
一株产酸性α-淀粉酶菌株的筛选及酶学性质研究
2014-04-24 13:25:12权淑静解复红刘德海陈国参
中国酿造 2014年5期
权淑静,解复红,马 焕,刘德海 *,陈国参
(1.河南省工业酶工程技术研究中心,河南 郑州 450008;2.河南省科学院生物研究所有限责任公司,河南 郑州 450008)
α-淀粉酶是一种催化淀粉内部α-1,4-糖苷键水解,释放出葡萄糖及不同长度直链或支链低聚糖的胞外酶。该酶广泛应用于淀粉深加工、酿酒、酒精生产、食品、纺织、饲料及医药等行业。按照酶的活性、最适pH可分为酸性、碱性和中性α-淀粉酶3种,大多数α-淀粉酶为中性[1],在酸性条件下酶活明显降低,已不能满足一些酸性条件下淀粉原料的深加工工艺的要求。
随着商业需求的增加,通过α-淀粉酶生产高质量麦芽糖的需求也在不断增加[2],目前用于淀粉加工的淀粉酶最适pH值为6.8,液化过程需要将淀粉浆从其原有的pH值为3.2~4.5提高至5.8~6.2,此外,添加Ca2+以提高酶的活性和稳定性。接下来的糖化步骤需要调整pH值至4.2~4.5。这些步骤耗费时间和成本。因此,需要适应极端酸性环境的高活性淀粉酶产品[3-4]。
近年来,国内外对耐酸性α-淀粉酶进行了广泛的研究,并相继发现多种细菌、真菌、酵母菌可产生耐酸性α-淀粉酶,最适pH大多在4.0~6.0。我国开展相关研究起步较晚,目前主要集中在菌种选育和构建基因工程菌等方面,所选菌株的产酶活力不高且缺乏可靠的安全性。为了满足工业化生产需要,本实验从酒曲中筛选一株产耐酸性α-淀粉酶的菌株,并对酶的组成成分和酶学性质进行了研究,为工业化应用提供重要的基础理论。
1 材料与方法
1.1 材料与试剂
1.1.1 样品来源
河南省王勿桥醋业有限公司酿造用砖曲。
1.1.2 培养基
富集培养基:马铃薯200g/L,葡萄糖20g/L,1%链霉素3mL/L。
平板分离培养基:KH2PO41.0g/L,MgSO4·7H2O 0.5g/L,蛋白胨5.0g/L,葡萄糖10.0g/L,琼脂18.0g/L,pH5.0,1%孟加拉红水溶液3.3mL/L。临用时每100mL培养基中加1%链霉素液0.3mL。
液体发酵培养基:可溶性淀粉40.0g/L,尿素2.0g/L,蛋白胨2.0g/L,KH2PO43.0g/L,CaCl20.1g/L,FeSO4·7H2O 0.01g/L,MgSO4·7H2O 1.0g/L,pH5.0。
斜面保藏采用马铃薯葡萄糖琼脂(potato dextrose agar,PDA)培养基:马铃薯200g/L,葡萄糖20g/L,琼脂18g/L。
1.1.3 试剂
2×TaqPCR MasterMix:天根生化科技(北京)有限公司;通用引物NS1、NS8:深圳华大基因科技有限公司;Trans2K plus:北京全式金生物技术有限公司;琼脂糖:北京恒奥生物科技有限公司;蛋白胨:英国OXOID公司;其他试剂为国产分析纯。
1.2 仪器与设备
Multifuge X1R离心机:德国Thermo公司;Phenom proX台式扫描电镜:美国FEI公司;T6新世纪紫外可见光分光光度计:北京普析通用仪器有限公司。
1.3 实验方法
1.3.1 产耐酸性α-淀粉酶真菌菌株的筛选
平板分离:将砖曲在30W紫外灯下照射30min表面消毒,用无菌刀在内部多点取样并混匀。取1g粉末于富集培养基中,30℃、180r/min富集培养24h。取培养液分别稀释成10-2、10-3、10-4、10-5、10-6五个不同梯度与平板分离培养基混合倒平板,30℃恒温培养3~5d,挑取单菌落保藏至斜面。
菌株筛选:将分离到的菌株接入液体发酵培养基,30℃、180r/min培养3d后测酶活。将酶活最高的菌株作为进一步研究的出发菌株。
粗酶液的制备:将发酵液经4层灭菌纱布过滤后,8 000r/min离心10min,取上清液。
酶活测定方法参照文献[5]:180μL底物加入20μL酶液,对照(CK)为20μL缓冲液,在一定温度条件下反应10min,加入200μL 3,5-二硝基水杨酸(dinitrosalicylic acid,DNS)试剂,沸水浴5min终止反应,冷却后加蒸馏水2.1mL,在波长520nm条件下测定吸光度值。
酶活力定义:70℃、pH 3.6条件下每分钟从1%可溶性淀粉中释放1μmol葡萄糖所需的酶量为一个酶活单位(U)。
相对酶活的测定:相对酶活是以同组实验中活性最高点的值为100%,其余实验点的值与该点的值之比,通常以百分数表示。
1.3.2 菌株的鉴定
(1)形态观察[6]。
将分离纯化的菌株点种在PDA固体培养基平板上30℃培养48h,观察其菌丝生长情况,菌落形态、边缘、菌丝长度和颜色(正面、反面);直接制片观察法观察孢子、菌丝形态。
(2)分子生物学鉴定
真菌DNA的提取采用十六烷基三甲基溴化铵法(cetyltrimethyl ammonium bromide,CTAB)法[7]。
菌株18S rDNA:PCR扩增采用引物为:NS1(5′-GTA GTCATATGCTTGTCTC -3′);NS8(5′-TCCGCAGGTTC ACCTACGGA-3′)。
PCR扩增体系为50μL,包括无菌双蒸水19μL,正向和反向引物各2.0μL,模板DNA 2.0μL,2×TaqPCR MasterMix 25μL。
反应体系:每个反应30个循环,每个循环包括94℃变性30s,54℃退火30s,72℃延伸1.5min;最后72℃延伸10min,首次循环在95℃预变性2min。
将聚合酶链反应(polymerase chain reaction,PCR)扩增产物经琼脂糖凝胶电泳检测后,送华大基因测序。通过GenBank数据做相似性分析。
1.3.3 粗酶液酶学性质初步研究
酶反应最适温度:将离心后的粗酶液稀释一定倍数,与底物在不同温度下进行反应,温度梯度设置为30℃、40℃、50℃、60℃、70℃、80℃,测定酶活,确定最适温度。
酶反应温度稳定性:将稀释适当倍数的粗酶液分别在30℃、40℃、50℃、60℃、70℃、80℃保温1h,最适温度条件下测定剩余酶活。
酶反应最适pH:用pH分别为3.0、3.6、4.0、4.6、5.0、5.6、6.0、6.6、7.0的缓冲液配制底物1%淀粉溶液,用相应pH的缓冲液测定酶活(NaH2PO4-柠檬酸缓冲体系)。
酶反应pH稳定性:将酶液用相应pH的缓冲液稀释至适当倍数,在50℃保温1h,70℃测定剩余酶活。
金属离子对酶活力的影响:在反应体系中分别加入2mmol/L的乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)、Mg2+、Ca2+、Ba2+、Zn2+、Fe3+、Mn2+、K+、Co2+,测定酶活。
2 结果与分析
2.1 产酸性α-淀粉酶淀粉酶菌株筛选
经过筛选获得了6株酶活较高的产α-淀粉酶菌株,粗酶液的酶活见表1。由表1可以看出,A-5-3的酶活最高,达到282.38U/mL,因此确定A-5-3为出发菌株。

表1 产α-淀粉酶菌株筛选结果Table 1 Screening results of strain with α-amylase high yield
2.2 菌种鉴定
2.2.1 形态学观察
将菌株A-5-3接种至PDA培养基上培养2d,观察其形态特征结果见图1。由图1可知,菌落生长速度快,菌落初期为淡黄色,有褶皱;后期菌落中央黄色,边缘为白色菌丝,背面黄色。产生大量黑色分生孢子。结合显微观察和电镜下观察结果见图2,初步鉴定为黑曲霉(Aspergillus niger)。

图1 菌株A-5-3菌落形态特征Fig.1 Colonial morphological properties of strain A-5-3

图2 分生孢子梗、分生孢子头及分生孢子的扫描电镜图Fig.2 Scanning electron microscope of conidiophore,conidial head and conidia
2.2.2 18s rDNA基因的PCR扩增
以提取的A-5-3的DNA为模板,以NS1、NS8为引物进行PCR扩增。对扩增产物进行琼脂糖凝胶电泳分析,结果如图3,得到一条1 500bp左右的片段。将PCR产物送华大基因测序,结果显示该片段长1 448bp,将测序结果用美国国家生物技术信息中心(national center of biotechnology information,NCBI)的Blast软件进行核苷酸同源性比对,得知该菌株与黑曲霉的18S rDNA的序列同源性为100%,因此将该菌株归属为黑曲霉(Aspergillus niger)。

图3 PCR扩增的18S rDNA电泳图Fig.3 Electrophoretogram of PCR amplified 18S rDNA
2.3 菌株A-5-3产酸性淀粉酶性质研究
2.3.1 温度对酶活力的影响
在30℃、40℃、50℃、60℃、70℃、80℃测定粗酶液酶活,测得淀粉酶活性最大值为100%,结果见图4。
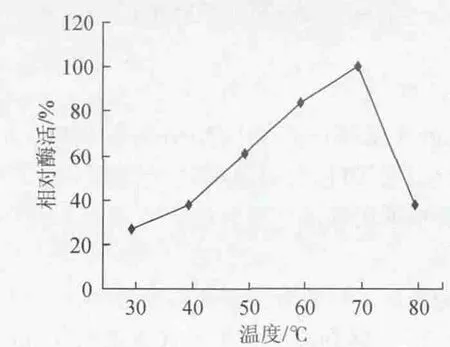
图4 温度对酶活力的影响Fig.4 Effect of temperature on enzyme activity
由图4可知,菌株A-5-3所产淀粉酶反应的最适温度为70℃。
2.3.2 酶的热稳定性研究
粗酶液在30℃、40℃、50℃、60℃、70℃、80℃保温1h后,进行酶促反应,以不保温酶液为对照,对照相对酶活值为100%,结果见图5。
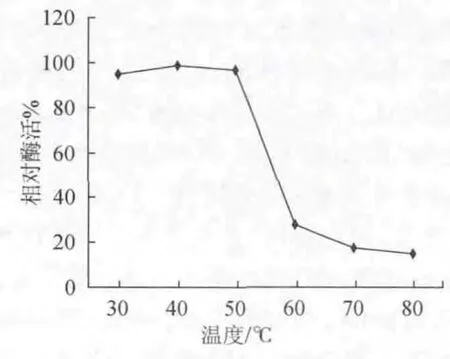
图5 温度对酶的稳定性影响Fig.5 Effects of temperature on enzyme stability
由图5可以看出,在30℃、40℃、50℃条件下,该酶的热稳定性好,保温1h后,剩余的相对酶活分别达到95.53%、98.55%、97.11%。当温度高于60℃时,酶活下降很快,到80℃时,剩余淀粉酶活力仅剩下最高酶活力的15%。
2.3.3 pH值对酶活力的影响
在pH 3.0、3.6、4.0、4.6、5.0、5.6、6.0、6.6、7.0条件下测酶活,结果见图6。以活性最大值为100%,由图6可知,酶的最适pH为3.6,pH 4.0、4.6时相对酶活分别为99.73%和99.69%,pH>5.0时酶活迅速降低,pH值为3.0时相对酶活为71.12%。
2.3.4 酶的pH稳定性研究
将酶液用不同pH值的缓冲液处理后测剩余酶活,结果见图7。以未经过处理的原酶液活力为100%,考察该酶的pH稳定性,由图7可知,该酶在pH 3.0~5.0非常稳定,相对酶活≥85%。在pH>6.0条件下,相对酶活力降低至50%以下。

图6 pH值对酶活力的影响Fig.6 Effect of pH on enzyme activity

图7 pH值对酶的稳定性影响Fig.7 Effect of pH on enzyme stability
2.3.5 金属离子对酶活力的影响
在反应体系中加入一定浓度的EDTA、MgCl2、CaCl2、BaCl2、ZnCl2、FeCl3、MnCl2、KCl、CoCl2溶液,使其在反应体系中的浓度达到2mmol/L,以未处理原酶液为对照(CK)100%,考察金属离子对酶活力的影响,结果见图8。由图8可知,Mn2+、K+对酶活有明显的激活作用,相对酶活达到153%和121%;EDTA对酶活有明显的抑制作用,相对酶活降至81%;Ca2+及其他金属离子对酶活没有明显影响。

图8 金属离子对酶活力的影响Fig.8 Effect of metal ions on enzyme activity
3 结论
α-淀粉酶是水解淀粉酶类,是商品化生产最早、应用最早、应用范围最广和用量最大的工业酶类之一。耐酸性α-淀粉酶作为一种极端环境淀粉酶,可广泛应用于发酵、纺织、饲料、医药等多种领域,可以明显地提高收得率,降低消耗,节约工业成本,具有很大的应用潜力和开发前景。目前工业用的酸性α-淀粉酶大多来源于微生物,产酸性α-淀粉酶的微生物主要是芽孢杆菌和曲霉[8],如酸热脂环酸芽孢杆菌(B.acidocaldarius)A-2[9],酸热脂环酸芽孢杆菌(B.acidocaldarius)ATCC2709[10],地衣芽孢杆菌(B.licheniformis)[11],酸热脂环酸芽孢杆菌(B.acidocaldarius)101[12],酸热脂环酸芽孢杆菌(Alicyclobacillus acidocaldarius)[13],黑曲霉(Aspergillus niger)[14],白曲霉(Aspegillus kawachii)[15],疏绵状嗜热丝孢菌(Thermomyoes lanuginosus)[16],嗜热脂肪芽孢杆菌(Bacillus steorothermopilius)[17]等。黑曲霉是最早发现能够产耐酸性α-淀粉酶的菌株,在日本、欧洲等国已经有用其进行工业化应用的报到。虽然这些微生物都能产生酸性α-淀粉酶,但是不同菌株产生的酶在耐酸性,耐热性及淀粉的水解程度均有差异。大多数的酸性α-淀粉酶的最适反应温度为50~60℃,其最适pH值为4.0~5.0。
本研究从酿造酒曲中分离筛选出的高产组选育的α-淀粉酶菌株A-5-3,经初步鉴定为黑曲酶(Aspergillus niger)。通过对其产生α-淀粉酶的酶学性质的研究,表明该酶的最适作用温度为70℃,热稳定性良好;最适作用pH值为3.6,该酶在pH 3.0~5.0范围内酶活稳定,当pH 3.0时,有71%的酶活;Mn2+、K+离子对此酶有激活作用,EDTA有明显的抑制作用,Ca2+对该酶没有明显的作用。与国内已报道文献相比,该酶的活力较高,且不依赖Ca2+调控,能够实现在弱酸性条件下淀粉液化和糖化的同步进行,从而简化了淀粉加工工艺,节约了成本,具有良好的应用前景。
[1]PANDEY A,NIGAM P,SOCCOL C R,et al.Advances in microbial amylases[J].Biotechnol Appl Bioc,2000,31(2):135-152.
[2]FOGARTY W M,COLLINS B S,DOYLE E M,et al.The high maltose-formingα-amylase ofSaccharomonospora viridis:mechanisms of action[J].J Ind Microbiol,1993,11(3):199-204.
[3]SHIVARAMAKRISHNAN S,GANGADHARAN D,NAMPOOTHIRI K M,et al.α-amylases from microbial sources-an overview on recent developments[J].Food Technol Biotech,2006,44(2):173-184.
[4]ARCHANA S,SATYANARAYANA T.Microbial acid-stableα-amylase:characteristics,genetic engineering and applications[J].Process Biochem,2013,48(2):201-211.
[5]XIE F H,QUAN S J,LIU D H,et al.Purification and characterization of a novelα-amylase from a newly isolatedBacillus methylotrophicusstrain P11-2[J].Process Biochem,2014,49(1):47-53.
[6]魏景超.真菌鉴定手册[M].上海:上海科技出版社,1979.
[7]孙立夫,张艳华,裴克全.一种高效提取真菌总DNA 的方法[J].菌物学报,2009,28(2):299-302.
[8]MORIMURA S,ZHANG W X,LCHIMURA T,et al.Genetic engineering of white shochu-koji to achieve higher levels of acid-stableα-amylase and glucoamylase and other properties when used for shochu making on a laboratory scale[J].J I Brewing,1999,105(5):309-314.
[9]KANNO M.A Bacillus acidocaldarius alpha-amylase that is highly stable to heat under acidic conditions[J].Agr Biol Chem,1986,50(1):23-31.
[10]CRABB W D,MITCHINSON C.Enzymes involved in the processing of starch to sugars[J].Trends Biotechnol,1997,15(9):349-352.
[11]Oziengbe E O,Onilude A A.Production of a thermostableα-amylase and its assay usingBacillus licheniformisisolated from excavated land sites in Ibadan,Nigeria[J].Bayero Journal of Pure and Applied Sciences,2012,5(1):132-138.
[12]BUONOCORE V,CAPORALE C,DE ROSA M,et al.Stable,inducible thermoacidophilic alpha-amylase fromBacillus acidocaldarius[J].JBacteriol,1976,128(2):515-521.
[13]EEKERT K,SCHNEIDER E.A thermoacidophilic endoglucanase(CelB) fromAlicyclobacillus acidocaldariusdisplays high sequence similarity to arabinofuranosidases belonging to family 51 of glycoside hydrolases[J].Eur J Biochem,2003,270(17):3593-3602.
[14]OKOLO B N,EZEOGU L I,MBA C N.Production of raw starch digesting amylase byAspergillus nigergrown of native starch sources[J].J Sci Food Agr,1995,69(1):109-115.
[15]SUDO S,ISHIKAWA T,SAKAMOTO Y,et al.Characteristics of acid-stableα-amylase production by submerged culture ofAspergillus kawachii[J].J Ferment Bioeng,1993,76(2):105-110.
[16]PETROVA S D,LLIEVA S Z,BAKALOVA N G,et al.Production and characterization of extracellularct-amylase from the thermophilie fugusThermomyees lanuginosus(wild and mutant strains)[J].Biotechnol Lett,2000,22(20):1619-1624.
[17]KAMASAKA H,SUGIMOTO K,TAKASHI K,et al.Bacillus stearothermophilusneopullulanase selective hydrolysis of amylose to maltose in the presence of amylopectin[J].Appl Environ Micro,2002,68(4):1658-1664.
猜你喜欢
农药科学与管理(2019年6期)2019-11-23 08:17:12
农药科学与管理(2019年6期)2019-11-23 08:17:06
农药科学与管理(2019年8期)2019-11-23 08:04:46
中国粮油学报(2019年4期)2019-07-12 09:06:32
湖南大学学报·自然科学版(2016年12期)2017-05-12 10:41:26
当代化工研究(2016年7期)2016-03-20 16:21:54
现代纺织技术(2015年6期)2015-02-28 14:03:19
食品工业科技(2014年15期)2014-03-11 18:17:52
食品工业科技(2014年9期)2014-03-11 18:15:55
食品工业科技(2014年7期)2014-03-11 18:14:49
