
展示在毕赤酵母表面稻米脂肪酶的性质研究
2014-05-03 13:56朱婧谢定文李盛敏焦玲方芳
食品与机械 2014年3期
朱 婧 谢 定 文 李 盛 敏 焦 玲 方 芳
ZHU Jing XIE Ding WEN Li SHENG Min JIAO LingFANG Fang
(长沙理工大学化学与生物工程学院,湖南 长沙 410114)
(College of Chemical and Biology,Changsha University of Science and Technology,Changsha,Hunan 410114,China)
脂肪酶是重要的工业酶制剂品种之一,可以催化解脂、酯交换、酯合成等反应,广泛应用于食品、化工、造纸等工业生产,目前主要产自微生物,中国长期依赖于进口,价格奇高[1]。稻谷糊粉层中含有大量的脂肪酶,在稻谷碾米时,由于细胞壁被破坏,脂肪酶与稻谷中的油脂接触而迅速发生反应导致米糠酸败,因而稻米脂肪酶在粮食加工厂一直被当作有害物质,一般都要进行钝化或灭酶活处理后再进行油脂提取和其它深加工[2]。20世纪末,有学者[3]发现稻米脂肪酶具有对中短链脂肪酯解及手性反应催化活力高等特性,开始进行稻米脂肪酶的变废为宝的研究,采取的方法是将米糠脱脂后提取分离得到脂肪酶后再研究利用。
近些年,酵母菌的应用研究[4,5]越来越广,而利用酵母菌展示技术更使得脂肪酶的开发利用达到一个新的阶段。酵母菌表面展示脂肪酶具有提高脂肪酶的产量,在使用过程中不用分离纯化等优点,但并不是所有脂肪酶均能展示成功且具酶活性[6,7]。毕赤酵母(Pichaia pastoris)是一种新型的真核表达系统,可以高效表达外源蛋白而细胞内蛋白分泌极少,可以进行高密度发酵,糖基化程度较低,同时具备了原核生物生长速度快的优点,以及真核细胞蛋白翻译后的加工修饰功能,适用于大量表达有生物活性的外源蛋白[7,8],但目前尚未见酵母成功展示稻米脂肪酶及其应用的报道。本研究拟以毕赤酵母GS115作为表达宿主,异源表达稻米脂肪酶基因并初步研究了其酶学性质。
1 材料与方法
1.1 材料
1.1.1 主要仪器与设备
紫外—可见分光光度计:UV2600型,上海舜宇恒平有限公司;
琼脂糖凝胶电泳仪:DYCP-31DN型,北京天驰仪诚生物科技有限公司;
凝胶成像仪:DOC 2000型,美国Bio-Red公司;
恒温摇床:NS-2102C型,天津市欧诺仪器仪表有限公司。
1.1.2 菌株和质粒
pPIC9K:美国Invitrogen公司;
毕赤酵母(Pichia pastoris)GS115宿主菌:中国农业微生物菌种保藏管理中心。
1.1.3 工具酶、试剂与试剂盒
限制性内切酶 EcoR I、Not I、Mlu I、Sac I,T4 DNA连接酶,DNA Marker等:大连宝生物科技有限公司(takara);
质粒小量提取试剂盒、胶回收试剂盒、PCR纯化试剂盒:德国Qiagen公司;
对硝基苯酚丁酸酯(p NPB)、PEG3350、LiCl:美国Sigma公司;
YPD培养基、BMGY培养基、BMMY培养基、YNB培养基:北京索莱宝生物科技有限公司;
其他为进口分装或国产分析纯。
1.1.4 GCW14-RL基因 根据张轩薇等[8]的 GCW14序列以及Chuang等[9]的Rlipase序列,设计了 GCW14-RL融合基因序列,GCW14-RL基因的5’端添加了Eco RI限制性内切酶位点,3’端添加了Not I位点,GCW14和RL之间是Mlu I位点。Eco RI-GCW14-Mlu I-RL-Not I基因由南京金斯瑞生物科技有限公司合成并克隆到p UC57载体的多克隆位点上,命名为p UC57-GCW14-RL。
1.2 方法
1.2.1 pPIC9K-GCW14-RL载体的构建 分别将p UC57-GCW14-RL和p PIC9K载体进行Eco RI和 Not I双酶切反应,37℃水浴4 h之后用1.2%琼脂糖凝胶电泳鉴定。采用胶回收试剂盒回收目的基因片断GCW14-RL和pPIC9K载体片段。之后用T4 DNA Ligase进行连接反应,22℃过夜连接,之后转化到DH5a感受态细胞中,涂布在含有amp和kana抗性的LB固体培养基上37℃过夜培养。挑取克隆进行摇菌,质粒抽提,并进行Eco RI和Not I双酶切反应,之后用1.2%琼脂糖凝胶电泳鉴定[10]。将阳性克隆命名为pPIC9K-GCW14-RL。
1.2.2 pPIC9K-GCW14-RL转化到毕氏酵母GS115中
(1)煮沸1 m L鲑鱼精DNA 5 min,迅速冰浴以制备单链担体DNA;
(2)将pPIC9K-GCW14-Rl用Sac I内切酶进行线性化处理;
(3)将感受态酵母菌离心,以Tips去除残余的LiCl溶液;
(4)对于每一个转化,按以下顺序加入:50%PEG3350,240μL;1 M LiCl,36μL;2 mg/m L 单 链 Salmon sperm DNA,25μL;5~10μg/50μL H2O 质粒DNA,50μL。
(5)剧烈旋涡混匀直至沉淀菌体完全分布均匀(约1 min);
(6)30℃水浴孵育30 min;
(7)42℃水浴热休克20~25 min;
(8)6 000~8 000 r/min离心收集酵母菌体;
(9)重悬酵母于1 m L YPD培养基,30℃摇床孵育;
(10)1~4 h后,取25~100μL菌液铺YD选择性培养基平板(1.67%YNB+2%dextrose+1%琼脂粉),于30℃培养2~3 d鉴定(将表达菌株和对照菌株在固体培养基平板,30℃培养至转化子出现)。因为GS115是组氨酸缺陷型菌,YD培养基里面没有组氨酸,因此GS115在YD培养基上不能生长,而pPIC9K带组氨酸合成酶基因,只有成功转化了pPIC9k-GCW14-RL的GS115菌才能在YD培养基上生长。
1.2.3 粗酶液制备 将毕赤酵母重组菌接种至YPD液体培养基中,30℃,250 r/min培养24 h,按1%的接种量转接到新鲜的BMGY培养基中,30℃,250 r/min培养5 d,每隔24 h加甲醇诱导,收集发酵菌体,用p H 8.0缓冲液洗涤3次,用少量缓冲液重悬,即得重组脂肪酶粗酶液。
1.2.4 脂肪酶活力测定 采用对硝基苯酚法测定脂肪酶的活力[8-11],酶活力测定反应体系总体积为10 m L,包括1 m L p NPB底物溶液(50 m M)以及9 m L 50 m M Tris—HCl(p H 8.0);反 应 体 系 混 匀 后 在 加 热 振 荡 器 中 35 ℃,800 r/min预热5 min后,加入底物进行反应,10 min后立即12 000 r/min离心终止反应;反应结束后取上清液在405 nm处测定吸光度,只加底物跟缓冲液的样品为空白,设3个平行,记录A405值。一个酶活力单位为每分钟水解所加底物生成1μmol p NP所需的酶量。
1.2.5 最适温度 以p H 8.0的Tris—HCl缓冲液重悬菌体,在30,35,40,45,50,55,60,65,70℃分别保温10 min后,以p NPB为底物,测定重组毕赤酵母在35℃下的水解活性。
1.2.6 热稳定性 以p H 8.0的Tris—HCl缓冲液重悬菌体,在50 ℃下保温0.5,1.0,1.5,2.0,2.5,3.0 h后,以p NPB为底物,测定重组毕赤酵母在35℃下的水解活性。
1.2.7 最适p H 分别以p H 6.0,6.5,7.0的磷酸缓冲液,p H 7.5,8.0,8.5的 Tris—HCl缓冲液,p H 9.0,9.5,10.0的甘氨酸缓冲液洗涤菌体3次,并重悬,20℃保温24 h,再用p H 8.0的Tris—HCl缓冲液洗涤3次,p NPB为底物,测定重组毕赤酵母在35℃下的水解活性。
1.2.8 固定化效果 将使用过后的酵母菌体用缓冲液洗涤3次并重悬,重复1.2.4的步骤测重组酵母菌的脂肪酶活力。
2 结果与分析
2.1 pPIC9k-GCW14-RL载体的构建结果
p UC57-GCW14-RL的 双 酶 切 结 果 见 图1(a)。回 收GCW14-RL与pPIC9k载体连接后的质粒pPIC9k-GCW14-RL双酶切鉴定结果见图1(b),得到了预期大小的两个片段。说明成功将GCW14-RL亚克隆到了pPIC9k载体。

图1 琼脂糖凝胶电泳结果Figure 1 Agarose gel electrophoresis
2.2 pPIC9k-GCW14-RL载体转化 GS115的结果
将pPIC9k-GCW14-RL质粒转化到GS115感受态的结果见图2。由于GS115是组氨酸缺陷型菌,因此在YD培养基上不能生长,只有成功转化了pPIC9K的GS115菌才能在YD培养基上生长。

图2 GS115在YD培养基上的生长情况Figure 2 Grouth of GS115 in YD medium
2.3 酵母表面展示稻米脂肪酶的最适温度及热稳定性
测定不同温度下该重组脂肪酶的酶活,由图3可知,该重组脂肪酶最适作用温度为35℃,温度30~45℃范围内都能保持80%以上的相对酶活力。比RL游离酶的最适温度[10]低15℃。但该重组脂肪酶在50℃保温1 h仍能保持97.3%的相对酶活性,在50℃保温3 h后仍能保持80%以上相对的酶活力,证明该酶在50℃较高温度下仍有较好的温度稳定性(图4)。
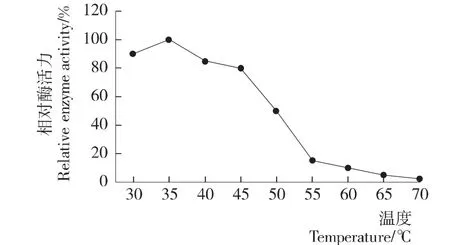
图3 酵母表面展示稻米脂肪酶的最适温度Figure 3 Optimum of temperature on the activity of surface displayed RL

图4 酵母表面展示稻米脂肪酶的热稳定性Figure 4 Thermal stablity of the activity of surface displayed RL
2.4 酵母表面展示稻米脂肪酶的最适p H
测定不同p H缓冲液条件下的酶活绘制曲线见图5。该重组脂肪酶最适p H 为8.0,在p H 7.0到p H 10.0范围内都能保持80%以上的酶活力。与RL游离酶的最适p H[8]基本一致。但p H低于7.0时,酶活下降较快,p H为6.0时,酶活只有最高酶活的52%。
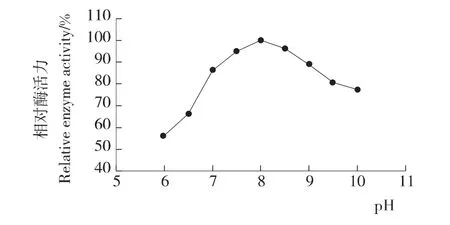
图5 酵母表面展示稻米脂肪酶的最适p HFigure 5 Effect of p H on the activity of surface displayed RL
2.5 酵母表面展示脂肪酶的固定化效果
酶的操作稳定性是固定化酶的重要特性之一,酵母展示稻米脂肪酶省去了酶的固定化操作却具有可重复使用的固定化酶的特性。图6显示了酵母展示稻米脂肪酶的固定化效果,该重组脂肪酶在初次使用后,酶活性基本没损失,重复使用5次之后仍具有80%以上的相对酶活力,活性降低的原因可能是因为每次重复回收有少量的酵母展示脂肪酶的损失以及回收过程中多次冲洗导致活性降低以及反应过程中酶部分热失活所致。
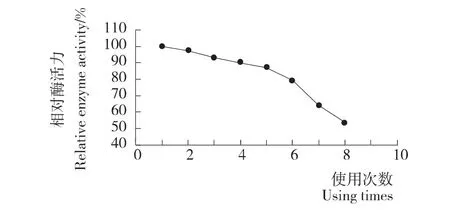
图6 酵母表面展示稻米脂肪酶的使用次数与相对活性Figure 6 Activity of surface displayed RL after repeated use times
3 结论
稻米脂肪酶对油脂具有很好的酯解活性,过去由于其难于分离提纯而被废弃。本试验将来自稻米脂肪酶(RL)的基因转化到毕赤酵母GS115宿主菌中,通过摇瓶发酵,测定其最适作用温度为35~45℃,最适p H为8.0,与游离RL相差不大,而且酵母菌通过高密度发酵可获得大量具有RL活性的菌体,不需要特别的分离提纯和固定化处理,其重复使用5次依然可以保持80%以上的活性,这就为拓广稻米脂肪酶的利用提供了依据。
1 童志勇.脂肪酶国产化开启绿色合成新篇章[J].现代化工,2009,29(5):91.
2 吕莹果,季慧,张晖,等.米糠资源的综合利用[J].粮食与饲料工业,2009(4):19~22.
3 Fadnavis N W,Jadhav V.Enantioselective hydrolytic reactions of rice bran lipase(RBL):a first report[J].Tetrahedron:A-symmetry,1997,8(14):2 361~2 366.
4 武运,葛凤,陈卫林,等.酿酒废酵母吸附共存重金属离子的研究[J].食品与机械,2012,28(4):73~76.
5 刘文山.脂肪酶在酿酒酵母中的表面展示研究[D]武汉:华中科技大学,2010.
6 Tanaka T,Yamada R,Ogino C,et al.Recent developments in yeast cell surface display toward extended applications in biotechnology[J].Appl Microbial Biotechnol,2012,95(8):577~591.
7 Brunel L,Neugnot V,Landucci L,et al.High-level expression of Candida parapsilosis lipase/acyltransferase in Pichia pastorise[J].Journal of Biotechnology,2004,111(1):41~50.
8 张轩薇,叶燕锐,刘晓肖,等.毕赤酵母新启动子GCW14在细胞表面展示CALB 中的应用[J].生物技术通报,2013(4):129~135.
9 Chuang H H,Chen P T,Wang W N,et al.Functional proteomic analysis of rice bran esterases/lipases and characterization of a novel recombinant esterase[J].Journal of Agricultural and Food Chemistry,2011,59(2):2 019~2 025.
10 王盛,金大勇,蔡宇杰,等.谷胱甘肽生产菌重组巴斯德毕赤氏酵母的构建[J].食品与机械,2007,23(6):17~19.
11 Teng Y,Xu Y.A modified para-nitrophenyl palmitate assay for lipase synthetic activity determination in organic solvent[J].Analytical Biochemistry,2007,363(2):219~224.
猜你喜欢
美食(2022年5期)2022-05-07
少儿科学周刊·儿童版(2021年21期)2021-12-11
中国粮食经济(2018年5期)2018-12-27
食品与发酵工业(2018年3期)2018-04-12
中国洗涤用品工业(2017年2期)2017-04-16
广东饲料(2016年1期)2016-12-01
工业微生物(2016年5期)2016-11-11
化学与生物工程(2016年10期)2016-11-10
国外医药(抗生素分册)(2016年2期)2016-07-12
中国塑料(2016年7期)2016-04-16
