维生素的药物基因组学研究进展
2014-05-17 02:27张月丽周宏灏
中国药理学通报 2014年11期
张月丽,周宏灏,张 伟
(1.中南大学湘雅医院临床药理研究所,湖南长沙 410008;2.中南大学临床药理研究所,湖南省遗传药理学重点实验室,湖南长沙 410078)
药物基因组学(pharmacogenomics,PGx)广泛研究DNA和RNA的变化特点及与之相关的药物反应,有助于更好地理解药物疗效的个体化差异和药物的安全性。例如6-巯基嘌呤和硫唑嘌呤,与巯基嘌呤甲基转移酶 (TPMT)之间的关系;华法林应用过程中的CYP2C9和VKORC1药物基因组学研究[1-2]和阿巴卡韦与携带HLA-B*5701患者使用后的毒性问题[3],这些例子都涉及高危人群的安全性和药物有效性剂量的调整。药物基因组学研究的目的是提高药物的有效性和安全性,尤其是在癌症治疗的领域。基因检测的应用提高了人们研究心血管疾病、癫痫和HIV的兴趣[4]。在这些疾病中药物的药代动力学(PK)和药效动力学(PD)受代谢酶、膜转运蛋白和受体的遗传差异的影响。即使当药物基因组学研究评估的结果都是中立时(没有发现基因组效应),这些信息也可以通过确认某些可疑路径对PK、PD、疗效或安全的个体化差异的变化指导药物的应用。然而,药物基因组学对营养成分包括碳水化合物、蛋白质、脂肪、维生素和矿物质的研究非常少,在本文中,我们主要总结近年来有关维生素的药物基因组学研究进展。
维生素(vitamin),是维持人体生命活动必需的一类有机物质,也是保持人体健康的重要活性物质。虽然,维生素每日的需求量很少,但却是机体维持生命所需,是一种有着不同的化学结构和生理功能的多种不同类型的低相对分子质量的有机化合物。各种维生素的化学结构以及性质不同,却具备一些共同点:维生素多以前体的形式存在于食物中;维生素不是构成机体组织和细胞的组成成分,也不会产生能量,它的主要作用是参与机体代谢的调节;大多数的维生素,机体不能合成或合成量不足,必须经常通过食物获得;人体对维生素的需求量很小,日需要量常以毫克 (mg)或微克(μg)计算[5],但一旦缺乏就会引发相应的维生素缺乏症,如缺乏维生素A会出现夜盲症、干眼病和皮肤干燥;缺乏维生素D可患佝偻病等,对人体健康造成损害。
维生素是人体代谢中必不可少的有机化合物。人体犹如一座极为复杂的化工厂,不断地进行着各种生化反应。酶要发挥其活性,必须有辅酶的参与。已知许多维生素是酶的辅酶或者辅酶的重要组成分子,人体基因多态性可能通过改变维生素的吸收、转运、生物转化等环节影响疾病的发生或维生素的保健或治疗效应。
1 维生素D
维生素D是一类脂溶性维生素,属类固醇化合物。天然的维生素 D有两种:麦角钙化醇(D2)和胆钙化醇(D3)。在人类所需的维生素中,维生素D非常特殊,它是激素的前体,而且在阳光充足的情况下,人体自身可以合成维生素D3。维生素D3由7-脱氢胆固醇经紫外线照射后进行光化学反应转变而成,动物皮肤细胞中含有7-脱氢胆固醇,所以多晒日光是获取维生素D的简易方法。但它的活性不高,必须经肝脏及肾脏作用转化成骨化三醇[1,25-二羟胆钙化醇,1,25-(OH)2-D3],1,25-(OH)2-D3是维生素 D3在体内活性形式[6],可以明显调节钙、磷的代谢活性,促进小肠粘膜对磷的吸收和转运,同时也促进肾小管对钙和磷的重吸收。
维生素D内分泌系统功能异常可引起甲状腺自身免疫性疾病。1,25-(OH)2-维生素D3通过免疫调节降低HLA-II类分子在甲状腺细胞的表达,还可抑制淋巴细胞增殖和炎症细胞因子的分泌,所以,在自身免疫性甲状腺疾病情况下血浆中的1,25-(OH)2-维生素 D3明显下降[7]。维生素 D结合蛋白(DBP)作为1,25-(OH)2-维生素 D3系统内的主要转运体,介导1,25-(OH)2-维生素 D3的细胞内吞作用,其基因微卫星序列多态性可通过调节与1,25-(OH)2-维生素 D3的亲和力,明显影响维生素D在体内的作用。Michael等发现,DBP基因第8内含子(TAAA)n可变重复序列变异基因多态性与Graves’发病明显相关。在DBP基因敲除小鼠体内,维生素D代谢明显受到影响[8]。在另一项包括健康男性和患骨质疏松相关椎骨骨折的男性(n=170)的研究中,在检测DBP基因内含子间可变串联重复序列(TAAA)n-Alu变异情况时,发现DBP-Alu*10和*11等位基因对骨质疏松症具有保护作用(分别 OR=0.39,P<0.0005;OR=0.09,P<0.007),即当患者携带19-20个重复拷贝数(基因型为9/10,9/11,10/10)时,循环系统中 DBP浓度和游离型维生素D浓度较高,因此,具有较高骨密度和较低的骨质疏松发生风险[9]。
2 维生素 E
维生素 E又名生育酚,是所有具有α-生育酚活性的生育酚和生育三烯酚及其衍生物的总称,属于脂溶性维生素,是人体内一种重要的抗氧化剂。食物中维生素E主要被小肠上部吸收,在血液中主要由β-脂蛋白携带,运输至各组织。维生素E的主要功能包括:防止多元不饱和脂肪酸及磷脂质被氧化,故可维持细胞膜的完整性;保护维生素 A不受氧化破坏,并加强其作用;降低血液中的过氧化脂质;防止血小板过度凝集;增进红血球膜稳定及红血球的合成;维持细胞呼吸等[10]。
维生素E在体内由富含甘油三酯(TG)的脂蛋白运输,载脂蛋白A5(APOA5)基因多态性可明显影响高甘油三酯血症的发生,因此,维生素E的体内转运可能受APOA5基因调节。在169名2型糖尿病患者中发现,APOA5-1131T-->C位点TC突变型杂合子体内维生素E含量明显升高约13%,维生素E高含量人群中TC基因型的发生率约为正常含量人群的 2.6倍[11]。
1型纤溶酶原激活物抑制物(plasminogen activator inhibitor type 1,PAI-1)是一种心血管疾病的独立危险因子,在2型糖尿病患者体内表达增高。PAI-1基因4G/5G多态性可调节PAI-1蛋白表达,进而影响心血管疾病发生。维生素E可有效降低体内PAI-1的含量。Testa等[12]研究发现,连续10周给予93名2型糖尿病患者每天500 IU剂量的维生素E,检测4G/5G多态性及 PAI-1含量,发现4G/4G和4G/5G基因型患者使用维生素E后PAI-1含量从第10周才开始降低,而5G/5G基因型患者在第5周时血浆PAI-1含量已经降低(P<0.01),说明PAI-1的4G等位基因影响PAI-1蛋白表达,5G/5G基因型患者对于维生素 E起效更快,使用维生素E预防心血管疾病可能取得更好的效果。
3 维生素K
维生素K是促进血液正常凝固及骨骼生长的重要维生素。维生素K和肝脏合成4种凝血因子(凝血酶原、凝血因子Ⅶ、Ⅸ及Ⅹ)密切相关,同时也是谷氨酸γ羧化反应的辅因子。如果缺乏维生素K,则肝脏合成的上述4种凝血因子为异常蛋白质,它们催化凝血的能力下降,凝血因子的γ-羧化将不能进行,会出现凝血迟缓。维生素K主要功能包括:促进血液凝固,是凝血因子γ-羧化酶的辅酶;凝血因子7、9、10的合成也依赖于维生素 K;参与骨骼代谢,维生素 K参与合成维生素K依赖蛋白质(BGP),BGP能调节骨骼中磷酸钙的合成[13]。老年人的骨密度和维生素 K呈正相关。
维生素K治疗抗凝过度在临床上呈现明显的个体间差异,维生素K环氧化物还原酶复合物亚基1(VKORC1)的多态性可解释部分该现象。将维生素K应用于抗凝过度的病人(INR≥4),实验者根据基础INR值给予2.5~5 mg维生素 K治疗,于 3、6、24、72 h复查 INR值,发现 VKORC1-1639G>A位点G等位基因携带者服用维生素K后3 h INR值降低程度明显强于AA型纯合子病人(P<0.001)。说明VKORC1-1639G>A基因多态性与维生素 K的急性促凝效果明显相关,G等位基因携带者体内INR值降低更迅速[14]。
4 维生素 B12
维生素B12,又称为钴胺素,含有金属元素钴,是唯一含有金属元素的维生素。在体内主要以腺苷钴胺素和甲基钴胺素形式存在,分别参与两类重要的生化学反应过程:①腺苷钴胺素作为甲基丙二酸单酰辅酶A变位酶的辅助因子将甲基丙二酰辅酶A转化为琥珀酰辅酶A;② 甲基钴胺素作为蛋氨酸合成酶的辅酶将同型半胱氨酸再甲基化转变为蛋氨酸。人体维生素B12主要来源于食物,经消化道微生物进行生物合成,然后通过咕啉结合蛋白(haptocorrin,HC)、内因子(IF)和钴胺传递蛋白II(TCII)完成体内吸收、转运和细胞摄取。维生素B12参与体内红细胞形成、DNA合成和髓磷脂神经鞘的维持等体内关键过程。维生素 B12缺乏可导致机体免疫力缺损、巨幼红细胞性贫血、冠心病、胃肠道和神经系统疾病,还可以导致高同型半胱氨酸血症,后者是心血管疾病的独立危险因子[15],高同型半胱氨酸血症还与非胰岛素依赖型糖尿病、终末期肾病、神经管缺损、阿尔采末病等发病相关[16-17]。
维生素B12与其他B族维生素不同,一般植物中含量极少,而仅有某些土壤中的细菌生成。萎缩性胃炎也可能导致维生素B12吸收不良。近年来,通过候选基因和全基因组关联(GWAS)分析法证实,数个基因多态性与维生素 B12的含量、体内代谢及疾病明显相关。岩藻糖基转移酶(fucosyltransferase 2,FUT2)基因多态性与维生素 B12血浆水平有关,rs602662、rs601338和rs492602等多个位点的作用已经得到证实,其中rs602662(G772A位点)作为被GWAS研究反复验证的位点,与维生素 B12血浆水平明显相关。在1 146名印度人中使用单碱基延伸法进行FUT2基因分型,考虑年龄、性别、饮食习惯、高血压、糖尿病和冠心病等伴随疾病在内进行回归分析,rs602662 AA型纯合子人群平均血浆维生素 B12含量为175.3 pmol·L-1,而GG型纯合子人群维生素B12含量明显降低,仅为 149.5 pmol·L-1,降低比率约为15%,而GA杂合子基因型人群居中,为152.7 pmol·L-1,血浆维生素 B12含量表现为明显的基因剂量效应(P=4.0×10-5)。同时,发现G772A突变纯合子基因型的素食主义者体内血浆维生素 B12水平明显降低,GG型的素食主义者较AA型非素食主义者而言体内血浆维生素 B12水平降低约20%(140.7 pmol·L-1vs 174.2 pmol·L-1)。该研究证实素食主义和FUT2基因多态性的交互作用可能是造成患者体内血浆维生素 B12水平明显降低的原因[18]。
当维生素B12以甲基钴胺素形式存在于体内时,作为蛋氨酸合成酶的辅酶,可转运来自于5-甲基四氢叶酸的甲基基团,维生素B12缺乏,导致同型半胱氨酸再甲基化转变为蛋氨酸的过程受阻,血循环中同型半胱氨酸水平升高,可能导致心血管疾病、异常妊娠等多种临床疾病。维生素 B12在血液中主要与咕啉结合蛋白和钴胺传递蛋白(transcobalamin)结合。前者结合约80%的血浆维生素 B12,与细胞摄取无关;后者仅结合20%的血浆维生素 B12,但却是细胞摄取维生素B12所必需,钴胺传递蛋白-维生素 B12复合物被细胞膜上特异性受体识别,并摄取进入胞内,而细胞膜上特异性受体不能识别和摄取体内游离的维生素 B12和咕啉结合蛋白-维生素B12复合物。因此,钴胺传递蛋白的功能性基因多态性可能影响维生素 B12复合物的细胞摄取过程。钴胺传递蛋白最常见突变是776C>G(造成脯氨酸被精氨酸取代),776GG变异型纯合子个体约占人群总数的20%,野生型纯合子776CC和突变型杂合子776CG分别占人群的30%和50%。研究发现,776 C>G变异不仅影响钴胺传递蛋白和维生素B12之间的亲和力,还影响钴胺传递蛋白-维生素B12复合物向组织内的转运能力。Miller等发现,776GG变异基因型人群体内咕啉结合蛋白-维生素 B12复合物浓度明显降低,总维生素 B12和钴胺传递蛋白结合的百分比降低,伴随甲基丙二酸(MMA)浓度在血浆中明显增高,说明776 C>G基因多态性可改变细胞对维生素B12的利用,加剧维生素 B12缺乏状况。von Castel-Dunwoody等[19]在359名年轻妇女中研究发现,776GG变异型纯合子血浆钴胺传递蛋白-维生素 B12复合物明显低于CC野生型个体(74 pmol·L-1vs 87 pmol·L-1,P=0.02),776 C>G变异通过调节血浆钴胺传递蛋白-维生素B12复合物浓度,影响体内同型半胱氨酸浓度,进而影响心血管等疾病的发生。
5 维生素C
维生素 C又称为L-抗坏血酸,是一种水溶性维生素,能够治疗坏血病,并且具有酸性,所以称作抗坏血酸。维生素C是人体内的高效抗氧化剂,用来减轻抗坏血酸过氧化物酶(ascorbate peroxidase)基底的氧化能力,同时还参与体内许多重要化合物的生物合成过程。维生素C对胶原合成具有重要作用,维生素C缺乏使动脉粥样硬化斑块中的胶原含量降低,从而易导致斑块破裂,严重者可导致血栓,甚至死亡。维生素C是最不稳定的一种维生素,在体内极易被氧化。植物及绝大多数动物均可在自身体内合成维生素C,可是人类因缺乏将L-古洛酸转变成为维生素C的酶类,不能合成,因此,必须从食物中摄取[20]。维生素C能够捕获自由基,因此能够预防癌症、动脉粥样硬化、风湿病等疾病。此外,它还能够增强免疫力[21]。
钠离子依赖型维生素 C转运体(sodium dependent vitamin C transporters,SVCT,编码基因为 SLC23A2)1和 2正常功能有助于维持维生素C的体内平衡。SVCT1主要分布于肠道和肾脏,控制维生素C的摄入和排出,SVCT2主要分布于高度活跃的代谢组织,保证如主动脉等特定组织细胞内抗坏血酸逆浓度梯度聚集。细胞内抗坏血酸是动脉管壁及斑块帽胶原合成的关键因子,同时,可起到减轻内皮功能障碍和炎症的作用,保护和稳定血管斑块。人类SVCT2基因多态性与多种肿瘤及早产有关[22]。SVCT2的2个多态性位点rs6139591和rs2681116可改变维生素 C的摄取和循环中抗坏血酸浓度。在一项大型临床研究中,根据对57 053名病例队列研究对象进行6.4年的随访,发现rs6139591 TT基因型的女性摄取食物中的维生素 C较少,患急性冠脉综合症的风险较rs6139591 CC基因型的患者增高5.39倍;rs1776964 TT女性纯合子相对于CC基因型患者而言,从食物中可摄取的维生素 C较多,优势比为3.45。说明SVCT2基因多态性与女性急性冠脉综合症有关,补充高含量维生素C食物可能成为预防该疾病风险的有效手段[23-24]。
6 叶酸
叶酸作为重要的一碳载体,在核苷酸合成、同型半胱氨酸再甲基化等重要生理功能中起重要作用,尤其是在快速的细胞分裂期和生长过程(如婴儿发育、怀孕)。叶酸能促进骨髓中的幼细胞发育成熟,形成正常形态的红细胞,从而避免贫血。
Meta分析显示,叶酸的体内主要代谢酶—亚甲基四氢叶酸还原酶基因(MTHFR)677位C>T基因多态性的TT基因型患心血管疾病的风险增高 14% ~21%[25-26]。Torre等[27]研究发现,叶酸与MTHFR 677C>T位基因多态性存在交互作用,共同影响结直肠癌的发生(P=0.037),体内叶酸含量低且同时为MTHFR 677TT基因型的人群具有更高的结直肠癌发病风险(OR=2.4),说明体内叶酸水平相对于MTHFR 677TT基因型的人群具有更重要的保健意义。
还原叶酸载体(reduced folate carrier,RFC-1,编码基因为SLC19A1)是一种叶酸的体内转运体,参与叶酸跨胎盘转运、血脑屏障吸收、肾小管通透等多个生理过程。胞内叶酸缺乏可引起DNA甲基化异常,最终与先天性疾病、肿瘤、心血管疾病及神经精神类疾病均可能相关。RFC-1基因80A>G基因多态性与血浆中同型半胱氨酸含量增加和叶酸水平降低有关,进而可能导致先天性疾病的发生[28]。一项针对930名唐氏综合症(DS)儿童的母亲及1 240例正常对照母亲的Meta分析发现,RFC-1基因80GG基因型发生DS风险增高1.27倍,单独G等位基因增加DS风险为1.14倍[29]。说明还原叶酸载体蛋白基因多态性可通过调节叶酸在胎儿的转运而影响其神经精神系统发育。
虽然,目前针对维生素的实验数据和人群研究尚不足以或根据基因多态性指导维生素的个体化使用,但是已经为维生素的药物基因组学研究提供了一种思路。本文中涉及的一些基因多态性以及Tab 1、2补充的与维生素体内代谢相关的基因多态性的研究结果是本综述对目前有关维生素基因多态性研究的一个简要总结。这些维生素的基因多态性的研究结果能否在不同人群和种族中得到重复验证还需要进一步研究。已有的基因多态性和维生素的交互作用的证据还不充分,许多研究存在观察时间短、样本量小、实验设计有缺陷等问题。今后的研究需要在此基础上改进,在实验设计、随机对照样本、严格的病例入选条件等方面进行完善。随着检测方法和手段的发展,希望在未来的几年,维生素的遗传药理学和药物基因组学研究可以取得可重复性的、高水平的科学证据,推动维生素的个体化进程。

Tab 1 Effect of genetic polymorphisms on fat-solution vitamins
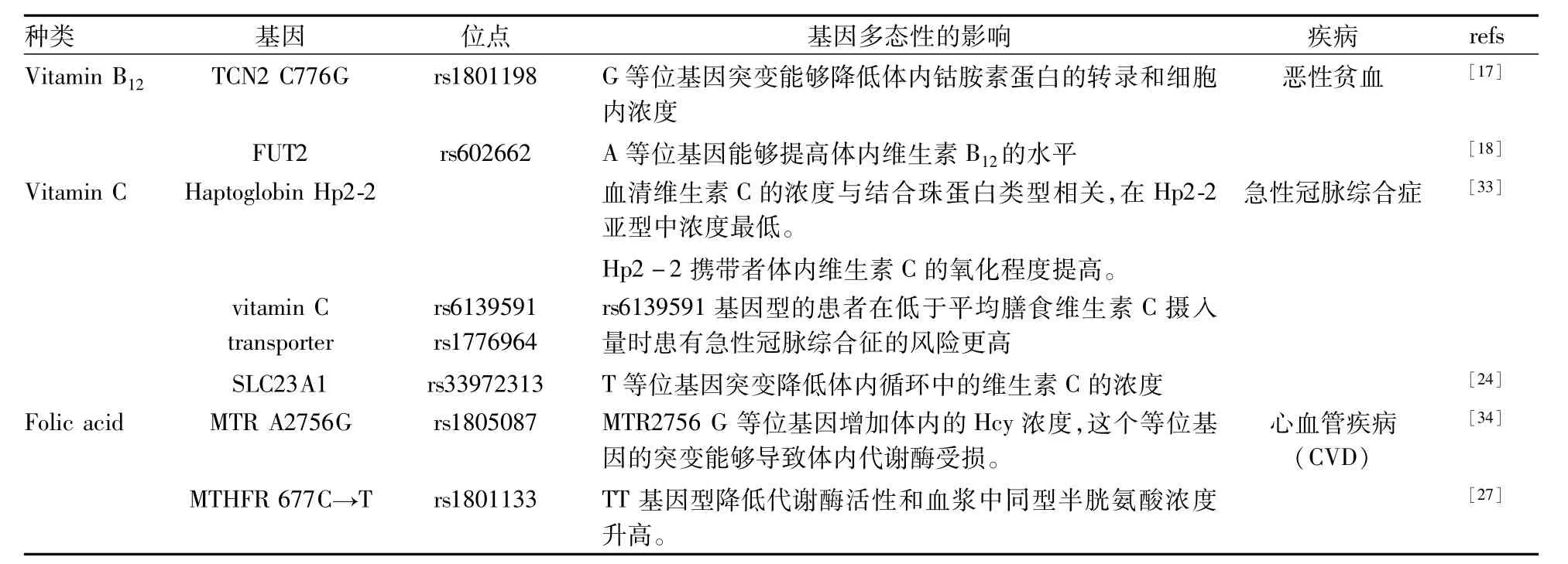
Tab 2 Effect of genetic polymorphisms on water-solution vitamins
参考文献:
[1] Krishna K D,Shewade D G,Loriot M A,et al.Effect of CYP2C9,VKORC1,CYP4F2 and GGCX genetic variants on warfarin maintenance dose and explicating a new pharmacogenetic algorithm in South Indian population[J].Eur J Clin Pharmacol,2014,70(1):47-56.
[2] Shaw K,Amstutz U,Hildebrand C,et al.VKORC1 and CYP2C9 genotypes are predictors of warfarin-related outcomes in children[J].Pediatr Blood Cancer,2014,61(6):1055-62.
[3] UK Collaborative HIV Cohort Study Steering Committee.HLA B*5701 status,disease progression,and response to antiretroviral therapy[J].AIDS,2013,27(16):2587-92.
[4] Schnitzler M.Diabetes mellitus after kidney transplantation in the United States[J].Am J Transplant,2003,3(10):1318-9.
[5] Cline J.Calcium and vitamin D metabolism,deficiency,and excess[J].Top Companion Anim Med,2012,27(4):159-64.
[6] Engelman C D,Meyers K J,Iyengar S K,et al.Vitamin D intake and season modify the effects of the GC and CYP2R1 genes on 25-hydroxyvitamin D concentrations[J].J Nut,2013,143(1):17-26.
[7] 江 巍,高凤荣.维生素 D缺乏相关性疾病研究进展[J].中国骨质疏松杂志,2014,20(3):331-7.
[7] Jiang W,Gao F R.The research progress between vitamin D and correlation disease[J].Chin J Osteoporos,2014,20(3):331-7.
[8] Pani M A,Regulla K,Segni M,et al.Vitamin D 1alpha-hydroxylase(CYP1alpha)polymorphism in Graves'disease,Hashimoto′s thyroiditis and type 1 diabetes mellitus[J].Eur J Endocrinol,2002,146(6):777-81.
[9] Al-oanzi Z H,Tuck S P,Mastana S S,et al.Vitamin D-binding protein gene microsatellite polymorphism influences BMD and risk of fractures in men[J].Osteoporos Int,2008,19(7):951-60.
[10]Rigotti A.Absorption,transport,and tissue delivery of vitamin E[J].Mol Aspects Med,2007,28(5-6):423-36.
[11] Hsu L A,Ko Y L,Chang C J,et al.Apolipoprotein A5 gene-1131T/Cpolymorphism is associated with the risk of metabolic syndrome in ethnic Chinese in Taiwan[J].Clin Chem Lab Med,2008,46(12):1714-9.
[12]Testa R,Bonfigli A R,Sirolla C,et al.Effect of 4G/5GPAI-1 polymorphism on the response of PAI-1 activity to vitamin E supplementation in type 2 diabetic patients[J].Diabetes Nutr Met,2004,17(4):217-21.
[13]Shearer M J,Fu X,Booth S L.Vitamin K nutrition,metabolism,and requirements:current concepts and future research[J].Adv Nutr,2012,3(2):182-95.
[14]Skov J,Bladbjerg E M,Leppin A,et al.The influence of VKORC1 and CYP2C9 gene sequence variants on the stability of maintenance phase warfarin treatment[J].Thromb Res,2013,131(2):125-9.
[15]冯晓婷.维生素B12缺乏与相关疾病的关系[J].中国实用神经疾病杂志,2014,17(1):96-8.
[15]Feng X T.Vitamin B12deficiency and related disease[J].Chin J Pract Dis,2014,17(1):96-8.
[16]Mohanty P K,Kapoor S,Dubey A P,et al.Evaluation of C677T polymorphism of the methylenetetra hydrofolate reductase gene and its association with levels of serum homocysteine,folate,and vitamin B12as maternal risk factors for Down syndrome[J].Indian J Hum Genet,2012,18(3):285-9.
[17]Castro R,Barroso M,Rocha M,et al.The TCN2 776CNGpolymorphism correlates with vitamin B(12)cellular delivery in healthy adult populations[J].Clin Biochem,2010,43(7-8):645-9.
[18]Tanwar V S,Chand M P,Kumar J,et al.Common variant in FUT2 gene is associated with levels of vitamin B(12)in Indian population[J].Gene,2013,515(1):224-8.
[19]von Castel-Dunwoody K M,Kauwell G P,Shelnutt K P,et al.Transcobalamin 776C->G polymorphism negatively affects vitamin B-12 metabolism[J].Am J Clin Nutr,2005,81(6):1436-41.
[20]Amr M,El-Mogy A,Shams T,et al.Efficacy of vitamin C as an adjunct to fluoxetine therapy in pediatric major depressive disorder:a randomized,double-blind,placebo-controlled pilot study[J].Nutr J,2013,12:31.
[21]李 能,陈忠东,周 波.维生素C体外抑制宫颈癌HeLa细胞株生长及其机制的研究[J].中国药理学通报,2005,21(9):1096-9.
[21]Li N,Chen Z D,Zhou B.The effect of vitamin Con the growth of cervical carcinoma HeLa cells in vivo[J].Chin Pharmacol Bull,2005,21(9):1096-9.
[22]Lucock M,Yates Z,Boyd L,et al.Vitamin C-related nutrient-nutrient and nutrient-gene interactions that modify folate status[J].Eur J Nutr,2013,52(2):569-82.
[23]Dalgard C,Christiansen L,Vogel U,et al.Variation in the sodiumdependent vitamin C transporter 2 gene is associated with risk of acute coronary syndrome among women[J].PLoS One,2013,8(8):e70421.
[24] Cahill L E,El-Sohemy A.Vitamin C transporter gene polymorphisms,dietary vitamin Cand serum ascorbic acid[J].J Nutrigenet Nutrigenomics,2009,2(6):292-301.
[25]McNulty H,Pentieva K,Hoey L,et al.Nutrition throughout life:folate[J].Int J Vitam Nutr Re s,2012,82(5):348-54.
[26]Diop-Bove N,Jain M,Scaglia F,et al.A novel deletion mutation in the proton-coupled folate transporter(PCFT;SLC46A1)in a Nicaraguan child with hereditary folate malabsorption[J].Gene,2013,527(2):673-4.
[27]Torre M L,Russo G T,Ragonese M,et al.MTHFR C677T polymorphism,folate status and colon cancer risk in acromegalic patients[J].Pituitary,2014,17(3):257-66.
[28]Wang Q,Li X,Ding Y,et al.The first Chinese case report of hereditary folate malabsorption with a novel mutation on SLC46A1[J].Brain Dev,2014,pii:S0387-7604(14)00032-1.
[29]Coppede F,Lorenzoni V,Migliore L.The reduced folate carrier(RFC-1)80A>G polymorphism and maternal risk of having a child with Down syndrome:a meta-analysis[J].Nutrients,2013,5(7):2551-63.
[30]Wang Z,Schuetz E G,Xu Y,et al.Interplay between vitamin D and the drug metabolizing enzyme CYP3A4[J].J Steroid Biochem Mol Biol,2013,136:54-8.
[31]Engelman CD,Meyers K J,Iyengar SK,et al.Vitamin D intake and season modify the effects of the GCand CYP2R1 genes on 25-hydroxyvitamin D concentrations[J].J Nutr,2013,143(1):17-26.
[32]Lippi G,Franchini M,Favaloro E J.Pharmacogenetics of vitamin K antagonists:useful or hype[J]?Clin Chem Lab Med,2009,47(5):503-15.
[33]Langlois MR,Delanghe M L,De Buyzere.Effect of haptoglobin on the metabolism of vitamin C[J].Am J Clin Nut,1997,66(3):606-10.
[34]Biselli JM,Zampieri B L,Goloni-Bertollo E M,et al.Genetic polymorphisms modulate the folate metabolism of Brazilian individuals with Down syndrome[J].Mol Biol Rep,2012,39(10):9277-84.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年2期)2022-03-29
中国生殖健康(2020年6期)2020-02-01
中国生殖健康(2019年12期)2019-01-07
中国生殖健康(2018年6期)2018-11-06
中国防痨杂志(2018年3期)2018-03-07
家庭科学·新健康(2017年8期)2017-08-17
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
作物研究(2014年6期)2014-03-01
中国神经精神疾病杂志(2013年4期)2013-03-11