
镉处理下平邑甜茶DNA的甲基化分析
2014-07-07 15:38:55王利李鹏飞杨洪强谢会成李龙谢永波
山东农业大学学报(自然科学版) 2014年3期
王利,李鹏飞,杨洪强,谢会成,李龙,谢永波
1.山东农业大学农业生态与环境重点实验室,山东泰安271018 2.山东省东营市林业局,山东东营257091 3.山东农业大学农业资源利用博士后流动站,山东泰安271018
镉处理下平邑甜茶DNA的甲基化分析
王利1,3,李鹏飞2,杨洪强3*,谢会成1,李龙1,谢永波1
1.山东农业大学农业生态与环境重点实验室,山东泰安271018 2.山东省东营市林业局,山东东营257091 3.山东农业大学农业资源利用博士后流动站,山东泰安271018
采用甲基化敏感扩增多态性(Methylation sensitive amplified polymorphism,MSAP)技术,对镉处理的平邑甜茶叶片DNA进行甲基化位点分析,研究镉对其甲基化多态性的影响。结果表明:在0(CK)、2.5 mg·L-1、5 mg·L-1、10 mg·L-1镉处理下,9对MSAP引物扩增,甲基化出现三种不同模式即未甲基化A、半甲基化B和全甲基化C,共扩增出3150条谱带,单对引物最多扩增656条,最少为610条。5 mg.L-1镉处理的平邑甜茶甲基化较高,比例达到60.1%;10 mg.L-1镉处理的平邑甜茶甲基化较低,比例达到57.6%;与对照相比,镉处理的平邑甜茶甲基化水平降低,可能镉抑制了其基因的表达。
平邑甜茶;甲基化;甲基化敏感扩增多态性
DNA甲基化是指生物体在DNA甲基转移酶(DNA methyltransferase)的催化下,将甲基供体S-腺苷甲硫氨酸(SAM)转移到特定碱基上的过程。在动物和植物中普遍存在DNA甲基化,它与植物的生长发育、进化有密切的关系。植物DNA甲基化水平受发育时期[1-2]、不同发育组织[3-4]、不同种质资源[5-6]、DNA损伤与修复[7]及不同胁迫等的影响[8]。不同胁迫对植物甲基化影响存在差异,低温使水稻甲基化差异片段CIDM 7的去甲基化表达增强[9],盐胁迫使小麦叶片发生超甲基化[10],水分胁迫导致水稻叶片根DNA甲基化平均水平明显增加,其中根部增加幅度尤为明显[11],生物胁迫使小麦甲基化增加[12],5~100 mg/L铬处理小麦幼苗的研究表明100 mg/L铬浓度导致3 d龄小麦幼苗根系DNA胞嘧啶甲基化水平降低,其他处理浓度却增加。重金属镉引起拟南芥、萝卜甲基化程度的提高[13-14],但是,关于镉究竟如何影响平邑甜茶甲基化还未见阐述,为此,以平邑甜茶为材料,利用镉对其处理,探明镉处理下平邑甜茶DNA甲基化的变化。
1 材料与方法
1.1 材料与处理
取大小一致的平邑甜茶种子,经消毒层积1个多月后播在育苗容器中。当幼苗第2片真叶刚出现时,选生长相近的植株移至1/2 Hoagland营养液中,每3 d换1次培养液,培养至出现第6片叶后,在营养液中加入氯化镉,使其终浓度为0(CK)、0.5 mg·L-1、2.5 mg·L-1、5 mg·L-1、10 mg·L-1,每处理3株,重复3次,30 d后,提取叶片DNA,利用甲基化敏感扩增多态性(Methylation sensitive amplified polymorphism,MSAP)检测叶片DNA甲基化的多态性。
1.2 引物

1.3 引物对的筛选
从5个供试样品中选出2个具有代表性的样品利用银染技术对63对引物进行筛选,筛选出E-AAC/H-TCG、E-AAC/H-TGT、E-AAC/H-TCC、E-AAG/H-TCG、E-AAG/H-TGC、E-AAG/H-TCC、E-ACA/H-TCG、E-ACA/H-TGT、E-ACA/H-TCC9对引物对供试样品进行扩增。从中筛选出检测位点较多,分布均匀,清晰可辨且多态位点百分比较高的引物对各个样品进行MSAP分析。
1.4MSAP分析
据北京鼎国生物技术有限责任公司FISH-MSAP试剂盒说明,先进行EcoRI/HpaⅡ、MspⅠ酶切与T4连接酶连接,取2 μL酶切连接液做模板,用带有1个选择性碱基的引物进行预扩增,再将预扩增液按1:15比例稀释,以2 μL预扩增稀释液为模板,用带有3个选择性碱基的引物对进行选择性扩增。变性产物(含有内标GeneScan-500)上样于4%变性聚炳烯酰胺凝胶,恒定功率50 W、最大电压3000 V,进行电泳2.4 h,利用ABI 377测序仪对其进行检测。
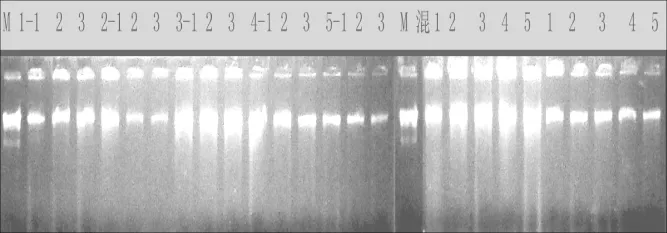
图1 DNA琼脂糖电泳结果Fig.1 Result of DNAagarose electrophoresis
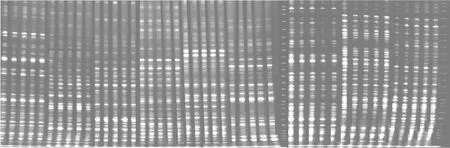
图2 EcoR I/Hpa II酶切扩增结果Fig.2 Results of EcoR I/Hpa II enzyme amplification

图3 EcoR I/Msp I酶切扩增结果Fig.3 Results of EcoR I/Msp I enzyme amplification
1.5 统计分析
利用GeneScan3.1软件对图像进行处理,构建0、1数学矩阵。通过比较电泳谱带的差异,可以推测出CCGG的甲基化情况[15]。
2 结果与分析
2.1 基因组的质量
本实验提取平邑甜茶DNA经琼脂糖电泳检测(如图1),主带清晰,片段大小在23 kb左右,无降解。这说明该法提取的银杏DNA质量较高,符合MSAP技术要求。
2.2 镉处理下平邑甜茶DNA甲基化模式分析
平邑甜茶DNA经过EcoR I/Hpa II、EcoR I/Msp I酶切、连接、预选扩增、聚丙烯酰胺凝胶电泳后得到MSAP图谱(图2,图3),谱带清晰。图中谱带表明其甲基化主要有三种不同模式:模式A是指EH、EM均有带,代表非甲基化位点,说明未发生甲基化(或单链酶切位点内侧甲基化);模式B是指EH有带、EM无带,代表半甲基化位点,说明该位点发生了单链酶切位点外侧的胞嘧啶甲基化;模式C是指EH无带、EM有带,代表全甲基化位点,说明该位点发生了双链酶切位点内侧的胞嘧啶甲基化。
2.3 镉处理下平邑甜茶DNA甲基化水平分析
MSAP图谱中每个泳道的每一条带代表一个酶切位点,根据有带记为1,无带记为0的方法分析统计其电泳谱带。9对引物共扩增出3150条谱带,单对引物最多扩增656条,最少为610条。在镉处理中,5 mg.L-1镉处理的平邑甜茶甲基化条带最多,共385条,甲基化达到60.1%;2.5 mg.L-1镉处理的平邑甜茶甲基化条带最少,共355条,甲基化达到57.8%。在扩增位点中,检测到甲基化位点1871个,甲基化比例为59.4%;其中,半甲基化位点941个,比例为29.9%,全甲基化位点为930个,比例为29.5%,半甲基化与全甲基化位点比例接近,说明镉处理对单链酶切位点外侧的胞嘧啶甲基化和双链酶切位点内侧的胞嘧啶甲基化的影响差异不大。另外,模式B中对照扩增谱带最多206条,10 mg.L-1镉处理最少167条,总体呈现减少的趋势;模式C中对照扩增谱带最多197条,2.5 mg.L-1最少168条,出现先减少后增加的变化。与对照相比,镉处理使平邑甜茶甲基化位点数量总体出现减少的趋势(表1)。
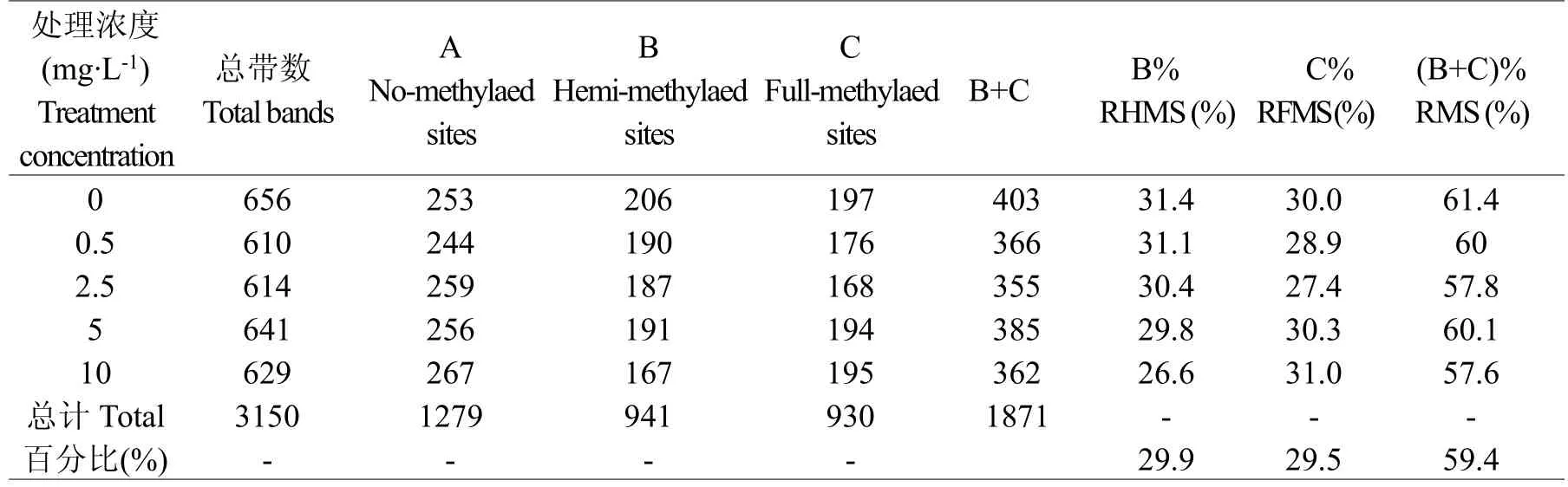
表1 镉处理下平邑甜茶叶片甲基化多态性分析Table1Analysis of methylation polymorphism on Malus hupehensis leaves under Cd treatment
3 讨论
表观遗传学是指在DNA序列没有发生改变的情况下,遗传信息在基因功能上发生可遗传的变化,导致表型变异[16],近10年来逐渐受到广泛重视,其研究成为当今生命科学研究的热点领域。植物DNA甲基化是在甲基转移酶的作用下,把DNA的一个甲基添加在DNA分子的碱基上,通常加在胞嘧啶上,形成甲基胞嘧啶[16]。在植物正常发育过程中,DNA甲基化能够调控生长发育等过程,对植物生长发育具有重要的调节作用。DNA甲基化的水平模式发生改变,就会影响到植物的生长发育,使其生长发育异常。李海林等研究巴西橡胶树DNA甲基化模式表明,其存在非甲基化、半甲基化和全甲基化三种模式[17],本实验研究也得到了上述三种不同的甲基化模式。不同植物甲基化多态性水平不一[14,17]。随着镉处理浓度的增加,拟南芥甲基化程度增加[14]。本研究结果表明,镉处理影响平邑甜茶叶片DNA的甲基化水平,使半甲基化和甲基化多态性发生了改变,它们的多态性比例为26.6~31.1%,尤其是使平邑甜茶叶片DNA半甲基化多态性出现减小的趋势,这与马子成等研究结果相反,甲基化多态性出现减小可能是镉处理抑制了基因的表达。
4 结论
(1)在镉处理中,甲基化出现三种不同模式即未甲基化A(EcoR I/Hpa II、EcoR I/Msp I酶切扩增均有带)、半甲基化B(EcoR I/Hpa II酶切扩增有带、EcoR I/Msp I酶切扩增无带)
(2)在镉处理中,5 mg.L-1镉处理的平邑甜茶甲基化较高,比例达到60.1%;10 mg.L-1镉处理的平邑甜茶甲基化较低,比例达到57.6%;与对照相比,镉处理的平邑甜茶甲基化水平降低,可能镉抑制了其基因的表达和全甲基化C(EcoR I/Hpa II酶切扩增无带、EcoR I/Msp I酶切扩增有带)。
[1]Finnegan EJ,Genger RK,Peacock W J.et al.DNA methylation in plant.Ann.Rev[J].Plant Mol.Biol.,1998,49:233-247
[2]Finnegan EJ,Peacock WJ,Dennis ES.DNA methylation,a key regulator of plant development and other proc esses[J].Current Opinion in Genetics and Development,2000,10:217-223
[3]Lu CM,J iang GM,Wang B S,et al.Photosystem II photochemistry and photosynthetic pigment composition in salt-adapted halophyte Artimisia anethfolia grown under outdoor conditions[J].J Plant Physiol,2003,160:403-408
[4]仪治本,孙毅,牛天堂,等.高梁基因组DNA胞嘧啶甲基化在杂交种和亲本间差异研究[J].作物学报,2005,31(9):1138-1143
[5]Guo M,Davis D,Birchler J A.Dosage effects on gene expression in a maize ploidy series[J].Genetics, 1996,142:1349-1355
[6]Keyte AL,Percifield R,Liu B,et al.Infraspecific DNA methylation polymorphism in cotton(Gossypium hirsutum L.). J Heredity,2006,97(5):444-450
[7]武立鹏,朱卫国.DNA甲基化的生物学应用急检测方法进展[J].中华检验医学杂志,2004,27(7):468-474
[8]范建成,刘宝,王隽媛,等.萘胁迫对水稻基因组DNA甲基化模式及水平的影响[J].环境科学,2010,31(3):793-800
[9]华扬,陈学峰,熊建华,等.水稻冷胁迫诱导的甲基化差异片段CIDM7的分离和分析[J].遗传,2005,27(4):595-600
[10]钟兰,王建波.DNA超甲基化在小麦耐盐胁迫中的作用[J].武汉植物学研究,2007,25(1):102-104
[11]潘雅娇,傅彬英,王迪,等.水稻干旱胁迫诱导DNA甲基化时空变化特征分析[J].中国农业科学,2009,42(9):3009-3018
[12]付胜杰,王晖,冯丽娜,等.叶锈菌胁迫下的小麦基因组MSAP分析[J].遗传,2009,31(3):297-304
[13]杨金兰,柳李旺,龚义勤,等.镉胁迫下萝卜基因组DNA甲基化敏感扩增多态性分析[J].植物生理与分子生物学学报,2007,33(3):219-226
[14]王子成,马洪霞,何艳霞.重金属镉对拟南芥DNA甲基化的影响[J].植物生理学通讯,2009,45(2):115-118
[15]陆光远,伍晓明,陈碧云,等.油菜种子萌发过程中DNA甲基化的MSAP分析[J].科学通报,2005,50(24):2750-2756
[16]薛京伦,旺旭,吴超群,等.表观遗传学-原理、技术与实践[M].上海:上海科学技术出版社,2006
[17]李海林,吴春太,李卫国.巴西橡胶树DNA甲基化的MSAP分析[J].分子植物育种,2011,9(1):89-73
Analysis of Methylation Polymorphism on Malus hupehensis Leaves under Cd Treatment
WANG Li1,3,LI Peng-fei2,YANG Hong-qiang3*,XIE Hui-cheng1,LI Long1, XIE Yong-bo1
1.Key Laboratory of Agricultural Ecology and Environment,Shandong Agricultural University,Taian 271018,China 2.Forestry Station,Dongying Municipal Forestry Bureau,Dongying 257091,China 3.Post-doctoral Mobile Station of Agricultural Resource Utilization,Shandong Agricultural University,Taian 271018,China
Methylation site and methylation polymorphism were analyzed on DNA of Malus hupehensis Rehd.under Cd treatment with methylation sensitive amplified polymorphism(MSAP).The results showed that amplification of 9 primer combinations formed three different modes and produced 3150 bands.The maximum number of bands was 656 and the minum number of bands was 610 per primer combination under different concengtration Cd treatment of 0(CK),2.5 mg·L-1, 5 mg·L-1,10 mg·L-1.The highest rate of methylation polymorphism appeared under 5 mg·L-1Cd treatment with value of 60.1%,the lowest rate of methylation polymorphism appeared under 10 mg·L-1Cd treatment with value of 57.6%.The level of methylation polymorphism on Malus hupehensis Rehd under Cd treatment reduced compared with control sample.Its reason might be Cd incurred the expression of gene.
Malus hupehensis Rehd.;methylation;methylation sensitive amplified polymorphism
F812.4
A
1000-2324(2014)03-0372-05
2013-03-06
2013-03-11
山东农业大学博士后课题,苹果优质高效生产关键技术研究与示范(2014BAD16B02)
王利(1968-),男,山东高密人,博士,研究方向为林木遗传与生理.
*通讯作者:Author for correspondence.E–mail:labft@sdau.edu.cn
猜你喜欢
家庭影院技术(2021年7期)2021-08-14 02:58:28
现代仪器与医疗(2021年2期)2021-07-21 02:19:16
吉林大学学报(理学版)(2021年3期)2021-05-26 02:24:00
中国当代医药(2020年26期)2020-11-06 07:22:10
生物信息学(2020年2期)2020-07-09 01:27:56
中国果业信息(2019年10期)2019-01-05 07:42:30
农产品市场周刊(2016年20期)2016-06-22 16:35:16
分析化学(2015年10期)2015-11-03 07:38:01
百科知识(2015年13期)2015-09-10 07:22:44
质谱学报(2015年5期)2015-03-01 03:18:25
