陈 玲1,2张宇斌1,2乙 引1,2
(1.贵州师范大学生命科学学院,贵州 贵阳 550001;2.贵州省植物生理与发育调控重点实验室,贵州 贵阳 550001)
以茅台酒厂生态功能区4种优势树种为材料,通过光合作用系统仪测定夏、冬两季光合作用日变化,进而评价树种对环境的适应性。结果表明,各树种上午的净光合速率(Pn)高于下午,暗呼吸速率(Rd)和光能利用率(LUE)无明显变化规律;对Pn日均值而言,在夏季,青冈栎显著高于箬竹、复羽叶栾树和构树,而在冬季,复羽叶栾树显著高于青冈栎、箬竹和构树;从夏冬两季总的Pn来看,青冈栎最高(6.90 μmol/(m2·s)),构树最低(2.34 μmol/(m2·s));复羽叶栾树的总Rd显著高于青冈栎、箬竹和构树,而青冈栎的总LUE最低。根据几个光合生理参数指标,该地区几种优势树种的适应能力表现为青冈栎>复羽叶栾树>箬竹>构树。
光合作用;优势树种;生态系统;自然植被
贵州茅台酒为世界名酒,相关试验与研究表明,茅台酒之所以具有独特的品质,原因之一是茅台酒厂坐落于独特的地质生态环境之中[1]。随着工业化进程和城镇化建设步伐的不断增快,导致很多原有保持良好的生态环境遭到毁灭性的破坏。同时,部分物种由于自身生物学、生态学原因,其生长和繁衍力显著下降,进而影响该地区的生态系统多样性。
生态系统是一个组成复杂、相互影响的有机组合,植物在生态系统中扮演着十分重要的角色,具有维持生态系统的物种多样性、主导群落更新演替、加速群落植被恢复进程等多种功能[2]。在森林生态系统的植物群落中,通常把个体数量多、盖度大、生物量高、体积较大、生活能力较强,占有竞争优势的植物物种称为优势种。植物群落中的优势种,特别是演替顶极中的优势种,控制着植被的结构,能更好地体现一个地区现有生态系统的状态[3-4]。前期调查研究发现,茅台酒厂生态功能区物种丰富度和分布均匀程度较低,该区域的主要优势树种有箬竹(Indocalamustessellatus)、复羽叶栾树(Koelreuteriabipinnata)、构树(Broussonetiapapyrifera)、青冈栎(Cyclobalanopsisglauca)。了解该地区群落植被的优势物种对环境的适应性,对保护生物多样性具有重要的意义。
光是影响植物生长发育和生存最重要的环境因子之一,也被认为是植物群落演替过程中促进物种相互取代的主要因子之一,优势植物对环境因子变化的适应性,以及植被群落演替均可以通过其光合生理生态学特性(尤其是净光合速率、呼吸速率及光能利用率等参数)快速反映出来[5-7]。本研究对该地区4种优势树种夏、冬两季的净光合速率、呼吸速率及光能利用率日变化进行测定,比较他们的光合作用能力,从光合生理角度进一步了解该地区植物与环境的适应关系。
1 研究地概况
贵州茅台酒厂生态功能区位于贵州省北部赤水河畔,地处东经106°22′,北纬27°51′,海拔约423 m,三面环山。年均温度17.4 ℃,年较差大,1月均温6.9 ℃,7月均温27.9 ℃。夏季最高气温达39.9 ℃,炎热季节持续半年以上;冬季气候暖和,温差小,霜期短,年平均无霜期达326 d,最低气温为2.7 ℃。干热少雨,年雨量仅有800~1 000 mm,日照丰富,年日照可达 1 400 h。土壤为紫色土,土层较厚,一般在50 cm左右。土壤(0~20 cm)有机质含量为1.4%,细菌2.37×104个/g,放线菌6.52×103个/g,真菌3.43×103个/g,土壤容重0.91 g/cm3;土壤全氮、全磷、全钾、钙、镁含量分别为0.3g/kg、201 mg/kg、7 775 mg/kg、3 978 mg/kg、6 931 mg/kg,土壤pH 6.77。
2 研究方法
2.1 研究对象
以茅台酒厂生态功能区优势树种箬竹、复羽叶栾树、构树、青冈栎为研究对象。
2.2 光合日变化的测定
分别在2010年8月上旬(夏季)和12月中旬(冬季)晴朗的天气,用LI-6400便携式光合测定系统(LI-COR Bioscience Inc., Lincoln, NE, USA)对4种自然生长的树种进行光合日变化的测定。直接测定的指标有大气温度(Tair,℃)、光合有效辐射(PAR,μmol/(m2·s))及相对湿度(RH,%)、净光合速率(Pn,μmol/(m2·s));净光合速率测定结束后,将叶片放在暗夹下适应10 min,之后测定叶片的暗呼吸速率(Rd,μmol/(m2·s))。光能利用率(LUE, %)=Pn/PAR。每次测定重复3~4次,测定时间从8:00—18:00,每隔2 h测定1次,对每个树种随机选择3~4株,选择阳生和充分展开的距枝条顶端第5、6片成熟叶测定,每次测量重复3~4次。
2.3 统计分析
用SPSS 18.0 (Statsoft Inc., College Station, TX, USA) 进行SNK均值比较分析,用SigmaPlot 10.0作图。
3 结果与分析
3.1 夏冬两季环境因子比较
在研究样地,测定的光合有效辐射、大气温度、相对湿度的夏冬两季日平均值见表1。光合有效辐射和大气温度夏季显著高于冬季;而相对湿度则在2个季节相差不显著。

表1 茅台酒厂生态功能区光合生态因子夏冬两季日平均值
注:同列中不同的小写字母表示差异显著(P<0.05)。
3.2 生态功能区4个优势树种气体交换变化比较
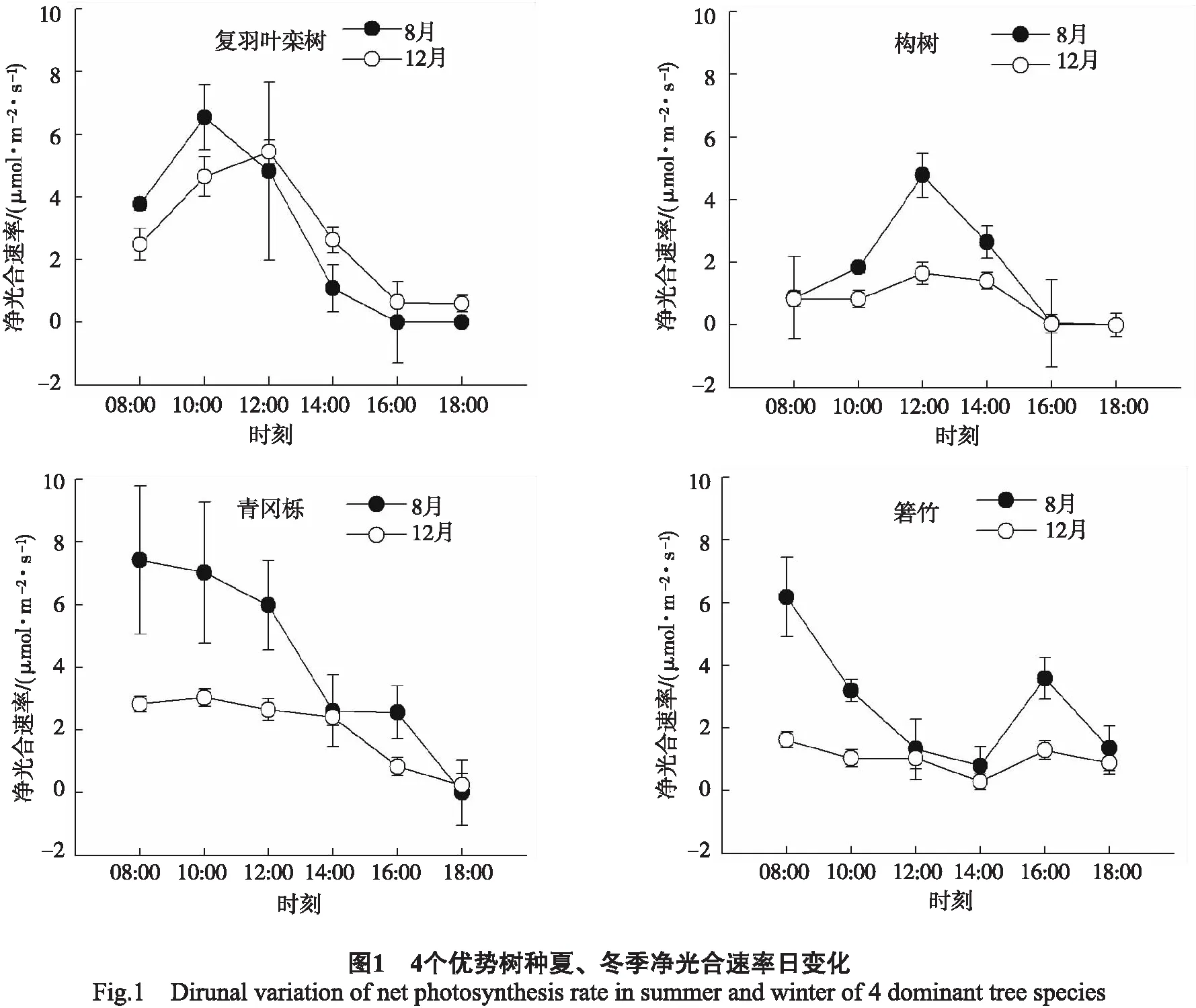
3.2.1净光合速率日变化 对一个具体树种而言,夏冬两季的Pn日变化趋势基本相同,而不同树种的Pn日变化不同,具体表现为:复羽叶栾树、构树的Pn呈单峰型的变化趋势,青冈栎为下降型变化趋势,而箬竹则为双峰型的变化。复羽叶栾树峰值出现在12:00前后,构树的峰值也出现在12:00, 而青冈栎的Pn峰值则出现在8:00左右,箬竹第1个峰值出现在8:00左右,第2个峰值出现在16:00左右(图1)。
在4个树种中,夏季各树种的Pn最大值和最小值为复羽叶栾树(6.54、0.003 μmol/(m2·s))、箬竹(6.10、0.800 μmol/(m2·s))、构树(4.78、0.001 μmol/(m2·s))、青冈栎(7.42、0.005 μmol/(m2·s)); 冬季各树种的Pn最大值和最小值为复羽叶栾树(5.44、0.590 μmol/(m2·s))、箬竹(1.62、0.290 μmol/(m2·s))、构树(1.65、0.001 μmol/(m2·s))、青冈栎(2.83、0.230 μmol/(m2·s))(图1)。对日平均值而言,夏季从大到小分别为青冈栎﹥箬竹﹥复羽叶栾树﹥构树; 冬季从大到小分别为复羽叶栾树﹥青冈栎﹥箬竹﹥构树(表2)。
3.2.2暗呼吸速率日变化 4个树种的Rd夏季均高于冬季(图2)。在夏季,Rd日变化的最大值从高到低依次为青冈栎(3.60 μmol/(m2·s))﹥复羽叶栾树(3.11 μmol/(m2·s))﹥构树(1.98 μmol/(m2·s))﹥箬竹(1.33 μmol/(m2·s));在冬季4个树种的Rd值差异不明显(图2)。夏季Rd日平均值从大到小的顺序依次为复羽叶栾树﹥构树﹥青冈栎﹥箬竹;而冬季Rd值从大到小的顺序依次为复羽叶栾树﹥青冈栎﹥箬竹﹥和构树(表2)。
3.2.3光能利用率日变化 对日变化而言,夏冬两季相比,复羽叶栾树上午的LUE夏季高于冬季,而下午则冬季高于夏季;青冈栎的LUE夏季均高于冬季,构树和箬竹的LUE则为无规则的变化(图3)。对日平均值而言,构树夏季的LUE显著高于复羽叶栾树、箬竹和青冈栎;而冬季则是箬竹和复羽叶栾树显著高于青冈栎和构树(P<0.05)(表2)。
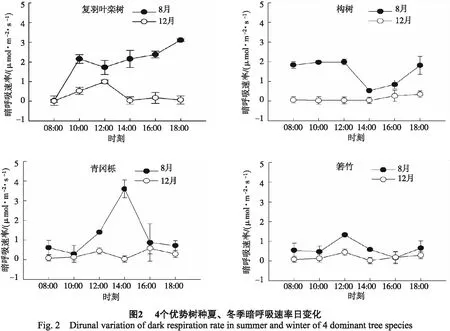
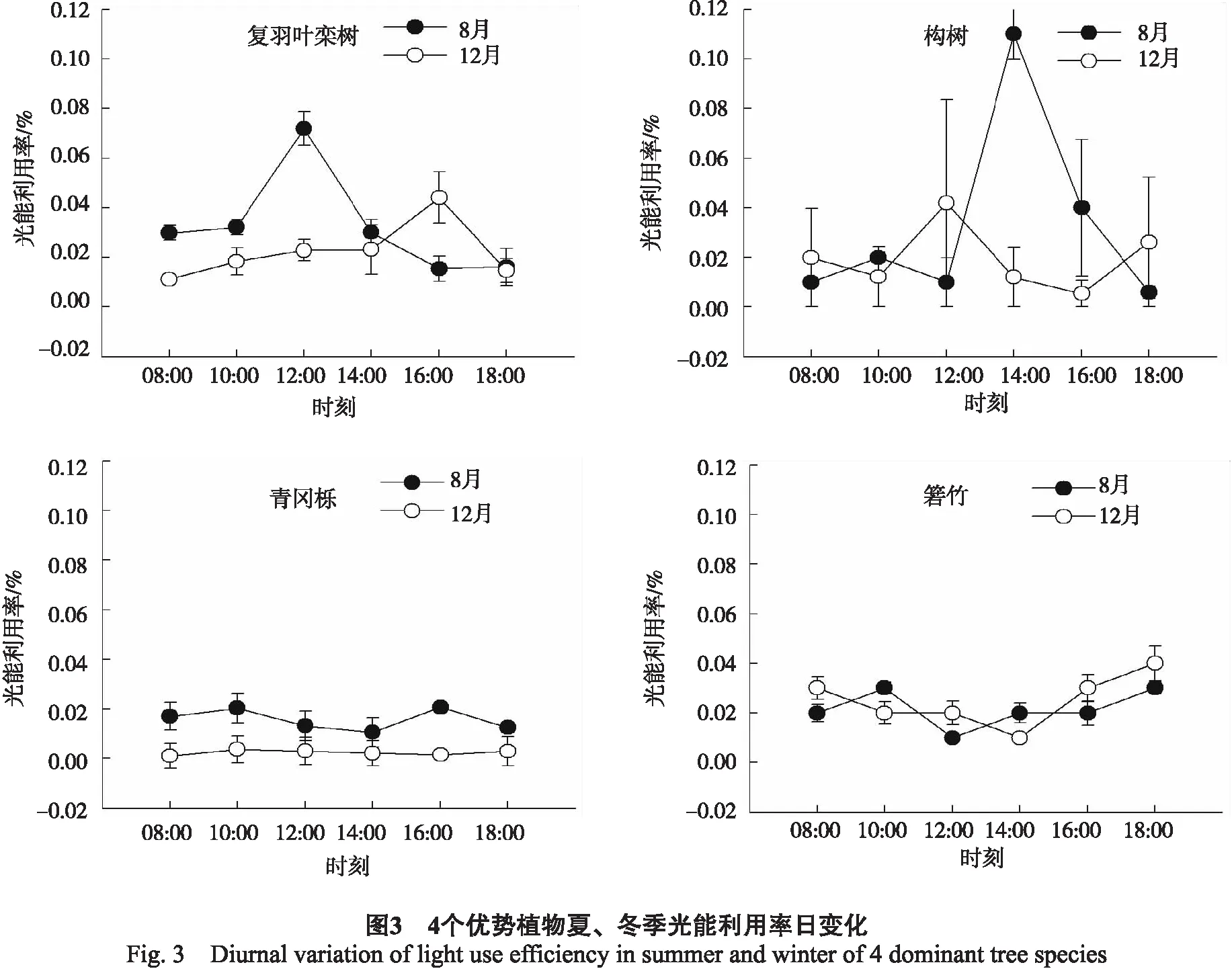
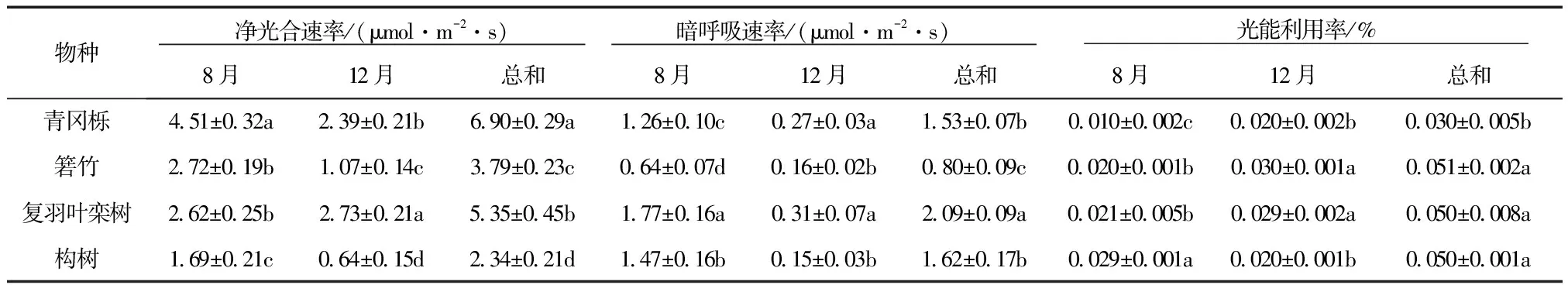
表2 4个树种光合特性指标的日平均值及两季总值
注:同列中不同小写字母表示不同树种之间差异显著(P<0.05)。
3.2.4光合代谢比较 将4个树种夏、冬两季Pn、Rd和LUE分别求和后,其物种间的差异较大(表2)。青冈栎的Pn(6.09 μmol/(m2·s))显著(P<0.05)高于其他3个树种;构树最低(2.34 μmol/(m2·s));复羽叶栾树的Rd(2.09 μmol/(m2·s))显著(P<0.05)高于其他3个树种,其中青冈栎与构树之间的差异不显著;对光能利用率而言,复羽叶栾树、构树和箬竹均显著(P<0.05)高于青冈栎。
4 结论与讨论
虽然被测定的4个树种位于茅台酒厂生态功能区内,但是由于分布的具体位置不同,从而表现出环境因子的微小变化(数据未列出),但总体趋势不变。植物光合作用的影响因子比较复杂,环境因子(如光合有效辐射、大气温度、相对湿度等)的日变化、季节变化均影响植物光合作用的日变化和季节差异[8-13]。在本研究中,光合有效辐射、大气温度夏季高于冬季,2个季节的相对湿度变化不明显(表1),表明夏冬两季光合作用主要受光照和温度等环境因子的影响。同时,光合作用也受到植物自身的内部因素(品种特性、叶龄、叶位、库源关系、叶绿素荧光特性等)影响[14]。
在4个树种中,上午的Pn几乎均高于下午,可能是上午的温度低于下午,下午的高温降低了参与光合代谢过程的相关酶活性;同时,高温也可能诱导植物叶片气孔关闭进而避免水分蒸腾,但同时也限制了环境CO2进入气孔,因而限制胞间CO2浓度及CO2同化速率。这与苏军德等[15]研究的祁连圆柏(Sabinaprzewalskii)和兰小中等[16]研究西藏巨柏(Cupressusgigantea)的结果一致。
4个树种的Pn、Rd和LUE日变化出现最大值的时间不同(图1,图3),这可能是它们在该区域的株高和冠幅不同。植株高、冠幅大的树种可以提前利用太阳光能,而植株矮小、冠幅小的树种需要在中午才能完全捕捉到充足的太阳光,因而达到最大光合速率的时间可能被推迟。箬竹的Pn在中午出现低谷,其后缓慢上升,这可能是箬竹在中午出现“午休”的现象。
净光合速率是反映植物光合能力最为重要的指标,光合速率越大,表明其光合生产能力越强。除复羽叶栾树外,青冈栎、箬竹、构树3个树种夏季的净光合速率日均值均高于冬季(表2),这可能是夏季光合有效辐射、温度等环境因子高于冬季(表1);同时,也可能是这些植物夏季生长旺盛,而冬季生长缓慢,光合代谢较慢。而复羽叶栾树夏、冬两季净光合速率日均值差异不明显,表明其在1年中受环境因子的影响较小。当然,环境因子和植物自身的因素对光合作用的影响大小还需要进一步研究。对于夏、冬两季总净光合速率日均值,青冈栎和复羽叶栾树较高,分别为6.90、5.35 μmol/(m2·s)(表2),表明它们具有较强的光合能力。暗呼吸速率反映在无光条件下消耗能量的大小[17],复羽叶栾树的总暗呼吸速率最大(表2),表明其消耗能量较大,即光合产物中大部分能量被呼吸消耗掉,因此其植株生长矮小,处于森林底层,这与本项目组的调查结论一致。在森林冠层底部,光合有效辐射较低,因此,植物叶片通过调整光能利用率来适应低光环境[18]。4个树种中,青冈栎植株最高,其光能利用率最低,箬竹、复羽叶栾树和构树的光能利用率较高(表2),表明其在林冠底层能够充分适应低光环境,特别是复羽叶栾树。综合分析,该地区优势树种的适应能力表现为:青冈栎>复羽叶栾树>箬竹>构树。
[1] 陈兴烯, 季克良. 茅台酒的独特性概述[J].酿酒科技, 2006, 140 (2):79-84.
[2] 魏作东, 杨大荣, 彭艳琼,等. 榕树在西双版纳热带雨林生态系统中的作用[J]. 生态学杂志, 2005, 24(3): 233-237.
[3] 胡楠, 范玉龙, 丁圣彦,等. 伏牛山自然保护区森林生态系统乔木植物功能型分类[J]. 植物生态学报, 2008, 32(5): 1104-1115.
[4] Box E O. Factors determining distributions of tree species and plant functional types[J]. Vegetatio, 1995, 121: 101-116.
[5] 吴统贵,吴明,萧江华.杭州湾滩涂湿地植被群落演替与物种多样性动态[J].生态学杂志, 2008, 27(8):1284-1289.
[6] 李薇,唐海萍. 准噶尔盆地荒漠区短命植物光合蒸腾特性及影响因素研究[J].西北植物学报, 2006, 26(12):2517-2522.
[7] Monteith J L. Solar Radiation and productivity in tropical ecosystems[J]. The Journal of Applied Ecology, 1972, 9(3):747-766.
[8] 张旺锋, 樊大勇, 谢宗强,等. 濒危植物银杉幼树对生长光强的季节性光合响应[J]. 生物多样性, 2005, 13 (5): 387-397.
[9] 施建敏, 杨光耀, 杨清培,等. 厚壁毛竹光合作用对环境因子响应的季节变化[J]. 林业科学研究, 2009, 22(6): 872-877.
[10] 李荣生,尹光天,杨锦冒,等. 蛇皮果幼株净光合速率初步研究[J].西南林业大学学报,2012,32(4):100-102.
[11] Chaerle L, Saibo N, Van D S. Tuning the pores: towards engineering plants for improved water use efficiency[J]. Trends in Biotechnology, 2005, 23:308-315.
[12] Israelsson M, Siegel R S, Young J, et al. Guard cell ABA and CO2signaling network updates and Ca2+sensor priming hypothesis[J]. Current Opinions in Plant Biology, 2006, 9: 654-663.
[13] El-sharkawy M A. International research on cassava photosynthesis, productivity, eco-physiology, and responses to environmental stresses in the tropics[J]. Photosynthetica, 2006, 44 (4): 481-512.
[14] 孙猛, 吕德国, 刘威生. 杏属植物光合作用研究进展[J]. 果树学报, 2009, 26(6): 878-885.
[15] 苏军德, 勾晓华, 曹宗英,等. 祁连圆柏光合作用日变化特征及其与生理生态因子的关系[J]. 西北植物学报, 2011, 31( 5) : 1011-1017.
[16] 兰小中, 廖志华, 王景升. 西藏高原濒危植物西藏巨柏光合作用日进程[J]. 生态学报, 2005, 12: 3172-3175.
[17] 王冉. 中国12种珍稀树种光合生理特性[J].东北林业大学学报, 2010, 38(11):15-20.
[18] 王军邦, 张秀娟, 韩海荣,等. 亚热带常绿人工针叶林冠层内光能利用率[J]. 生态学杂志, 2010, 29(4):611-616.
(责任编辑 张 坤)
Comparison of Photosynthetic Characteristics of 4 Dominant Tree Species in Maotai Liquor Production Area
ZHANG Xi-min1,2, SHEN Gang1,2, CHEN Ling1,2, ZHANG Yu-bin1,2, YI Yin1,2
(1.College of Life Sciences,Guizhou Normal University,Guiyang Guizhou 550001,China; 2 Kay Laboratory of Plant Physiology and Development Regulation, Guiyang Guizhou 550001,China)
4 dominant tree species in the Maotai liquor production area were analyzed their photosynthetic characteristics of diurnal variation in summer and winter by using a portable photosynthesis system LI-6400, and then evaluated the acclimation of these species to environment. The results showed that net photosynthesis rate (Pn) of tree species were higher in morning than those in afternoon, but dark respiration rate (Rd) and light use efficiency (LUE) had not evident changing-regular.Diurnal mean values ofPnwere higher inCyclobalanopsisglaucathan those inIndocalamustessellatus,KoelreuteriabipinnataandBroussonetiapapyriferain summer. In contrast, Diurnal mean value ofPnwere higher inK.bipinnatathan those inC.glauca,I.tessellatusandB.papyriferain winter. GrossPnin summer and winter was the highest forC.glauca(6.90 μmol/ (m2·s)) and was the lowest forB.papyrifera(5.35 μmol/ (m2·s)). GrossRdinK.bipinnatawas higher than those inC.glauca,I.tessellatus,B.papyrifera. Gross LUE was the lowest forC.glauca. According to the parameters of photosynthetic physiology, the acclimation of these species to environment wasC.glauca>K.bipinnata>I.tessellatus>B.papyrifera.
photosynthesis; dominant tree species; ecological system; natural vegetation
2014-05-04
长江学者和创新团队发展计划资助(PCSIRT-1227);贵州茅台科技联合基金(黔科合茅科联字[7006]号)资助;贵州省重点实验室建设项目(黔科合计Z字[2011]4005号)资助。
乙引(1967—),男,博士,教授。研究方向:植物生理生态。Email: yiyin@gznu.edu.cn。
10.3969/j.issn.2095-1914.2014.05.010
S718.45
:A
:2095-1914(2014)05-0053-06
第1作者:张习敏(1980—),男,硕士,讲师。研究方向:植物生理生态。Email:zhxm409@163.com。
猜你喜欢
世界竹藤通讯(2022年4期)2022-11-28
广东蚕业(2022年8期)2022-09-30
食品工业科技(2022年9期)2022-04-26
天天爱科学(2020年11期)2020-09-10
竹子学报(2019年4期)2019-09-30
作文成功之路·教育教学研究(2019年2期)2019-03-20
作文成功之路·小学版(2019年2期)2019-01-14
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
长江文艺(2017年2期)2017-02-13
