
小陇山林区锐齿栎原始林群落的空间结构特征
2014-03-27 00:41郭小龙刘文桢张宋智石小龙李安民
西北农林科技大学学报(自然科学版) 2014年11期
郭小龙,刘文桢,张宋智,吕 寻,石小龙,李安民
(甘肃省小陇山林业实验局 林业科学研究所,甘肃 天水 741022)
林分的空间结构特征体现了树木在林地上的分布格局及其属性在空间上的排列方式,即林分内林木的大小、分布和树种混交等空间关系,是与林木空间位置有关的林分结构[1],可从林木个体在水平地面上分布的空间格局、大小分化度和树种隔离程度3个方面加以描述[2-4]。森林空间结构是森林动态变化过程中测度时点林分状态的高度概况,是对森林发展过程中各种因素,如更新方式、竞争、自然稀疏及自然干扰和人为干扰作用结果的综合反映[5],已经成为森林经营和分析中的一个重要方面[1-2],其中涉及单木之间空间关系的林分空间结构及其空间关系的描述和解释已成为森林结构研究的焦点[6-7],越来越受到人们的重视。近年来,惠刚盈等[2,4-5]和汤孟平[7]提出了基于参照树与其最近4株相邻木关系的林分空间结构量化分析方法,许多学者将其应用于不同林分类型的空间结构特征分析、多样性分析、采伐木选择、结构重建等方面,展示出了良好的应用前景。
小陇山林区是我国西北地区重要的天然林区,在涵养水源、保持水土、维护地区生态平衡、提高环境质量、保护生物多样性以及林业生产等方面发挥着不可替代的作用。以往对小陇山林区的研究,主要集中在锐齿栎次生林群落的生物量、群落数量特征、群落结构、种间关系、生物多样性、空间结构特征等方面[8-11],而对其原始林空间结构特征的研究尚未见报道。本研究运用基于参照树与其最近4株相邻木关系的林分空间结构的量化分析方法,对小陇山林区锐齿栎原始林的空间结构特征进行研究,旨在为小陇山林区锐齿栎次生林的经营提供借鉴。
1 研究区及样地概况
1.1 研究区概况
小陇山林区处于黄河、长江中上游地区的渭河、嘉陵江流域交汇处(104°22′~106°43′E、30°30′~34°49′N),区内年均气温7~12 ℃,极端最低气温 -32.2 ℃,极端最高气温38.3 ℃,年积温3 563.9 ℃;年日照时数2 098.7 h,年太阳幅射量490.8~568.6 kJ/cm2。年均降水量460~800 mm,50%~60%的降水集中在7-9月份。年均绝对蒸发量1 287.6~1 339.0 mm,湿润度0.36~0.45,相对湿度69%;无霜期185 d;海拔750~3 200 m。研究区为我国典型的南北气候和植被过渡带,区内植物分布在中国植物区系分区系统中属中国-日本植物亚区、华北植物地区、黄土高原植物亚地区,区系组成具有明显的温带属性,区系成分以华北成分为主,兼有华中、喜马拉雅与蒙新成分,植物种类繁多,是北亚热带向温带过渡的植物基因库,有高等植物225科、877属、2 753种,其中木本植物有780多种,内含乔木281种。区内海拔2 200 m以下主要是以锐齿栎(Quercusalienavar.acuteserrata.)和辽东栎(Quercusliaotungensis)为主的天然林;在栎林带内分布有华山松(Pinusarmandi)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana)、漆树 (Rhusverniciflus)、冬瓜杨(Populuspurdomii)、千金榆(Carpinuscordata.)、甘肃山楂(Crataeguskansuensis)、刺楸(Kalopanaxseptemlobus)等乔木树种,灌木有美丽胡枝子(Lespedezathunbergii)、光叶绣线菊(SpiraeajaponicaL.f. var.fortunei)、中华绣线菊(Spiraeachinensis)、胡颓子(Elaeagnuspungens)、华北绣线菊(Spiraeafritschiana)、连翘(Forsytiasuspense)、卫矛(Euonymusalatus)、山豆花(Lespedezatomentosa)等。
1.2 样地概况
研究样地布设于百花林场坪营林区91林班的14小班内,样地海拔1 857 m,东北坡,平均坡度32°,土壤为山地棕壤;样地内林木平均胸径24.3 cm,平均树高16.6 m,蓄积量277.59 m3/hm2。样地内存在大量的枯立(倒)木,其中枯立木168株,枯倒木37株,最大树木胸径61.3 cm,年龄为120年以上(用生长锥钻木芯数年轮)。样地内灌木树种主要有箭竹(Sinarundinarisnitida)、鞘柄菝葜(Smilaxstans)、青荚叶(Helwingiajaponica(Thunb.) Dietr.)、长柄八仙花(HydrangealongipesFranch.)、桦叶荚蒾(ViburnumbetulifoliumBatal.)、绣线梅(NeilliathrysifloraD.Don)、宝兴茶藨子(RibesmoupinenseFranch.)等;草本有秦岭金腰子(ChrysospleniumbiondianumEngl.)、裂叶荨麻(Urticalotabifolia)、酢浆草(OxaliscorniculataLinn.)、羊胡子(Carexbreviculmisvar.fibrillosa)等。
2 研究方法
2.1 调查方法
在研究区采用NTS320全站仪测设面积为1 hm2(100 m×100 m)的样地,在确定样地的基轴坐标后,用全站仪将整个样地划分为25块小样地(20 m×20 m),样地四角用水泥桩标记。调查记录树高大于1.3 m的乔木树种的胸径、株数、年龄、树高等指标,并用全站仪(GTS-602AF)进行定位;选择样地内胸径最大的1株锐齿栎,用生长锥钻木芯,查数年轮确定林分年龄。采用超声波测高仪测量树高;采用GPS测定样地地理坐标、海拔,用罗盘仪测定样地坡向、坡度。
2.2 群落空间结构参数分析
森林的结构特征体现在时间和空间2个方面,时间特征用林分的直径分布来表现,空间结构特征以林分的水平结构和垂直结构特征来表现。以样地每木定位调查中胸径大于5 cm的林木数据为基础,分析林分的角尺度[12-16]、混交度[14,16-19]和大小比数[13-15,20]。
(1)
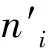
树木大小是林分结构的基本特征,具有重要的生态学意义[21]。与角尺度和混交度的特征相似,大小比数值以频率分布形式刻画了参照树与其最近相邻木之间大小的分化度,其重要意义在于明确了不同大小树木在与其最近4株相邻木组成的结构单元中的地位,可以用树高、胸径、断面积、冠幅、根系、叶面积等作为比较指标,其共有5种可能取值,即0,0.25,0.50,0.75和1.00,分别对应优势、亚优势、中庸、劣势和绝对劣势5种相对关系。当将某个树种作为研究对象时,大小比数反映了该树种在林分中的平均状态,其与相对显著度结合则反映了该树种的优势度。优势度用下式计算:
(2)
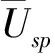
树种优势度的取值为0~1,接近1表示非常具有优势,接近0表示几乎没有优势。
本研究运用空间结构分析软件Winkelmass计算林分空间结构指标,为避免边缘效应对林分结构的影响,设置了5 m缓冲区,核心区面积为90 m×90 m。
3 结果与分析
3.1 锐齿栎原始林的林分特征
林分中胸径大于5 cm林木的基本概况如表1所示。样地林分类型为天然次生林,林分密度1 267株/hm2,总断面积34.46 m2/hm2;林分平均胸径25.1 cm,平均树高17.1 m,蓄积量276.45 m3/hm2。林分中锐齿栎无论是断面积还是频度都相对较大,是该林分中的优势种;辽东栎、桦椴等树种断面积较大,但其相对频度却较低,说明其在林分中主要以大径木的形式存在,而数量却较少;地锦槭、鹅耳枥、鄂椴、小叶鹅耳枥、川鄂鹅耳枥、水榆花楸等的断面积和频度都低于锐齿栎,但与其他树种相比,数量、断面积均在林分中具有相对优势。因此,将上述树种作为主要树种进行分析。
3.2 锐齿栎原始林中林木个体的水平空间分布格局
如图1所示,样地中林木在角尺度为0.5时的分布频率最高,为56%,角尺度为0.75和1.00时的分布频率之和大于角尺度为0和0.25时的分布频率之和,样地林木个体角尺度的均值为0.531,在[0.475,0.517][14]外,说明林分内林木整体分布格局属轻度团状分布。从林分中不同树种的空间结构单元来看,以锐齿栎、鹅耳枥、地锦槭、光叶泡花树、鄂椴、小叶鹅耳枥、川鄂鹅耳枥、水榆花楸为参照树的单元角尺度频率分布与整个林分的分布规律相似,且上述树种总株数在整个样地中所占比例为62.7%,因此以这几个树种为参照树的单元林木分布格局在很大程度上决定了整个林分林木的分布格局。
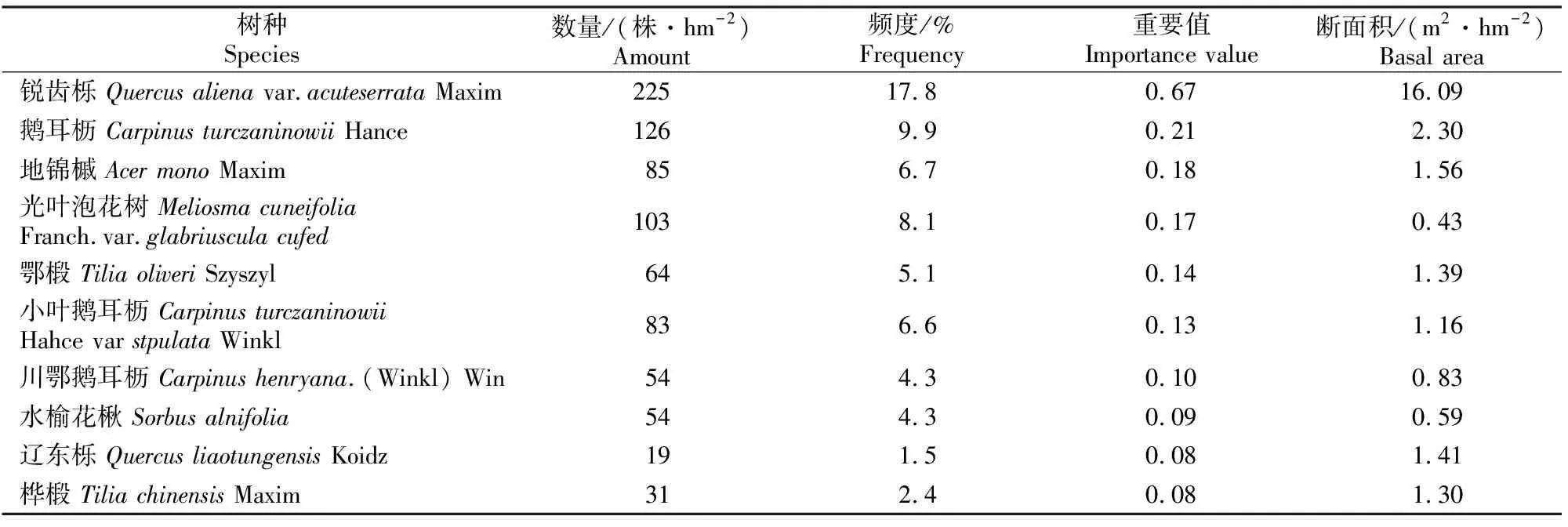
表1 小陇山锐齿栎原始林中主要树种的特征值
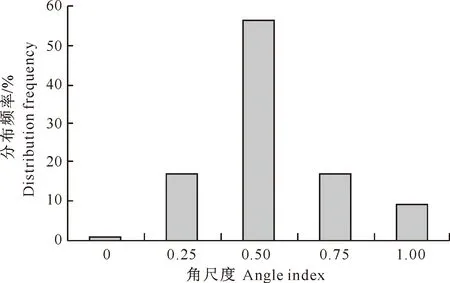
图1 小陇山锐齿栎原始林角尺度的分布频率
林分中主要树种锐齿栎、鹅耳枥、地锦槭、光叶泡花树、鄂椴、小叶鹅耳枥、川鄂鹅耳枥、水榆花楸的平均角尺度依次为0.526,0.586,0.535,0.604,0.556,0.604,0.634,0.559。可见主要树种的角尺度都在[0.475,0.517]外,平均角尺度均略高于0.517,属于轻度团状分布。一般而言,当次生林处于顶级群落时,其水平空间分布格局大多数是随机分布的,但是由于立地微环境的差异或者在枯倒木形成的初期,林木枯倒时砸压幼树、形成林窗、林分更新等都会暂时打破森林原有的稳定状态,进而促使林木的水平空间分布格局向轻度团状分布方向转变,但随着自然演替的进行,林分的水平空间分布格局会逐渐趋近于随机分布[10]。由此可知,样地所处林分目前正处于森林自然演替的中后期,在无人干扰的情况下,随着森林自然演替的发展,样地所处林分将会逐渐发展为当地的顶级群落,也就是说林木的水平空间分布格局将会逐渐趋近于随机分布。
3.3 锐齿栎原始林中树种的空间隔离程度
使用Winkelmass软件计算得出样地的平均混交度为0.71,修正混交度为0.475。小陇山锐齿栎原始林混交度的分布如图2所示。
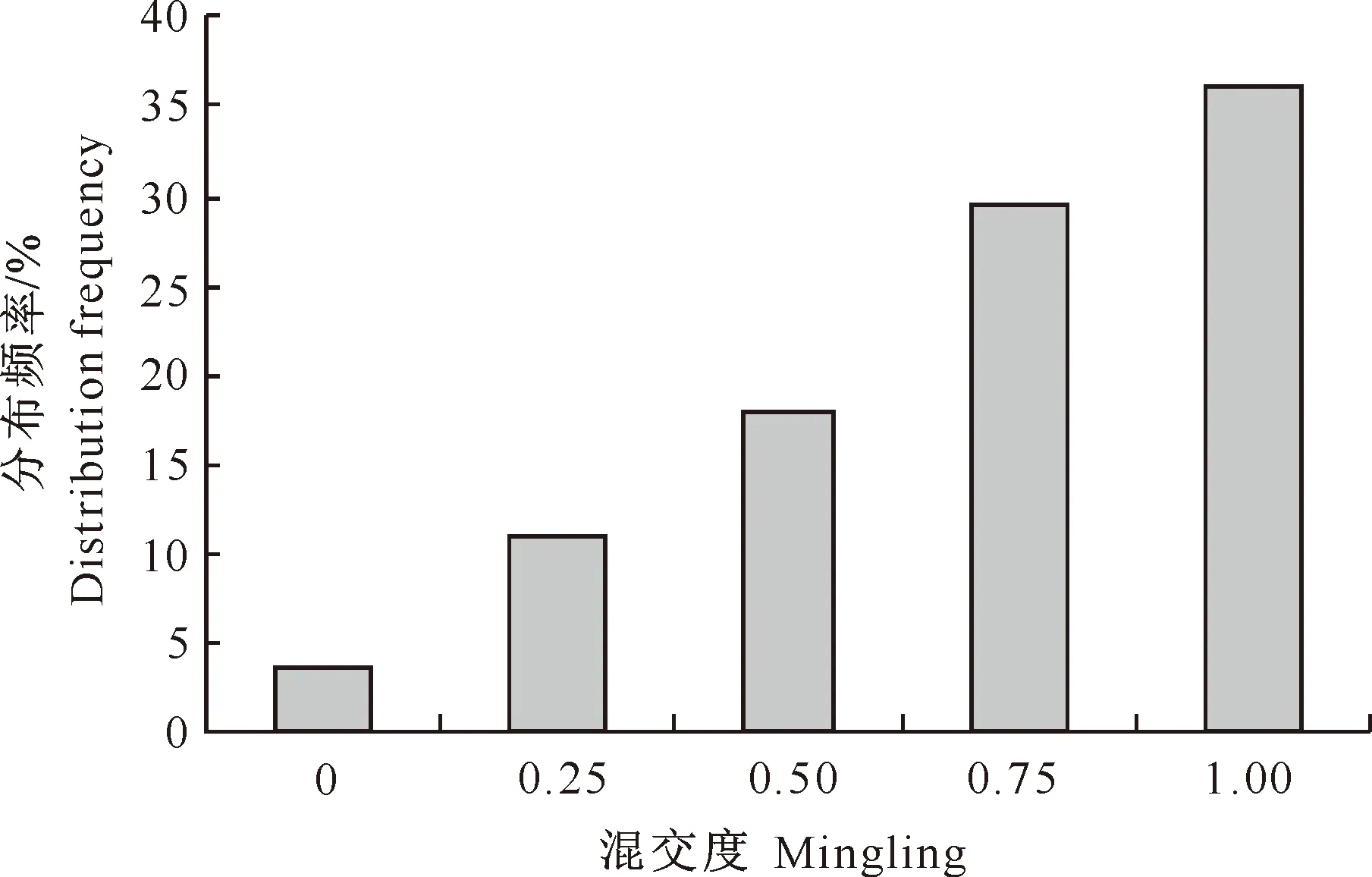
图2 小陇山锐齿栎原始林混交度的分布频率
由图2可知,林分中处于极强度混交(混交度为1.00,即参照树与周围4株最近相邻木不属于同一种)的林木比例为36.8%,处于强度混交(混交度为0.75)的林木比例为31.1%,处于中度混交(混交度为0.5)的林木比例为17.7%,处于弱度混交(混交度为0.25)的林木比例为10.8%,林分中只有3.6%的林木个体处于零度混交(混交度为0)。也就是说,在林分中只有极个别林木与周围最近4株相邻木均为同一树种,同种个体聚集较少,而大多数林木与其他树种相伴而生。
从表2来看,以锐齿栎为参照树的结构单元以强度和极强度混交为主,所占比例分别为39%和30%;鹅耳枥、光叶泡花树、鄂椴、小叶鹅耳枥、水榆花楸出现了零度混交,但所占比例都很小。林分中优势树种和亚优势树种的混交度均主要分布在强度混交和极强度混交中,这说明样地所处林分树种隔离程度很高。有研究表明,群落越向稳定的方向发展,强度和极强度混交的频率越高[22]。出现这一结果是由于在林分内相同树种个体对资源环境利用的一致性,导致种内竞争激烈,发生自疏现象,使最邻近的同种个体数量减少,而其他树种由于对环境资源的利用要求不同,逐步占据并取代由于同种竞争而被淘汰的林木个体的位置,最终形成多树种混交的稳定结构。
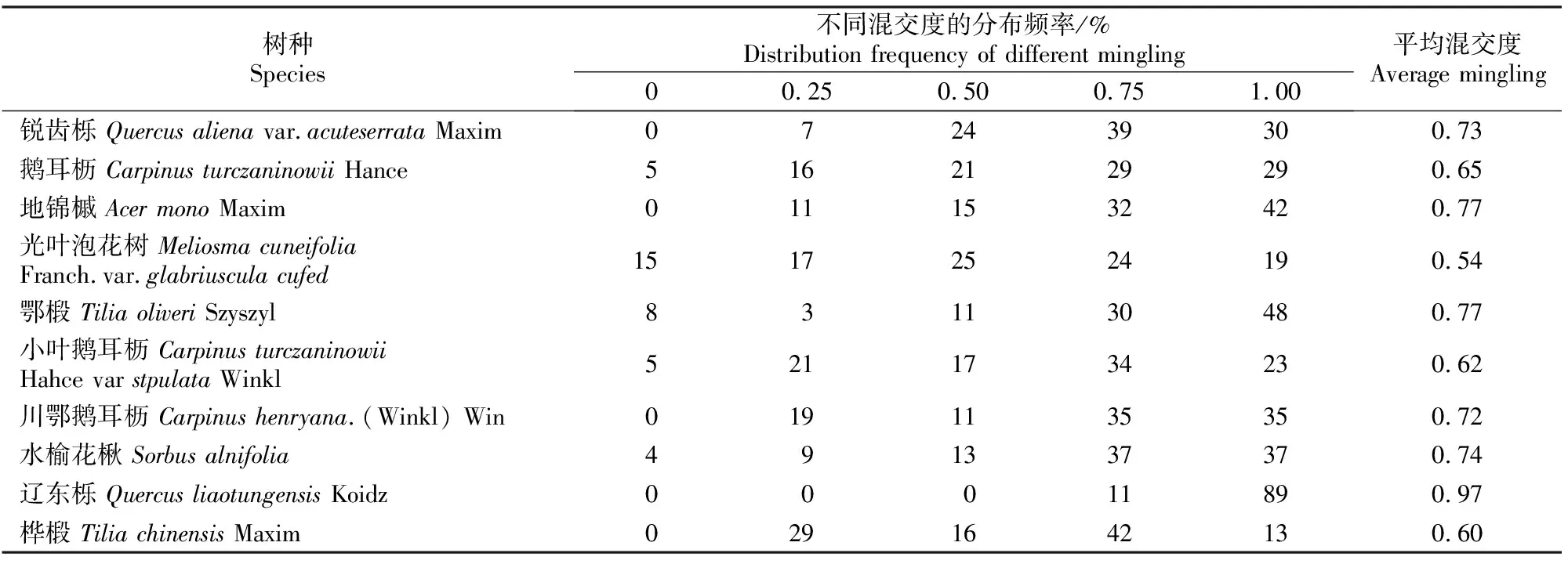
表2 小陇山锐齿栎原始林主要树种的平均混交度及其不同混交度的分布频率
3.4 锐齿栎原始林中林木的大小分化程度
由图3可知,样地内主要树种以胸径作为比较指标的大小比数排列顺序为:辽东栎<锐齿栎<桦椴<鄂椴<鹅耳枥<地锦槭<水榆花楸<小叶鹅耳枥<川鄂鹅耳枥<光叶泡花树。树种大小比数排序说明,在以该树种为参照树的结构单元中,辽东栎和锐齿栎处于优势状态,桦椴处于亚优势状态,而鄂椴、鹅耳枥、地锦槭、水榆花楸、小叶鹅耳枥、川鄂鹅耳枥则处于中庸状态;光叶泡花树受到压迫,处于劣态。
由表3可以看出,林分中辽东栎的大小比数为0和0.25的比例达到了100%,且取值主要集中在大小比数0上,这是由于辽东栎的数量很少并且胸径较大,因而在整个结构单元中处于明显的优势地位。优势树种锐齿栎大小比数频率分布主要集中在大小比数0上,所占比例达到53.3%,大小比数为0.25的个体所占比例为28.4%,说明以锐齿栎为参照树的结构单元,周围其他个体大部分较锐齿栎小或仅有1株比锐齿栎大,林分中仅有少数锐齿栎个体在结构单元中处于中庸、劣态或绝对劣态。而锐齿栎的数量在整个林分中所占比例最大,说明该林分中优势树种的生长受相邻木的干扰较小,锐齿栎的生长在整个林分中占绝对优势。光叶泡花树在整个林分中也占有相当的数量,但其大小比数分布频率多数处于劣态、绝对劣态或中庸状态的地位,这可能是由于其进入群落的时间较晚,该种群尚处于生长发育阶段,生长空间不足,因而造成其在林分中地位低下。
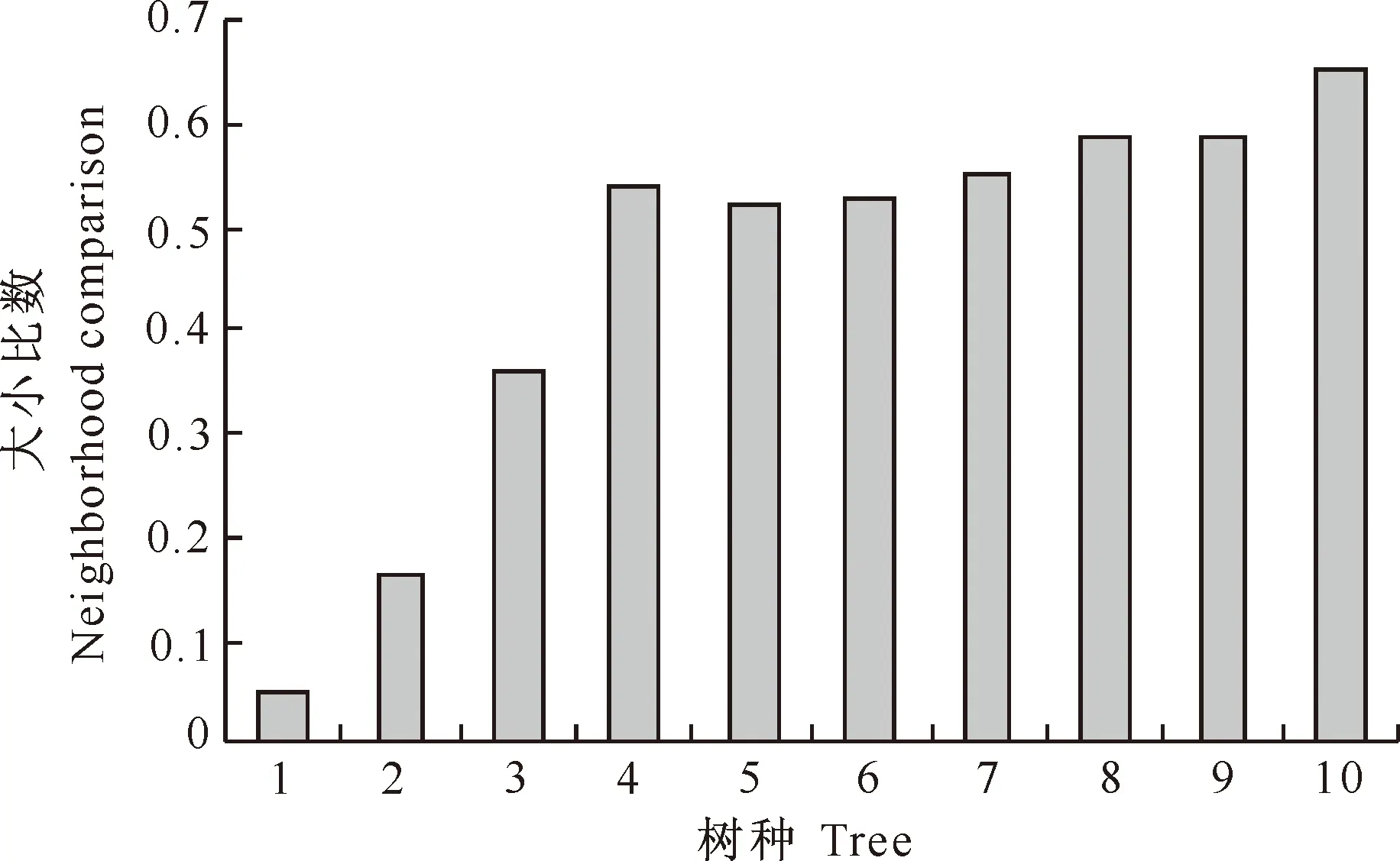
图3 小陇山锐齿栎原始林主要树种平均大小比数
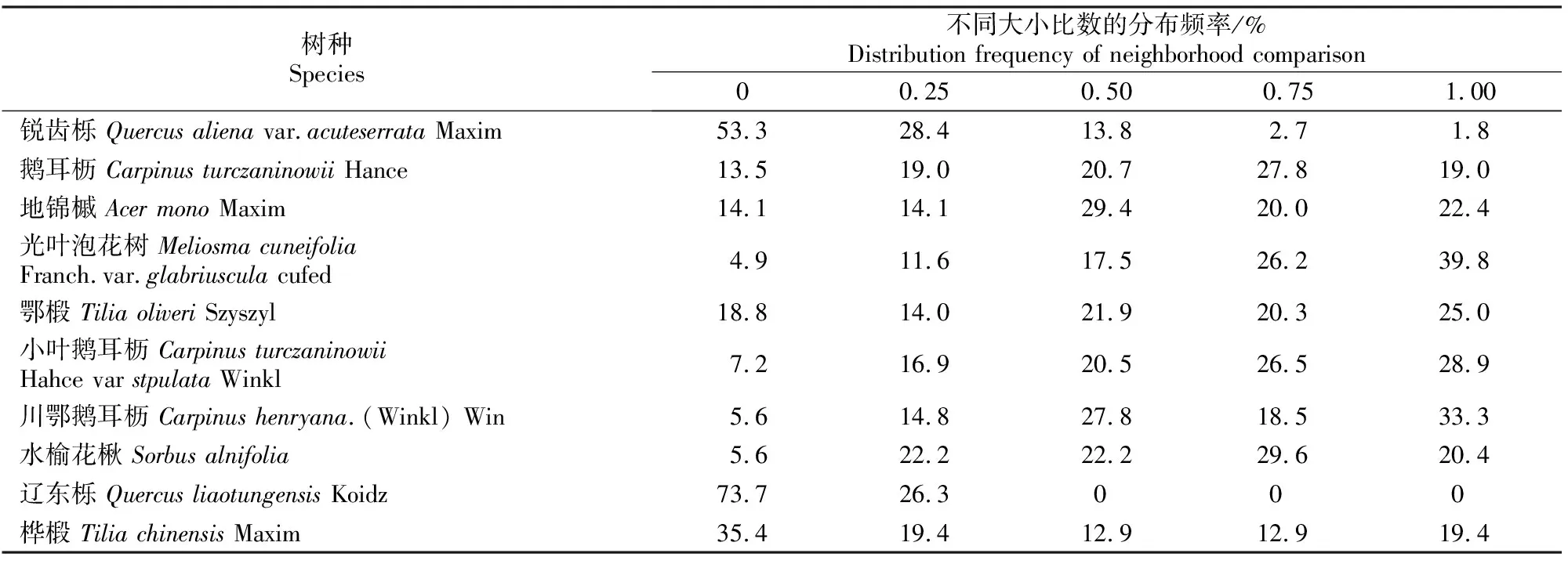
表3 小陇山锐齿栎原始林主要树种大小比数的分布频率
大小比数仅反映了树种在结构单元中的优劣状态,并不能反映树种在整个林分的优劣状态。因此,合理反映树种优势度的方法应该是将树种大小比数与相对显著度相结合。按照此方法计算以上几个树种的优势度,结果(图4)显示,树种大小比数与相对显著度相结合的优势度排列顺序为:锐齿栎>辽东栎>鹅耳枥>桦椴>地锦槭>鄂椴>小叶鹅耳枥>川鄂鹅耳枥>水榆花楸>光叶泡花树。结果表明,在林分中,锐齿栎处于绝对优势状态,辽东栎处于亚优势状态,光叶泡花树受到压迫,处于劣态。
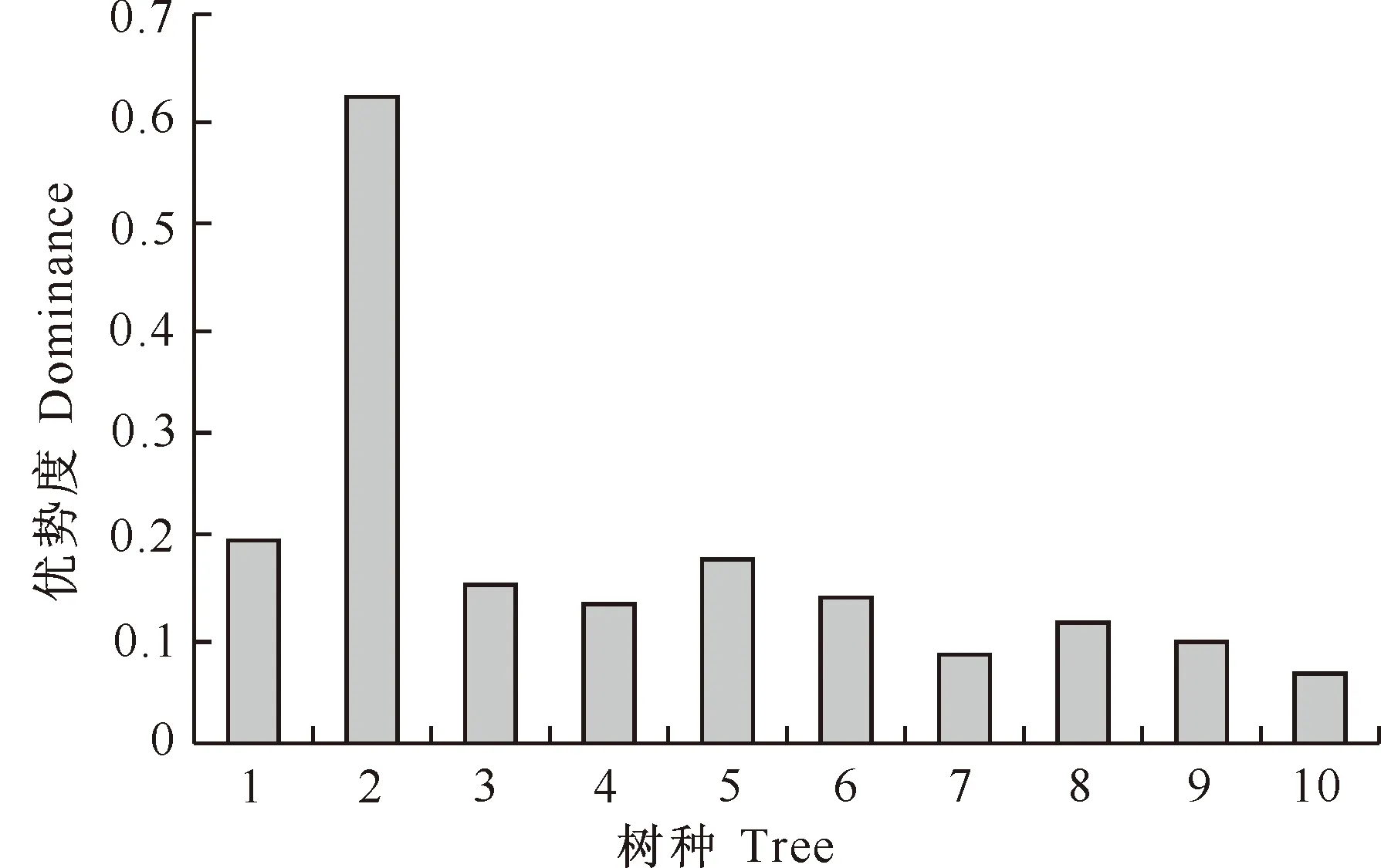
图4 小陇山锐齿栎原始林主要树种的优势度
4 讨论与结论
森林空间结构是森林生长的驱动因子,对森林未来的发展具有决定性作用[23]。原始林是森林生物与环境相互依存和相互作用、长期协同进化的产物,分析原始林的空间结构特征,有助于弄清原始林群落中的种间种内关系、各种群间的空间配置关系以及群落演替规律和生态功能的发挥机制,能够为研究区次生林和人工林经营、天然林群落模仿及顺向演替的人为促进提供理论依据,对实现森林资源的可持续经营具有重要的现实意义。以锐齿栎为主的栎类天然林是小陇山林区主要的林分类型,锐齿栎次生林经营对于小陇山林区实现森林可持续经营至关重要。对小陇山林区锐齿栎原始林样地的调查分析表明,锐齿栎原始林林木分布格局呈轻微团状分布,林分平均角尺度为0.531;样地内各树种呈高度混交,林分平均混交度为0.71,修正混交度为0.475,强度和极强度混交结构单元比例占全林分的67.9%,主要树种的混交度分布频率也偏向于强度和极强度混交。各树种的大小比数平均值差异较大,其中优势树种锐齿栎的数量和大小比数分布频率均处于优势地位;光叶泡花树在整个林分中也占有相当的数量,但其个体在所处的结构单元中,多数较其最近相邻木小,处于劣态、绝对劣态或中庸的地位。
赵中华等[11]研究了小陇山林区响潭沟和白营西沟锐齿栎次生林的空间结构特征,结果表明,响潭沟锐齿栎次生林在皆伐迹地经过长时间的自然恢复后,虽然从林分总体结构特征来看,是一个进展演替的过程,但林分中林木的分布格局仍然呈轻微团状分布,直径分布不尽合理,树种数较少,混交程度也不高,建群种锐齿栎处于亚优势的状态,与本研究中建群种在林分中的地位明显不同;白营西沟林分为人为干扰后形成的林分,由于经历了2次大强度的择伐利用,林木的分布格局为随机分布,但树种混交程度较低,建群种锐齿栎在林分中处于亚优势状态,而先锋树种白桦则处于中庸状态,这也与长期未经干扰的锐齿栎有明显不同。由此可以看出,在对小陇山锐齿栎次生林进行经营时,可根据生态学原理,从优化空间结构的角度出发,调整林分的水平分布格局、树种组成和林内树种的竞争关系,使林分结构更接近于自然状态的林分结构,尽快恢复到合理状态,从而提升林分的整体功能。
[参考文献]
[1] 汤孟平,唐守正,雷相东,等.两种混交度的比较分析 [J].林业资源管理,2004(4):25-27.
Tang M P,Tang S Z,Lei X D,et al.Comparison analysis on two minglings [J].Forest Resources Management,2004(4):25-27.(in Chinese)
[2] 惠刚盈.角尺度:一个描述林木个体分布格局的结构参数 [J].林业科学,1999,35(1):37-42.
Xi G Y.The neighbourhood pattern:A new structure parameter for describing distribution of forest tree position [J].Scientia Silvae Sinicae,1999,35(1):37-42.(in Chinese)
[3] 雷相东,唐守正.林分结构多样性指标研究综述 [J].林业科学,2002,38(3):140-146.
Lei X D,Tang S Z.Indicators on structural diversity within-stand:A review [J].Scientia Silvae Sinicae,2002,38(3):140-146.(in Chinese)
[4] 惠刚盈,Gadow K V.德国现代森林经营技术 [M].北京:科学技术出版,2001:66-134.
Xi G Y,Gadow K V.Modern forest management of Germany [M].Beijing:Science and Technology Press,2001:66-134.(in Chinese)
[5] 惠刚盈.基于相邻木关系的林分空间结构参数应用研究 [J].北京林业大学学报,2013,35(4):1-9.
Xi G Y.Studies on the application of stand spatial structure parameters based on the relationship of neighborhood trees [J].Journal of Beijing Forestry University,2013,35(4):1-9.(in Chinese)
[6] Moeur M.Characterizing spatial patterns of trees using stem ma-pped data [J].For Sci,1993,39(4):756-775.
[7] 汤孟平.森林空间结构分析与优化经营模型研究 [D].北京:北京林业大学,2003.
Tang M P.Analysis and optimal management model of forest spatial structure [D].Beijing:Beijing Forestry University,2003.(in Chinese)
[8] 索安宁,巨天珍,张俊华,等.甘肃小陇山锐齿栎群落生物多样性特征分析 [J].西北植物学报,2004,24(10):1877-1881.
Suo A N,Ju T Z,Zhang J H,et al.Analysis of biodiversity characteristics ofQuercusalienavar.acuteserratacommunity on Mt.Xiaolong in Gansu [J].Acta Botanica Boreali-occidentalia Sinica,2004,24(10):1877-1881.(in Chinese)
[9] 索安宁,巨天珍,张俊华,等.甘肃小陇山锐齿栎群落生物量动态研究 [J].生态学杂志,2005,24(4):377-381.
Suo A N,Ju T Z,Zhang J H,et al.Biomass structure ofQuercusalienavar.acuteserratcommunity on Mt.Xiaolongshan in Gansu [J].Chinese Journal of Ecology,2005,24(4):377-381.(in Chinese)
[10] 巨天珍.天水小陇山锐齿栎群落种间关系定量分析 [J].西北植物学报,1995,15(3):250-253.
Ju T Z.A quantitation study of the intespecific association ofQuercusalienavar.acuteserratacommunity in Xiaolongshan Mountain of Tianshui [J].Acta Bot Boreal-Occident Sin,1995,15(3):250-253.(in Chinese)
[11] 赵中华,惠刚盈,袁士云,等.小陇山锐齿栎天然林空间结构特征 [J].林业科学,2009,45(3):1-6.
Zhao Z H,Hui G Y,Yuan S Y,et al.Spatial structure characteristic ofQuercusalienavar.acuteserratanatural forest in Xiaolongshan [J].Scientia Silvae Sinicae,2009,45(3):1-6.(in Chinese)
[12] Gadow K V,Hui G Y,Albert M.Das winkelmass-ein strukturparameter zur beschreibung der individualverteilung in waldbeständen [J].Centralblatt für das Gesamte Forstwesen,1998,115(1):1-10.
[13] Aguirre O,Hui G Y,Gadow K V,et al.An analysis of spatial forest structure using neighbourhood-based variables [J].Forest Ecology and Management,2003,183:137-145.
[14] 惠刚盈,Gadow K V,胡艳波,等.结构化森林经营 [M].北京:中国林业出版社,2007.
Xi G Y,Gadow K V,Hu Y B,et al.Tructure-based forest management [M].Beijing:China Forestry Press,2007.(in Chinese)
[15] Albert M.Analyse der eingriffsbedingten strukturveänderung und durchforstungsmodellierung in mischbeständen [J].Dissertation Universität Göttingen Hainholz Verlag,1999,201S:63-68.
[16] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究 [J].林业科学研究,2001,14(1):23-27.
Xi G Y,Hu Y B.Measuring species spatial isolation in mixed forests [J].Forest Research,2001,14(1):23-27.(in Chinese)
[17] Gadow K V.Zur bestandesbeschreibung in der forsteinrichtu-ng [J].Forst und Holz,1993,48(21):602-606.
[18] Füldner K.Strukturbeschreibung von buchen-edellaubholz-mi-schwäldern [D].Göttingen:Cuvillier Göttingen,1995.
[19] 惠刚盈,赵中华,袁士云.森林经营模式评价方法:以甘肃小陇山林区为例 [J].林业科学,2011,47(11):114-120.
Xi G Y,Zhao Z H,Yuan S Y.Evaluation method of forest management models:A case study of Xiaolongshan forest area in Gansu Province [J].Scientia Silvae Sinicae,2011,47(11):114-120.(in Chinese)
[20] 惠刚盈,Gadow K V,Albert M.一个新的林分空间结构参数:大小比数 [J].林业科学研究,1999,12(1):1-6.
Xi G Y,Gadow K V,Albert M.A new parameter for stand spatial structure neighbourhood comparison [J].Forest Research,1999,12(1):1-6.(in Chinese)
[21] Kuuluvainen T,Penttinen A, Leinonen K,et al.Statistical opportunities for comparing stand structural heterogeneity in managed and primeval forests:An example from boreal spruce forests in Southern Finland [J].Silva Fennica,1996,30: 315-328.
[22] 安惠君.阔叶红松林空间结构特征研究 [M]//唐守正.东北天然林生态采伐更新技术研究.北京:中国科学技术出版社,2005:547-553.
An H J.Study on the spatial structure characters of broad-leaved Korean pine forest [M]//Tang S Z.Research on techniques of ecology-based harvesting and regeneration for natural forest in Northeast of China.Beijing:China Science and Technology Press,2005: 547-553.(in Chinese)
[23] Pretzsch H.Analysis and modeling of spatial stand structures Methodological considerations based on mixed beech-larch stands in Lower Saxony [J].Forest Ecology and Management,1997,3:237-253.
猜你喜欢
阿来研究(2020年1期)2020-10-28
现代园艺(2018年2期)2018-03-15
现代园艺(2017年2期)2017-03-16
新世纪水泥导报(2016年1期)2016-07-01
中央社会主义学院学报(2016年2期)2016-05-04
土木建筑工程信息技术(2013年1期)2013-10-17
