
湖北钉螺被目平外睾吸虫与日本血吸虫不同间隔时间感染后分泌物的检测与分析
2014-04-02 02:39:40唐崇惕黄帅钦彭午弦卢明科彭文峰
中国人兽共患病学报 2014年11期
唐崇惕,黄帅钦,彭午弦,卢明科,彭文峰,陈 东
湖北钉螺(Oncomelaniahupensis)被外睾吸虫(Exorchisspp.)感染后很快产生大量血淋巴细胞和分泌物并能杀害再进入其体内的日本血吸虫(Schistosomajaponicum)幼虫[1,3,11]。外睾吸虫和日本血吸虫先后双重感染钉螺间隔时间愈长,血吸虫被杀害的效果愈强,但钉螺血淋巴细胞的数目却是逐渐减少[2,4],钉螺体内还有什么东西在起作用?近年许多学者在南美洲和非洲大力开展曼氏血吸虫(Shistosomamansoni)与其中间宿主水生双脐螺(Biomphalariaspp.)的相互关系问题的研究[5]。探讨感染曼氏血吸虫的双脐螺体內血浆中的蛋白质组学(proteomics),有关它们的多态性、多态粘蛋白(polymorphic mucins)[8-9],与血纤维源有关的蛋白质(fibrinogen-related proteins) 等的特性、免疫应答基因家族情况[13-14],有关软体动物防御细胞中激活酶(kinase)多方面作用情况[10]。有学者研究棘口吸虫(Echinostomacaproni)和曼氏血吸虫幼虫在双脐螺中的排泄和分泌的蛋白质组(proteome)[7],棘口吸虫寄生的双脐螺血浆及血淋巴细的不同蛋白质的特性[6,12]。有关曼氏血吸虫与双脐螺相关的螺体血浆中物质的研究报告无数,可见其中复杂程度。笔者从外睾吸虫和日本血吸虫双重感染的钉螺体上观察到有复杂的分泌物增多,它们在不同间隔时间感染的钉螺体内外表现情况有所不同,兹将观察结果简单介绍于下。
1 材料与方法
用从湖南西洞庭湖鲶鱼肠管收集的目平外睾吸虫(ExorchismupingensisJiang,2011)的虫卵拌以少量面粉饲食从西洞庭湖采集的湖北钉螺,外睾吸虫感染后的钉螺分为5组,分别在感染后21 d、37 d、55 d、70 d、85 d,每粒钉螺与实验室日本血吸虫阳性小白鼠肝脏血吸虫虫卵孵化的毛蚴(40~90个)接触;各组钉螺均在感染血吸虫毛蚴后4~82 d之间不同时间,用10%福尔马林溶液固定。钉螺单独感染外睾吸虫后20 d,也用10%福尔马林溶液固定。所有实验钉螺均经石蜡连续切片及用苏木精与洋红染色制片。显微镜油镜检查各切片所有断面,比较观察各组无其它吸虫天然感染的钉螺其体内外分泌物的数量及它们的结构情况,并数码相机显微照相储存于电脑。
2 结 果
在不同间隔先后双重感染目平外睾吸虫和日本血吸虫的钉螺体内分泌物详细情况的本实验观察中,同样见到间隔时间长短与螺体杀害血吸虫幼虫的效力成正比,而与在螺体及异常血吸虫幼虫体内的3种血淋巴细胞数量成反比的现象。但钉螺体内外的分泌物密度没有减少,有增多并且更加复杂化。情况分述如下。
2.1单独感染外睾吸虫的钉螺体表分泌物结构 作为吸虫类的中间宿主贝类,其体表都有一层分泌物。切片观察单独感染日本血吸虫的钉螺,其体表的分泌物仅一层粘液性薄膜,而被外睾吸虫感染的钉螺其体表分泌物增厚[11]。本实验观察单独感染外睾吸虫后21 d的钉螺,其体表也有较厚的分泌物(图1),在油镜下可见它们由排列不规则、大小和形状各异、透明或不透明的晶体状结构所密布,其中及边缘有些微粘液样物质(图2~3)。
2.2钉螺体内分泌物 在钉螺体中部、肉足和鳃的下方有一大团副腺组织(图6),许多副腺细胞游离在螺体各组织中。在所有双重感染外睾和血吸虫的钉螺,副腺细胞亦可见于阔张的血管中[3]。在不同间隔双重感染外睾和血吸虫的钉螺,所有被击毁的血吸虫幼虫残体内外都可见到这些细胞(图7、9、22等)。
单独感染外睾吸虫的钉螺体内血淋巴细胞和分泌物也比单独感染日本血吸虫的多,明显的金黄色颗粒团状分泌物(图4~5)出现在钉螺体内一些区域。双重感染外睾和血吸虫的钉螺,双重感染间隔时间加长体内分泌物有增无减,分泌物颗粒和血淋巴细胞经常一起出现在被击毁的异常血吸虫幼虫体内外(图8、10、13、17、19等)。有时它们会呈褐色团状大量地分布在螺体组织及异常血吸虫体内,以间隔55 d双重感染后56 d(即外睾吸虫感染后111 d,血吸虫残体为6 d)的照片(图14)为例,大团褐色物占据血吸虫残体大部,虫体所在血腔外的螺体组织中密布许多小褐色团。如此情况时常可见。
前此在外睾和血吸虫双重感染的钉螺体内,已见到在金黄色分泌物颗粒每团边缘都有一个比小血淋巴细胞核更小的细胞核,其直径只有1.9~2.6 μm[3]。此次用油镜观察,仍然是此情况,而且在核的外围都可见到有一圈白色外围,它应该是其细胞质部位。说明它们确有细胞的结构,双重感染时间愈长此极小细胞愈明显(图8、11~12、16、18~20)。
2.3钉螺体内分泌物颗粒在异常血吸虫幼虫体内的异样产物 在外睾和血吸虫双重感染的钉螺体内,尚有奇怪现象:在很早期被击毁的异常血吸虫幼虫体内会出现一个大红色球团,它整个表面布满颗粒小点,球团边缘围绕甚多与上述分泌物颗粒上极小细胞同样大小的胞核和细胞样的结构(图8~10、17)。双重感染间隔时间愈长的钉螺,异常血吸虫幼虫体内此红球团数常会增到2~4个,而且历久不退,如间隔70 d后的6 d和25 d异常血吸虫幼虫的样品(图15、21)。
2.4外睾吸虫和日本血吸虫双重感染的钉螺体表分泌物结构 外睾吸虫和日本血吸虫双重感染的钉螺,它们体表分泌物均有增无减,其厚度可达单独感染外睾吸虫钉螺体表分泌物厚度数倍(图23)。两吸虫感染间隔时间增长,其体表分泌物不仅增多,而且其内容更加复杂化。其中晶体状物体逐渐呈有规则地成条成片地排列(图26~27),在许多晶体样物质之间除了有更多粘液样物体之外还有许多原本是在钉螺体内的物质。两吸虫间隔85 d双重感染,在血吸虫感染后5 d~25 d的钉螺,它们体中都只能查到已完全解体的血细虫幼虫残骸。在血吸虫感染后5 d的钉螺体表分泌物(图23)中在许多成片条状晶体物质之间除有许多粘液状物质之外还夹杂着许多原来在钉螺体内的血淋巴细胞和含极小细胞的分泌物颗粒(图24~28)。
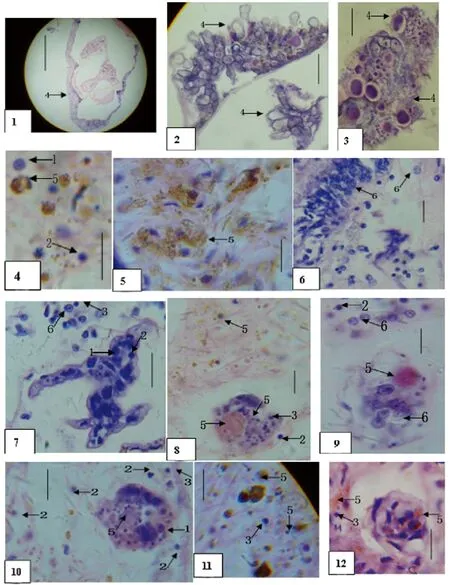

图版箭矢数字说明(Arrownumbersillustrate):
1=大血淋巴细胞(bighemo-lymphocyte);2=中血淋巴细胞(mediatebighemo-lymphocyte);
3=小血淋巴细胞(smallhemo-lymphocyte);4=螺体外分泌物(snailbodyoutersecretion);
5=螺体内分泌物及小核(snailinnersecretionandverysmallnucleus);
6=螺副腺细胞(snailaccessoryglandcell)。
图说明(Em=Exorchismupingensis;Oh=Oncomelaniahupensis;Sj=Schistosomajaponicum)
图1~6被Em感染的Oh其分泌物及副腺细胞
Figs.1-6SecretionsandaccessoryglandcellsofOhsnailsinfectedbyEmtrematodes
图7间隔21d双重感染Em和Sj的Oh示其体内分泌物和血淋巴细胞,及其对Sj幼虫的反应
Fig.7OhsnailduallyinfectedbyEmandSjfor21dinterval,showingthesnail’ssecretions,lymphocytesandtheirreactionstoSjlarvae
图8~11.间隔37d双重感染Em和Sj的Oh示其体内分泌物和血淋巴细胞,及其对Sj幼虫的反应
Figs.8-11OhsnailsduallyinfectedbyEmandSjfor37dinterval,showingthesnail’ssecretions,lymphocytesandtheirreactionstoSjlarvae
图12~14间隔55d双重感染Em和Sj的Oh示其体内分泌物和血淋巴细胞,及其对Sj幼虫的反应
Figs.12-14OhsnailsduallyinfectedbyEmandSjfor55dinterval,showingthesnail’ssecretions,lymphocytesandtheirreactionstoSjlarvae
图15~22间隔70d双重感染Em和Sj的Oh示其体内分泌物和血淋巴细胞,及其对Sj幼虫的反应
Figs.15-22OhsnailsduallyinfectedbyEmandSjfor70dinterval,showingthesnail’ssecretions,lymphocytesandtheirreactionstoSjlarvae
图23~28.间隔85d双重感染Em和Sj的Oh示其体内分泌物和血淋巴细胞,及其对Sj幼虫的反应
Figs.23-28OhsnailsduallyinfectedbyEmandSjfor85dinterval,showingthesnail’ssecretions,lymphocytesandtheirreactionstoSjlarvae
图1Oh体外分泌物 (Scale bar=0.15 mm)
Fig.1 Outer secretions ofOhsnail
图2Oh体外分泌物 (Scale bar=0.03 mm)
Fig.2 Outer secretions ofOhsnail
图3Oh体外分泌物(Scale bar=0.023 mm)
Fig.3 Outer secretions ofOhsnail
图4Oh体内分泌物(Scale bar=0.034 mm)
Fig.4 Inner secretions ofOhsnail
图5Oh体内分泌物(Scale bar=0.030 mm)
Fig.5 Inner secretions ofOhsnail
图6Oh体内副腺细胞(Scale bar=0.022 mm)
Fig.6 Accessory gland cells ofOhsnail
图7Em感染后37 dOh体中3种血淋巴细胞在16 d异常Sj幼虫体内外(Scale bar=0.034 mm)
Fig.7 Three species hemo-lymphocytes in 16-d-old abnormalSjlarva and tissue of snail post-infected byEmfor 37 d
图8 含极小细胞核的分泌颗粒散布在感染Em后42 d的Oh体组织中,具分泌物颗粒的球形红团在5d异常Sj幼虫体内(Scale bar=0.022 mm)
Fig.8 The secretion granules with very small cell nucleus spreading in the tissue ofOhpost-infected byEmfor 42 d and a spherical red mass with secretions granules in 5-d-old abnormalSjlarva
图9Em感染后42 d的Oh示其体中5d异常Sj幼虫含副腺细胞及具分泌物颗粒的红团(Scale bar=0.027 mm)
Fig.9 Body ofOhpost-infected byEmfor 42 d showing the 5-d-old abnormalSjlarva containing the accessory gland cells and a spherical red mass with secretions granules
图10Em感染后42 d的Oh示其体中血淋巴细胞及具红团的5d异常Sj幼虫 (Scale bar=0.030 mm)
Fig.10 Body ofOhpost-infected byEmfor 42 d showing the hemo-lymphocytes and 5-d-old abnormalSjlarva with hemo-cytes,red mass and secretion granules
图11 带有微小细胞核的分泌物颗粒团散布在Em感染后42 d的Oh组织中 (Scale bar=0.028 mm)
Fig.11 Secretion granules with small cell nucleus spreading in the tissue ofOhpost-infected byEmfor 42 d
图12Em感染后66 d 的Oh和11 d 异常Sj幼虫体中的具微小细胞核的分泌物颗粒(Scale bar=0.030 mm)
Fig.12 Secretion granules with small cell nucleus inOhpost-infected byEmfor 66 d and the body of 11-d-old abnormalSjlarva
图13Em感染后79 d的Oh示24 d 异常Sj幼虫残骸内的血淋巴细胞和具微小细胞核的分泌物颗粒 (Scale bar=0.025 mm)
Fig.13Ohpost-infected byEmfor 79 d showing the hemo-lymphocytes and secretion granules with small cell nucleus in the 24-d-old abnormalSjlarva wreckage
图14 大量分泌物团在Em感染后111 d的Oh及56 d异常Sj幼虫残骸内 (Scale bar=0.033 mm)
Fig.14Much secretions masses inOhpost-infected byEmfor 111 d and the 56-d-old abnormalSjlarva wreckage
图15Em感染后76 d的Oh示6 d异常Sj幼虫残骸内的具分泌物颗粒2个红团(Scale bar=0.016 mm)
Fig.15Ohpost-infected byEmfor 76 d showing two red masses with secretion granules and small cell nucleuses in the 6-d-old abnormalSjlarva wreckage
图16 间隔70 d双重感染Em和Sj后6 d的Oh体中散布许多具微小细胞核的分泌颗粒(Scale bar=0.028 mm)
Fig.16 Many secretion granules with small cell nucleus in theOhat the 6 d after dual infections ofEmandSjwith 70 d intervals
图17Em感染后84 d的Oh示14 d异常Sj幼虫残骸内的红团和微小细胞核(Scale bar=0.027 mm)
Fig.17Ohpost-infected byEmfor 84 d showing a red mass with small cell nucleuses in the 14-d-old abnormalSjlarva wreckage
图18 间隔70 d双重感染Em和Sj后25 d的Oh体中的许多具微小细胞核的分泌颗粒 (Scale bar=0.027 mm)
Fig.18 Many secretion granules with small cell nucleus in theOhat the 25 d after dual infections ofEmandSjwith 70 d intervals
图19Em感染后84 d的Oh示25 d异常Sj幼虫残骸内外的具微小细胞核分泌物颗粒 (Scale bar=0.028 mm)
Fig.19Ohpost-infected byEmfor 84 d showing the secretion granules with small cell nucleus inside and outside of 25d old abnormalSjlarva wreckage
图20 间隔70 d双重感染Em和Sj后25 d的Oh体内许多具微小细胞核的分泌颗粒 (Scale bar=0.025 mm)
Fig.20Many secretion granules with small cell nucleus in theOhat the 25 d after dual infections ofEmandSjwith 70 d intervals
图21Em感染后95 d的Oh示2个具分泌物颗粒的红团在25 d 异常Sj幼虫残骸内 (Scale bar=0.027 mm)
Fig.21Ohpost-infected byEmfor 95 d showing two red masses with secretion granules in the 25-d-old abnormalSjlarva wreckage
图22Em感染后95 d的Oh示副腺细胞在25d异常Sj幼虫残骸内 (Scale bar=0.030 mm)
Fig.22Ohpost-infected byEmfor 95 d showing the accessory gland cells in the 25-d-old abnormalSjlarva wreckage
图23 间隔85 d双重感染Em和Sj后5 d的Oh示体外分泌物(Scale bar=0.15 mm)
Fig.23Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing its thick body secretion
图24 间隔85 d双重感染Em和Sj后5 d的Oh示体外分泌物中的体内血淋巴细胞(Scale bar=0.027 mm)
Fig.24Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing hemo-lymphocytes in the body outside secretion
图25 间隔85d双重感染Em和Sj后5 d的Oh示体外分泌物中的体内小血淋巴细胞和分泌物颗粒微小细胞核(Scale bar=0.027 mm)
Fig.25Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing small hemo-lymphocytes and secretion granules with small cell nucleus in the body outside secretion
图26 间隔85 d双重感染Em和Sj后5 d的Oh示体外分泌物中的结晶体和体内分泌物颗粒微小细胞核(Scale bar=0.019 mm)
Fig.26Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing the opaque crystal structure of body secretion and inner secretion granules with small cell nucleus in the body outside secretion
图27 间隔85 d双重感染Em和Sj后5 d的Oh示体外分泌物中的结晶体和体内分泌物颗粒微小细胞核(Scale bar=0.019 mm)
Fig.27Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing the opaque crystal structure of body secretion and inner secretion granules with small cell nucleus in the body outside secretion
图28 间隔85 d双重感染Em和Sj后5 d的Oh的体外分泌物中的结晶体和体内分泌物颗粒微小细胞核(Scale bar=0.019 mm)
Fig.28Ohat the 5 d after dual infections ofEmandSjwith 85 d intervals showing the opaque crystal structure of body secretion and inner secretion granules with small cell nucleus in the body outside secretion
3 讨 论
实验证明外睾吸虫和日本血吸虫双重感染钉螺,其间隔时间愈长螺体杀害血吸虫幼虫的效力愈强,钉螺感染外睾吸虫的开始时会增生大量血淋巴细胞,但在感染后一个月左右螺体增生血淋巴细胞性能就开始逐渐衰退,时间稍久后其数量逐渐显著减少[2,4];外睾吸虫幼虫期在钉螺体内无性生殖期很长,感染后105 d还处于原始胚细胞大量增殖期,6~7个月才成熟[1]。本实验外睾吸虫阳性钉螺同样表现在其感染后37~85 d能逐渐地更有力地攻击后侵入的血吸虫幼虫,使其致命解体,是否与螺体内外睾吸虫胚细胞正在大量增生有关?螺体内分泌物逐渐大量增加是否也与其有关?但是为何螺体的血淋巴细胞却在逐渐减少?其中奥秘机理需要继续探究。
钉螺体内外的分泌物从那里产生?在外睾吸虫和日本血吸虫双重感染的钉螺其体外分泌物中呈晶体结构是何物质?体内金黄色分泌物颗粒有增无减,每个颗粒团都更明显出有细胞核和细胞质的极小细胞,这细胞和金黄色颗粒团是何关系?这些分泌物与血吸虫幼虫被击毁有何关系?其中机理亦需继续探究。
外睾吸虫和日本血吸虫双重感染的钉螺,其体中被杀害的血吸虫幼虫体内经常有球状红团,两吸虫双重感染间隔时间愈长这红团数会从1个增加到4个。这红团的产生与其外表分泌物颗粒及极小细胞核相仿的物体有何关系?与血吸虫幼虫的被杀有无关系?其中机理亦需继续探究。
应用无害的外睾吸虫作材料处理钉螺,所有实验都证明可以百分之百杀死再侵入的日本血吸虫幼虫,有关此生物控制的机制都需要从多方面,包括免疫学、蛋白质组学和基因组学等等,进行深入研究,有利于更好地应用。当时研究生郭跃和王逸难同学协助感染工作,在此致谢。
参考文献:
[1]Tang CT,Shu LM. Early larval stages ofExorchisovariolobularisin its molluscan hosts and the appearance of lymphatic cellulose reaction of host[J]. Acta Zoologica Sinica,2000,46(4): 457-463. (in Chinese)
唐崇惕,舒利民,外睾吸虫幼虫期的早期发育及贝类宿主淋巴细胞的反应[J].动物学报,2000,46(4):457-463.
[2]Tang CT,Lu MK,Guo Y,et al. Comparison among the bio-control effects on larvalSchistosomajaponicuminOncomelaniahupensiswith pre-infection by larvalExorchistrematodes at different intervals[J]. Chin J Zoonoses,2010,26(11): 989-994. (in Chinese)
唐崇惕,卢明科,郭 跃,等,日本血吸虫幼虫在先感染外睾吸虫后不同时间钉螺体内被生物控制效果的比较[J].中国人兽共患病学报,2010,26(11):989-994.
[3]Tang CT,Guo Y,Lu MK. et al. Reactions of snail secretions and lymphocytes toSchistosomajaponicumlarvae inOncomelaniahupensispre-infected withExorchistrematode[J]. Chin J Zoonoses,2012,28(2): 97-102. (in Chinese)
唐崇惕,郭跃,卢明科,等. 先感染外睾吸虫的钉螺其分泌物和血淋巴细胞对日本血吸虫幼虫的反应[J]. 中国人兽共患病学报,2012,28(2):97-102.
[4]Tang CT,Lu MK,Chen D. Comparison between the existence states of the hemo-lymphocytes toOncomelaniahupensissnails dually infected by larvalExorchismupingensisandSchistosomajaponicumat different intervals[J]. Chin J Zoonoses,2013,29(8): 735-742. (in Chinese)
唐崇惕,卢明科,陈东,目平外睾吸虫日本血吸虫不同间隔时间双重感染湖北钉螺螺体血淋巴细胞存在情况的比较[J]. 中国人兽共患病学报,2013,29( 8):735-742.
[5]Bayne CJ. Origins and evolutionary relationships between the innate and adaptive arms of immune systems[J]. Integr Comp Biol,2003,43: 293-299.
[6]Bouchut A,Sautiere PE,Coustau C,et al. Compatibility in theBiomphalariaglabrata/Echinostomacapronimodel: potential involvement of proteins from hemocytes revealed by a proteomic approach[J]. Acta Trop,2006,98(3): 234-246.
[7]Guillou F,Roger E,Mone Y,et al. Excretory-secretory proteome of larvalSchistosomamansoniandEchinostomacaproni,two parasites ofBiomphalariaglabrata[J]. Mol Biochem Parasitol,2007,155(1): 45-56.
[8]Roger E,Mitta G,Mone Y,et al. Molecular determinants of compatibity polymorphism in theBiomphalariaglabrata/Schistosomamansonimodel: new candidates identified by a global comparative proteomic approach[J]. Mol Biochem Parasitol,2008,157(2): 205-216.
[9]Roger E,Grunau C,Pierce RJ,et al. Controlled chaos of polymorphic mucins in a metazoan parasite (Schistosomamansoni) interacting with its invertebrate host (Biomphalariaglabrata)[J]. PLoS Negl Trop Dis,2008,2(11): e330.
[10]Plows LD,Cook RT,Davies AJ,et al. Integrin engagement modulates the phosphorylation of focal adhesion kinase,phagocytosis and cell spreading in molluscan defence cells[J]. Biochim Biophys Acta,2006,1763(8): 779-786.
[11]Tang CT,Lu MK,Chen D,et al. Development of larvalSchistosomajaponicumblocked inOncomelaniahupensisby pre-infection with larvalExorchissp[J]. J Parasitol,2009,95(6): 1321-1325.
[12]Vergote D,Bouchut A,Sautiere PE,et al. Characterisation of proteins differentially present in the plasma ofBiomphalariaglabratasusceptible or resistant toEchinostomacaproni[J]. Int J Parasitol,2005,35(2): 215-224.
[13]Zhang SM,Loker ES. Representation of an immune responsive gene family encoding fibrinogen-related proteins in the freshwater molluscBiomphalariaglabrata,an intermediate host forSchistosomamansoni[J]. Gene, 2004,341: 255-266.
[14]Zhang SM,Zeng Y,Loker ES. Expression profiling and binding properties of fibrinogen-related proteins (FREPs),plasma proteins from the schistosome-snail hostBiomphalariaglabrata[J]. Innate Immun,2008,14(3): 175-189.
猜你喜欢
热带病与寄生虫学(2022年4期)2022-09-20 03:30:16
热带病与寄生虫学(2022年6期)2022-02-11 01:04:32
畜牧兽医科技信息(2021年2期)2021-03-29 10:40:30
热带病与寄生虫学(2021年2期)2021-02-28 15:15:11
湖北大学学报(自然科学版)(2021年1期)2021-01-06 08:17:34
黑龙江八一农垦大学学报(2020年6期)2021-01-04 07:37:10
中国动物传染病学报(2020年5期)2020-10-17 06:56:46
中国感染与化疗杂志(2020年1期)2020-01-12 14:07:30
兽医导刊(2019年1期)2019-02-21 01:14:28
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:23
