
伏牛山油松群落格局与动态研究
2014-04-08 01:53:57袁志良任思远李廘鑫
河南农业大学学报 2014年3期
潘 娜, 袁志良, 任思远 , 李廘鑫, 王 婷
(1.河南农业大学林学院,河南 郑州 450002; 2.河南农业大学生命科学院,河南 郑州 450002)
空间分布格局是种群个体在水平空间的分布方式或者在水平空间上种群个体之间的相互关系[1],也是种群生态特性、种内种间关系及生物与环境相互作用等综合作用的结果[2]. 种群个体的空间分布一直是生态理论研究的核心[3~5],也是探索植物群落结构组成和动态变化机制的关键[6].对空间分布格局的研究,将有助于理解种群的动态变化、种间的竞争与共存机制及对森林群落的影响[7,8].因为1个种在某些尺度上可能服从聚集性分布,而在其他尺度上则可能变成随机分布或均匀分布[9],所以植物种群的空间分布格局的研究结果在特定的空间尺度上才有意义[10].点格局分析方法[11,12]以植物分布的空间坐标为基础,可以分析各种尺度下的种群格局和种间关系,目前,多尺度研究主要集中在热带森林.亚热带森林生物多样性丰富,但受大面积物种空间数据获得的限制,关于多尺度格局分析方面的研究较少[13~18].现代生态学表明,群落物种共存不但与局域尺度的生态学过程有关,而且受大尺度上生态学过程影响物种的空间格局有很强的尺度依赖性[19].伏牛山脉是中国北亚热带和暖温带的气候分区线,属暖温带落叶阔叶林向北亚热带常绿落叶混交林的过渡区,也是油松(PinustabulaeformisCarr.)分布的南界.油松是中国特有的针叶树种,油松适应性强、根系发达,材质优良并具有良好的水土保持功能,历来受到研究者的重视,目前对其生物学和生态学特性、地理分布、生产力等都进行过较深入的研究[20~24].油松在伏牛山呈纯林分布或和麻栎混生形成混交林,选取此处的油松纯林和油松-麻栎混交林进行空间格局研究,能够更好地反应出种群与环境及其周围个体之间相互影响和作用,为进一步阐明群落结构、协调种间关系提供理论依据,对森林生态系统的维护和发展也具有重要意义.
1 研究区域概况
伏牛山脉位于河南省西部,是秦岭东西方向上的余脉,山体呈西北—东南走向,西北与熊耳山相连,东南抵南阳盆地东北边缘.地处暖温带向北亚热带过渡地带,植被属暖温带落叶阔叶林向亚热带常绿阔叶林的过渡型.气温暖和,四季分明,无霜期为7个月,1月平均温度为-2~2 ℃,7月平均温度为24~27 ℃,年平均温度为12.2~14.6 ℃.年均降水量885.6 mm,年均蒸发量991.6 mm,年均相对湿度68%.研究区域位于伏牛山余脉,这一区域油松以纯林和混交林2种形式存在.
2 材料与方法
2.1样地调查
本试验选取伏牛山脉油松纯林和油松-麻栎混交林样地作为研究对象.在海拔1 300 m处油松(油松纯林,油松-麻栎混交林)自然状态保存良好的地段,同一海拔设置3个30 m×30 m的样方,样方坡度方向和水平方向均为30 m.对样方内的优势种群油松和麻栎每木调查其树高、胸径、冠幅、坐标.样地基本概况见表1.
2.2径级结构
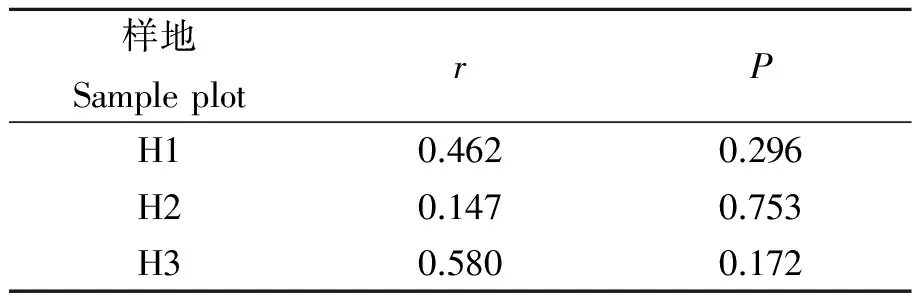
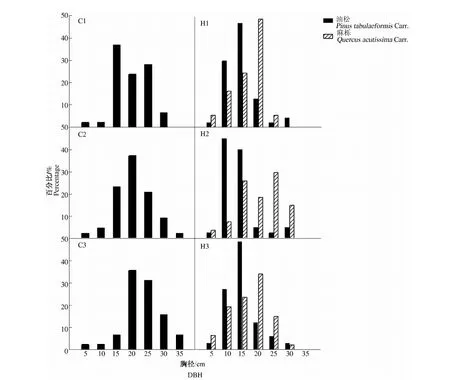
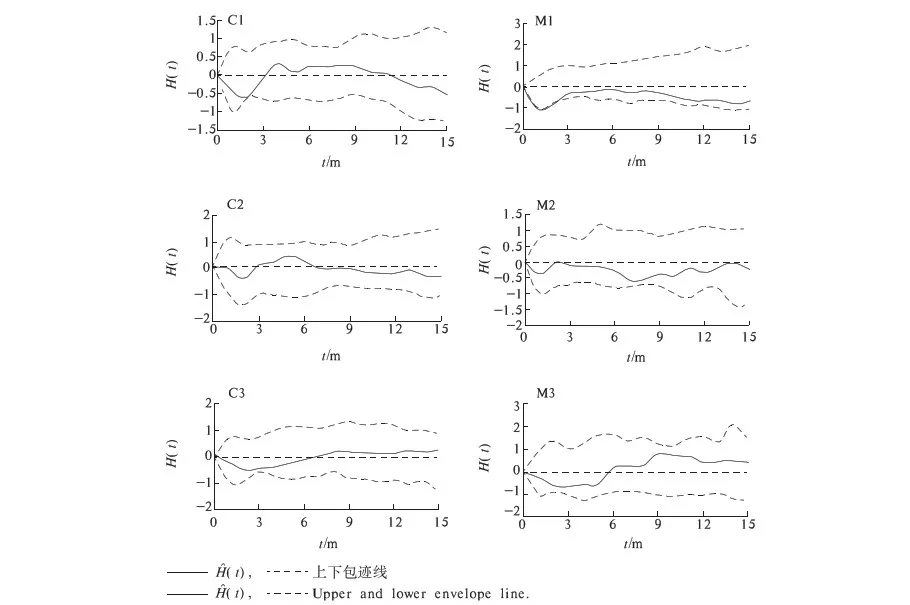
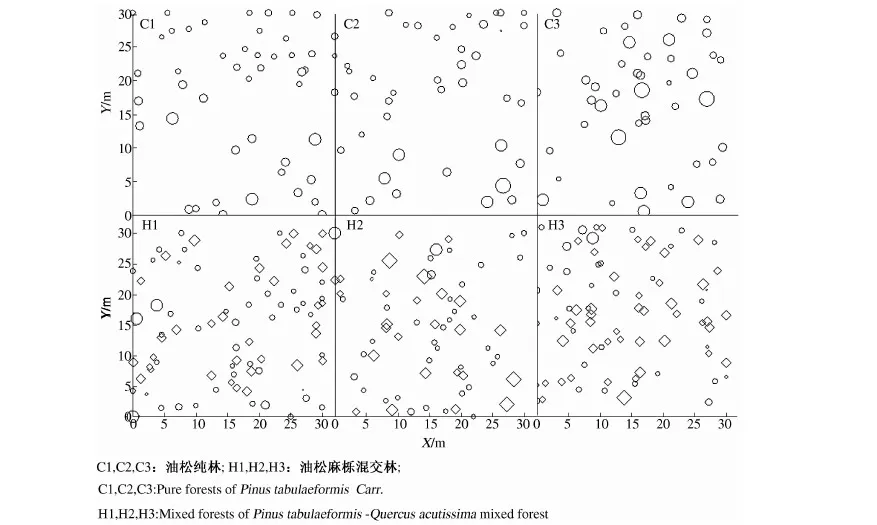
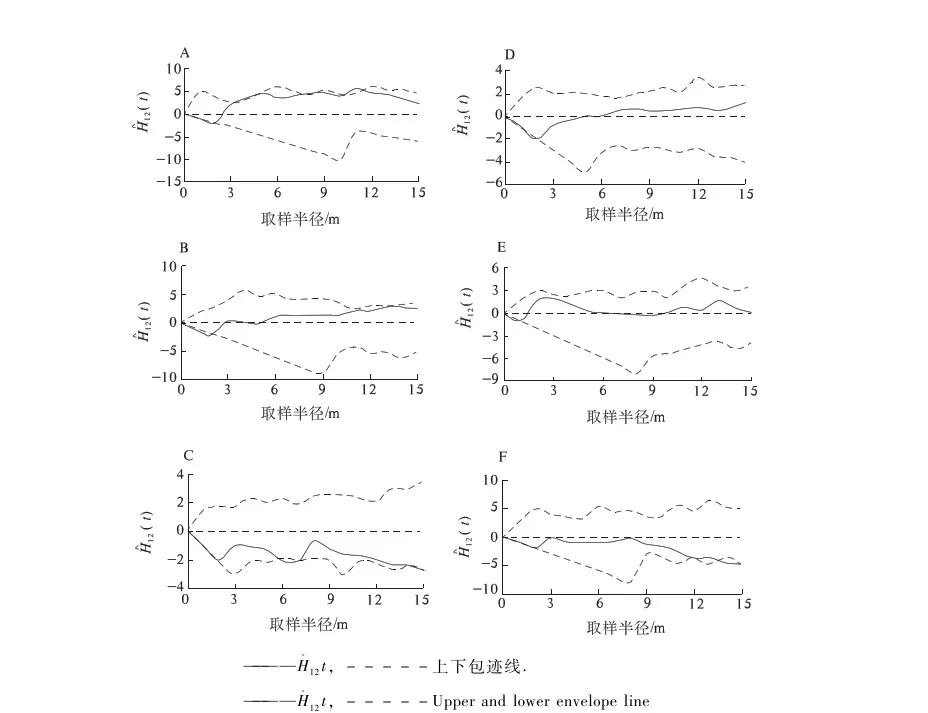
由于相同环境下同一树种龄级和径级对环境的反应规律具有一致性,所以研究采用树木径级代替龄级[25].以5 cm为1个径级,将油松、麻栎划分为7个径级: Ⅰ级:≦10 cm;Ⅱ级:10 cm 表1 样地概况 2.3种群空间格局分析 分析种群空间分布格局的2种主要方法是最近邻体分析和Ripley K函数[26].近年来,由于Ripley所提出的K(d)函数通过描述不同空间尺度下种群的分布格局,可以更好地反映种群的结构特点,已成为分析种群空间分布格局最常用的方法[27~31]和最广泛的点格局分析方法[32,33].点格局分析法将每个个体视为二维空间的1个点,以全部个体组成的二维点图为基础进行格局分析[34,35].Ripley K函数的最大研究尺度为矩形样地最小边长的1/2,研究为15 m.设n是样地内植物个体数,A是样地面积,Ripley K函数定义为[10,28,36~38]: (1) 式中:uij为个体i和j之间的距离,t是以目标树做圆点的取样圆半径,当uij≤t时,It(uij)=1;当uij>t时,It(uij)=0;Wij为边界效应校正系数,是指以个体i为圆心,以uij为半径的圆在样地中的面积比例. 对(1)式进行转换得(2)式,可以更好地观察物种的分布[32].用(2)式进行油松空间点格局分析. (2) (3) 式中:n1,n2是2种格局在样地内的个体数,其他符号同公式(1).基于同样的原因,对式(3)进行转换得(4)式,以对油松在不同年龄阶段之间空间格局的关联状况进行研究. (4) 3.1种群径级结构 3.1.1 油松纯林径级结构 纯林中油松作为主要乔木,它的发展除了受周围气候因素等的影响,还会受到与周围个体之间产生的种内竞争.所以依据油松径级的分布如图1(C1,C2,C3),可对种内关系分析.由图1可知,纯林中油松种群个体胸径主要在15~30 cm之间,胸径在15 cm以下及35 cm以上的个体较少.从纯林样地的径级分布来看,目前种群整体属于稳定型.这主要是油松个体自身的生态习性及环境因素造成的. 3.1.2 油松-麻栎混交林径级结构 油松和麻栎的径级分布如图1(H1,H2,H3)所示,由图可知,H1,H2,H3中油松个体胸径主要集中于10~20 cm之间,麻栎种群个体胸径主要集中在10~25 cm.某些范围内油松和麻栎的径级分布变化趋势是一致的,如H1的胸径在5~15 cm的个体、H2中5~10 cm和10~15 cm之间的个体,H3中胸径在5~15 cm和20~30 cm的个体. 3.2.混交林油松与麻栎相关性 根据油松和麻栎的径级分布情况,对油松和麻栎的相关性进行分析,结果如表2所示.从混交林中油松和麻栎的相关性分析可知,油松和麻栎径向生长呈正相关且相关性很高.差异显著性来看并不十分显著,可能是由于取样面积太小造成的,因为种群的分布状态会随空间尺度的增大有一定的改变. 表2 伏牛山混交林油松-麻栎径级分布的相关性分析 3.3种群格局分布及关联 3.3.1 不同群落油松的空间分布 根据Ripley K函数对伏牛山混交林和纯林油松数据进行分析,得出油松空间分布格局函数图(图2).由图2可知,纯林和混交林中6块样地均属随机分布,但不同样地之间又有差异. C1,C2,C3:油松纯林样地;H1,H2,H3:油松-麻栎混交林样地. 3个纯林样地中,在1~2 m油松由随机分布逐渐趋于均匀分布;之后随尺度增大,C1,C2油松种群逐渐变为随机,在4~5 m趋于聚集分布,之后随尺度进一步变大到15 m时,又趋于均匀分布;C3变化稍微缓慢,但趋势一致. 混交林H1中,在1~2 m油松几近于均匀分布,但随尺度变大,随机分布更明显,之后到11 m尺度时又再次趋于均匀分布,到15 m趋势仍有变化趋势.H2中在1~2 m和7~8 m最接近于均匀分布,之后在8~14 m略有波动的随机分布更明显,在14 m时转而趋向于均匀分布.H3中1~5 m更多的趋向于均匀分布,之后随机分布更明显,在14~15 m也开始有随机分布减弱、均匀分布增强的趋势. 3.3.2 纯林中油松不同径级的空间分布 本试验所用的点分布图是以植株所在样地中的位置来表示其空间位置的1种散点图.它是把植物个体当作空间中的点、以符号大小代表径级大小,直观地表达种群在样地内的空间分布状态的一种表现方式.油松纯林样地中,油松不同径级个体的空间分布情况如图3所示. 从图3可以看出,纯林中油松在样地内广泛分布,但Ⅰ,Ⅱ,Ⅲ径级个体有小范围的聚集分布现象.在样地内龄级较大个体附近有林窗形成.混交林中整体呈均匀分布,其中油松种群在样地内分布广泛,主要呈均匀分布.3个纯林(C1,C2,C3)整体来看均是随机分布,但从不同径级来分析可知,C1中胸径在15~20 cm的个体,呈聚集分布;其他径级个体呈随机分布格局.C2,C3中胸径在15~20 cm的个体呈聚集;在20~30 cm的个体为随机但近于均匀分布;其余各径级呈随机分布. 图2 油松不同种群空间分布格局 图3 伏牛山油松纯林、混交林不同径级的空间分布 混交林样地H1,H2,H3中油松和麻栎的分布格局如图2所示,3个样地中径级为10~15 cm的油松个体均为聚集分布,此径级范围的麻栎呈随机分布;25~30 cm径级间的油松呈随机分布,而此径级范围的麻栎呈聚集分布. 上下包迹线. 3.3.3 不同群落中油松种群不同径级间的空间关联 纯林中由图4-A可知,Ⅰ与Ⅱ径级在<2 m尺度上为趋于负关联,之后随距离的扩大,关联性不明显,在11~12 m尺度上是趋于正相关;Ⅰ与Ⅲ径级在<2 m的尺度上趋于负关联,11~15 m尺度上趋于正关联,其他尺度上关联性不明显(图4-B).第Ⅲ与Ⅳ、Ⅲ与Ⅴ径级在<2 m尺度上为负相关,其他尺度上相关性不明显(图4-C,图4-D).而纯林其他各径级两两之间关联性均不显著. 混交林中由图4-E知Ⅰ与Ⅱ径级在<1 m的尺度上为负关联,其他尺度关联性较弱;Ⅱ与Ⅳ径级在<2 m、14~15 m尺度上趋于负关联,其他尺度上关联性不明显(图4-F).混交林中其他各径级之间关联性不显著. 植物个体数按径级分布规律不论天然林或人工林,林木个体大小的数量分布均能反映出林分的生长状况和林木之间的竞争关系[39],能够反映种群结构与动态特征[40,41],是林分结构特征的重要指标. 1)从径级结构来看,纯林中油松个体胸径主要分布于15~30 cm,胸径在15 cm以下及35 cm以上的个体较少,即幼苗更新不良,以中龄树为主,群落短时间内稳定.这主要是油松个体自身的生态习性及环境因素造成的.混交林中油松径级主要分布在10~15 cm,以幼苗为主,整个油松种群径级分布可以看出老龄个体少,种群呈增长趋势.而麻栎也是以中龄个体居多,且从径级分布图可以看出,麻栎种群属于稳定型.由此可见,混交林中油松和麻栎之间虽然存在竞争,但并非抑制关系. 2)不同种群的油松空间格局研究 纯林在4~5 m趋于聚集分布;1~2 m及之后随尺度进一步变大均趋于均匀分布.混交林3个样地整体都属于随机分布,但与纯林不同的是,混交林中油松更多的随尺度增大格局由随机变为均匀.由于外界气候、土壤等因素纯林和混交林基本相似,所以混交林中油松的分布主要受麻栎种群的影响.物种在不同尺度下会表现出不同的空间分布特征,环境的异质性和聚集斑块的大小都能影响物种在不同尺度下的空间分布[42].随机性随尺度增加而减小,最终趋于均匀分布,这在一定程度上说明了种群的发展趋势,种内种间竞争使得空间分布由聚集趋于均匀,随机分布只是中间过程[43]. 3)油松种群不同径级的空间分布及关联分析 纯林中油松第Ⅲ径级个体呈聚集分布,其他各径级呈随机分布,表明纯林油松第Ⅲ径级个体种内呈正关联,种内竞争不明显.Ⅰ和Ⅲ径级之间在<2 m的范围内为负关联,11~12 m为正关联.Ⅲ和Ⅳ、Ⅲ和Ⅴ径级之间在<2 m的面积呈负关联,其他尺度上相关性不明显.表明在小尺度范围内第Ⅲ径级个体与其他径级个体种内竞争明显,随着尺度的增大,这种竞争逐渐减弱,甚至出现正关联的生态关系.Ⅰ和Ⅲ径级个体之间的关联变化可能是幼树与稍大个体对阳光等环境因子的需求不同造成的,而Ⅲ和Ⅳ、Ⅲ和Ⅴ径级之间可能是因为随着植物个体径级的增大,其对空间和环境资源的需求会随之加剧,导致种间和种内个体相互作用增强而造成自疏或它疏作用,这可能导致种群随径级的增加而聚集强度逐渐减弱的主要原因[10]. 混交林Ⅰ与Ⅱ径级个体的空间分布可以看出1~2 m范围油松幼龄个体之间的关联性较弱,而纯林中Ⅰ与Ⅱ径级个体的空间分布在<2 m均趋于负关联,11~12 m呈正关联,这可能是混交林中麻栎种群的影响.在1~2 m麻栎在一定程度上减弱了幼龄个体间的种内竞争.混交林中油松第Ⅱ与Ⅳ径级在14~15 m呈负相关,而纯林中第Ⅱ与Ⅳ径级在5~15 m呈负相关.这表明相同环境因素下,第Ⅱ与Ⅳ径级之间的关联性差异主要由混交林中麻栎种群的存在造成的.麻栎的存在减弱了一定范围内油松种群内部的负向作用.这也与种群径级结构分析的结果相一致. 虽然麻栎和油松都是喜光树种,但在幼苗期都能耐一定的庇荫.麻栎能在郁闭度较大的林冠下生长1~2年,而油松耐庇荫时间更长[44].油松和麻栎之间的竞争由于需要光照等时间段的差异,并未显现激烈的空间和环境资源的争夺.所以两者混交,并没有影油松种群的更新,反而缓和了种间竞争. 由此可见,伏牛山麻栎的存在促进了油松种群的发展,混交林比纯林的存在方式更有利于森林的发展,这与翟明普等[45]人关于油松麻栎混交林的研究结论一致.油松麻栎混交林群落稳定性比油松纯林更高,这与生物多样性能够影响生态系统的稳定性[46]、稳定性随物种多样性增加而提高[47]的结论相一致. 参考文献: [1]彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1996. [2]马华文,肖 锐,卢 军.凉水实验林场不同类型红松林空间分布格局[J].东北林业大学学报,2008,36(7):87-89. [3]藏润国.林隙更新动态研究进展[J].生态学杂志,1998,17(2):50-59. [4]王伯荪.植物群落学[M].北京:高等教育出版社,1987. [5]周灿芳.植物群落动态研究进展[J].生态科学,2000,19(6):53-59. [6]禄树晖,潘朝晖.藏东南高山松种群分布格局[J].东北林业大学学报,2008,36(11):22-24. [7]MAOABE T,NISHIMURA N,MIURA M,et al.Population structure and spatial patterns for trees in a temperate old-growth evergreen broad-leaved forest in Japan[J].Plant Ecology,2000,151:18l-197. [8]NISHIMURA N,ILARA T,MIURA M,et al.Tree competition and species coexistence in a warm-temperate old-growth evergreen broad-leaved forest in Japan[J].Plant Ecology,2002,164:235-248. [9]HE F,LEGENDRE P,LAFRANKIE J V.Distribution pattems of tree species in a Malaysian tropical rain forest[J].Journal of Vegetation Science,1997, 8:105-114. [10] 李 立,陈建华,任海保.古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J].植物生态学报, 2010,34(3):241-252. [11] RIPLEY B D.Modelling spatial pattern[J].Journal of the Royal Statistical Society,1977,39:172-212. [12] DIGGLE P J.Statistical analysis of spatial point patterns[M].New York:Academic Press,1983. [13] CONDIT R,ASHTON P S,BAKER P,et al.Spatial patterns in the distribution of tropical tree species[J].Science,2000,288:14l4-1418. [14] PLOTKIN JB,CHAVE J,ASHTON P S.Cluster analysis of spatial patterms in Malaysian tree species[J].The American Naturalist,2002,160:629-644. [15] WOODALL C W,GRAHAM J M.A technique for conducting point pattern analysis of cluster plot stem-maps[J].Forest Ecology and Management,2004,198:31-37. [16] YOUNG BLOOD A,MAX T,COE K.Stand structure in east side old-growth ponderosa pine forests of oregon and northern Califomia[J].Forest Ecology and Management,2004,199:191-217. [17] 王本洋,余世孝.种群分布格局的多尺度分析[J].植物生态学报,2005,29(2):235-241. [18] 汤孟平,周国模,施拥军,等.天目山常绿阔叶林优势种群及其空间分布格局[J].植物生态学报.2005,30(5):743-752. [19] HUBBELL S P.Neutral theory and the evolution of ecological equivalence[J].Ecology,2006, 87:1387-1398. [20] 徐化成.油松[M].北京:中国林业出版社,1993. [21] 徐化成,郭广荣,冯 林,等.油松天然林的生长与地理、气候因素的关系[J].北京林学院学报,1981(04):9-13. [22] 徐化成.油松地理变异和种源选择[M].北京:中国林业出版社,1992. [23] 中国植被编辑委员会.中国植被[M].北京:科学出版社,1983. [24] 马钦彦.油松分布区气候区划[J].北京林业大学学报,1989,11(2):1-9. [25] FROST I.RYDIN H.Spatial pattern and size distribution of the animal-dispersed Quercus rubur in two spruce dominated forests[J].Ecoscience,2000,7:38-44. [26] MOEUR M.Characterizing spatial patterns of trees using stem-mapped data[J].Forest Science, 1993,39(4):756-775. [27] SZWAGRZYK J.Small-scale spatial patterns of trees in a mixedPinussylvestris-Fagussylvaticaforest[J].Forest ecology and management,1992,51(4):301-315. [28] CRESSIE N A.Statistics for spatial data[M].New York:Wiley,1993:900. [29] BATISTA J, MAGUIRE D.Modeling the spatical structure of tropical forests[J].Forest Ecology,1998,110:293-314. [30] HANUS M,HANN D W, MARSHALL D D.Reconstructing the spatial pattern of trees from routine stand examination measurements[J].Forest Science,1998,44(1):125-133. [31] WELLS M L,GETIS A.The spatial characteristics of stand structure in Pinus torreyana[J].Plant Ecology,1999,143(2):153-170. [32] 张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349. [33] 张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35-44. [34] RIPLEY B D.Modelling spatial pattern[J].Journal of the Royal Statistical Society:Series B,1977,39(2):172-212. [35] DIGGLE P J.Statistical analysis of spatial point patterns[M].New York:Academic Press.1983. [36] MOEUR M.Characterizing spatial patterns of trees using stem-mapped data[J].Forest Science,1993,39(4):756-775. [37] BATISTA J , MAGUIRE D.Modelling the spatial structure of tropical forests[J].Forest Ecology,1998,110:293-314. [38] HANUS M,HANN D M, MARSHALL D D.Reconstructing the spatial pattern of trees from routine stand examination measurements[J].Forest Science,1998,44(1):125-133. [39] 陈光彩,郝士成,李 怡,等.麻池背油松天然林林分生长结构的研究[J].山西林业科技.2004,12(4):10-13. [40] GRAU H R,ARTURI M F,BROWEN A D,et al.Floristic and structure pattern along a chronosequence of secondary forest succession in Argentinean subtropical montane forests[J].Forest Ecology and Management,1997,95(2):161-171. [41] 范繁荣,潘标志,马祥庆,等.白桂木的种群结构和空间分布格局研究[J].林业科学研究,2008,21(2):176-181. [42] RIGINOS C,MILTON S J,WIEGAND T.Context-dependent in-teractions between adult shrubs and seedlings in asemi-aridshrubland[J].Journal of Vegetation Science,2005,16:331-340. [43] 王本洋,余世孝.种群分布格局的多尺度分析[J].植物生态学报,2005,29(2):235-241. [44] 河南森林编辑委员会.河南森林[M].北京:中国林业出版社,2000:92-190. [45] 翟明普,谭维军,谭占民.临朐地区油松麻栎混交林的研究[J].北京林业大学学报,1990,12(10):64-70. [46] 贾 力,赵 娜.生物多样性和生态系统功能的研究历史与现状[J].内蒙古科技与经济,2012,16(10):46-50. [47] 岳天祥.生物多样性研究及其问题[J].生态学报.2001,21(3):462-467.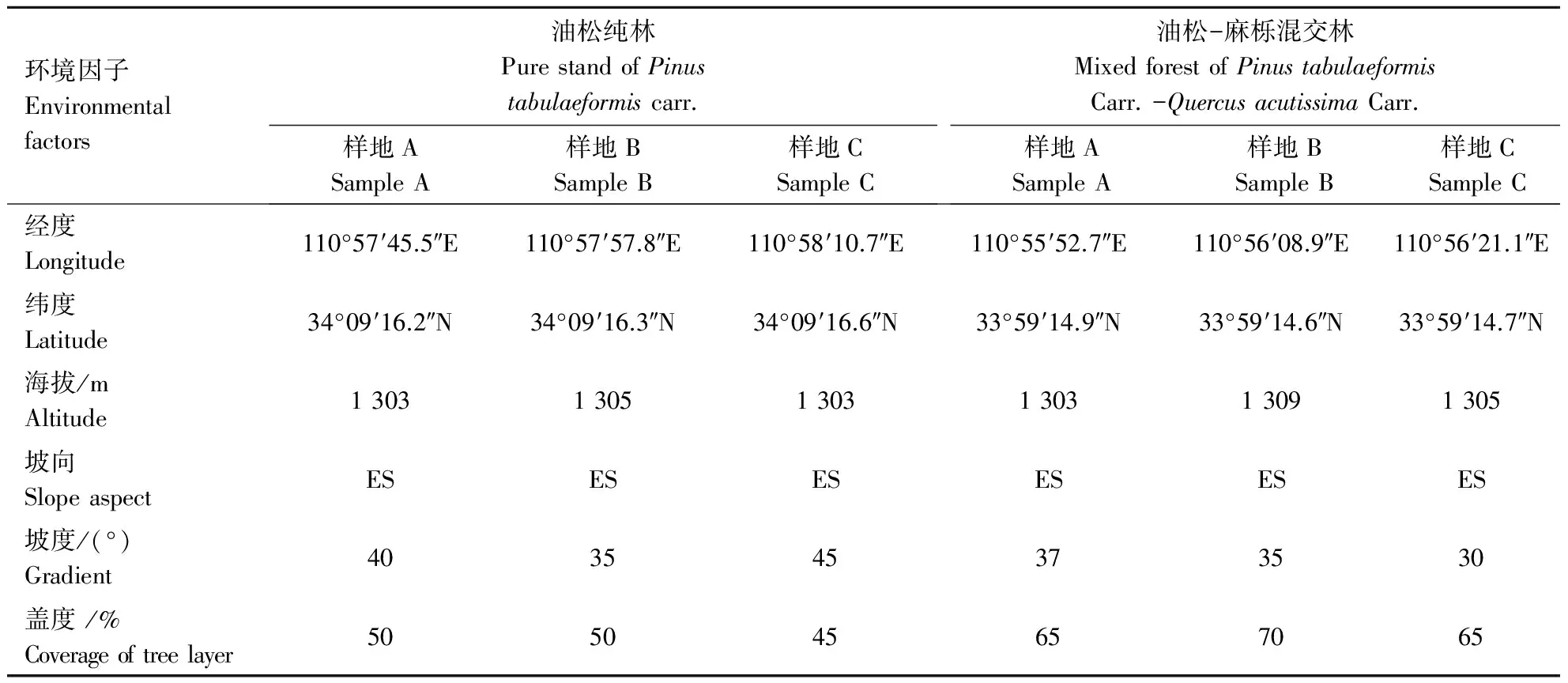
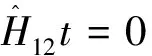
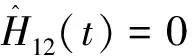
3 结果与分析
4 结论与讨论
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
森林工程(2018年4期)2018-08-04 03:23:10
方圆(2018年23期)2018-01-07 09:06:18
广西林业科学(2016年4期)2016-03-16 05:44:58
西北植物学报(2015年9期)2015-07-05 07:03:42
