
长江口北支浮游植物群落结构周年变化特征
2014-03-25 06:32薛俊增吴惠仙
生物学杂志 2014年3期
刘 笑, 薛俊增, 吴惠仙
(上海海洋大学 水产种质资源发掘与利用教育部重点实验室,上海 201306)
河口是淡水与海水之间的生态交错区, 该区域的环境因子变化与河流和海洋均有差异, 具有自己的特殊性。浮游植物是河口生态系统的重要初级生产者,食物链的基础,其群落结构的变化直接影响河口生态系统的结构和功能[1]。浮游植物的时空变化与环境因子的关系密切[2-4],因此,其群落结构特征在一定程度上反映了河口区生态状况。国内外学者对河口浮游植物的种类组成和在河口区的时空分布,以及光、温度、盐度、营养盐和径流等主要环境因子对初级生产力的影响有着深入的研究[2-25]。国内的研究注重对浮游植物种类组成和水质对浮游植物的影响,且常常设置较多的采样点和多断面[2-4,8-25],而国外研究着重长期监测,重视风、降雨等气象因素对浮游植物群落的影响[5-7]。长江口地处中国东南沿海,是中国最大的河口。长江口在径流、潮汐、沿岸流、降水等多种环境因子的共同作用下,形成了复杂的水域生态环境。从20世纪90年代开始,国内学者对长江口浮游植物的种类组成、丰度时空分布、生物多样性及浮游植物群落与环境因子相关性等方面进行了研究[8-25],但其研究的水域主要集中在长江口的南支和口外海域[4,9-25],有关长江口北支浮游植物的报道较少[8],长江口北支水面狭窄,仅能接受长江1%的径流[26],受潮汐的影响大,形成了独特的高盐水域环境,通过对长江口北支浮游植物分布的研究,分析长江口北支特有水文现象对该水域浮游植物的影响,为进一步阐明长江口北支生态环境变化对水生生物影响提供依据。
1 材料和方法
1.1 样本的采集与处理
在长江口北支共设置5个采样点:S1、S2、S3、S4、S5(图1),按照《海洋生态环境监测技术规程》[27]对长江口浮游植物水样进行采样。2010年10月—2011年9月,每月进行一次浮游植物标本采集,采集表层水1 L,采集后立即滴加15 mL鲁哥氏液固定,带回实验室静置沉淀48 h 弃上清浓缩至50 mL 后滴加福尔马林保存,显微镜镜检,进行浮游植物的分类和计数。各站位取表层水,立即经0.45 μm微孔滤膜过滤(预先采用1∶ 1000 HCl浸泡24 h并用双蒸馏水冲洗至中性)并冷冻保存于250 mL聚乙烯瓶中带回实验室,用于氨氮、硝酸盐测定。总氮、总磷和浊度经现场水样装入1 L聚乙烯瓶中常温保存,24 h内完成测定。测定方法根据《海洋生态环境监测技术规程》[27]相关内容进行。水温、盐度、电导率、酸碱度、溶解氧含量等指标使用 YSI 85型号的手持式野外水质检测仪在采样现场进行测定。
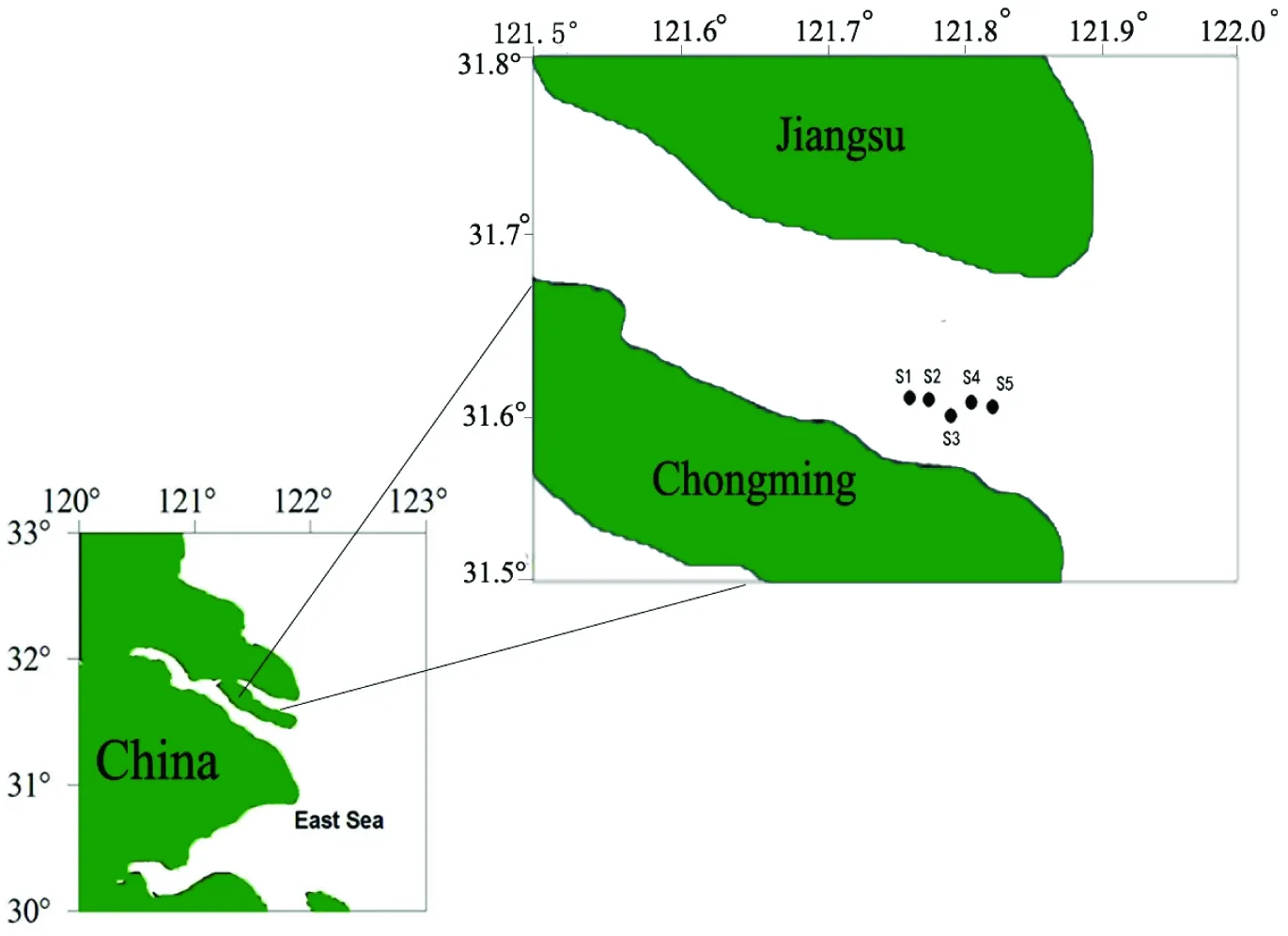
图1 长江口北支采样点分布情况示意图
1.2 数据分析
生物多样性指数采用Shannon-Wiener指数[28]、Margalef 指数[29]、Pielou均匀度指数[30]。
1)Shannon-Wiener多样性指数(H):
式中:ni为第i种物种的个体数,N为总个体数,S为物种总数;
2)Margalef 丰富度指数(D):
式中:S表示物种总数,N为总个体数。
3)Pielou 均匀度指数(J):
式中:H为多样性指数,S为物种总数
浮游植物优势度的确定[31]:
式中:Y为优势度,ni为第i种物种个体数,N为总个体数,fi为第i种物种出现频率。当Y值大于0.02时,表示该物种为优势种。
通过数据分析软件SPSS 16.0对长江口北支浮游植物丰度进行One-way ANOVA分析和浮游植物丰度与环境因子Pearson相关性分析。
2 结果
2.1 浮游植物种类组成周年变化
共采集到浮游植物183种(包括变种和变型),隶属于7门60属,其中硅藻门种类最多,有35属137种,占总种类数的75%;其次为绿藻门,有11属21种,占11%;蓝藻门7属14种,占8%;黄藻门1属4种,占2%;裸藻门3属4种,占2%;甲藻门2属2种,占1%;金藻门1属1种,占1%。对长江口北支浮游植物各个月份的优势种进行分析(表1),发现大多数的月份的优势种都来自硅藻门,但在3月份,优势种仅为3种且经常作为水华种的蓝藻门种类占了优势地位;在9月份,优势种中出现了黄藻门黄丝藻属的种类,该属种类为淡水种。具槽直链藻、中肋骨条藻和条纹小环藻是全年优势种中优势度最高的3个种类;具槽直链藻在4月、5月、8月、11月、12月都是优势度最高的优势种;中肋骨条藻在6月份是优势度最高的优势种;条纹小环藻在9月、10月、12月是优势度最高的优势种。
2.2 浮游植物多样性的周年变化
分析Shannon-Wiener 多样性指数,Pielou 均匀度指数和Margalef丰富度指数,结果见图2。从图2可以看出,全年均匀度指数变化平稳,仅在3月份骤然下降,降至0.5以下,其余各月均在0.55~0.85之间波动,1月的均匀度最高,达到了0.85;丰富度指数全年变化幅度较大,在1.8~5.5之间,9月和10月丰富度指数在4左右波动,进入12月丰富度指数开始降低,并一直持续到1月,两2个月丰富度指数均在2.6以下,2月丰富度指数升高,超过了前一年9月的水平,但在3月丰富度指数又反常的下降到了1月的水平,仅为2.3, 3月至6月丰富度指数逐月上升,但到了7月,又一次出现急降,仅次于全年最低的12月,8月和9月保持平稳,丰富度指数在全年中相对中等水平,在3.4左右;Shannon-Wiener指数在1.6~3.4之间,变化规律同丰富度指数相似,但峰值没有在6月出现,而是出现在了2月,6月的Shannon-Wiener指数相对于5月反而降低了。

表1 长江口北支浮游植物各月优势种、平均丰度及优势度
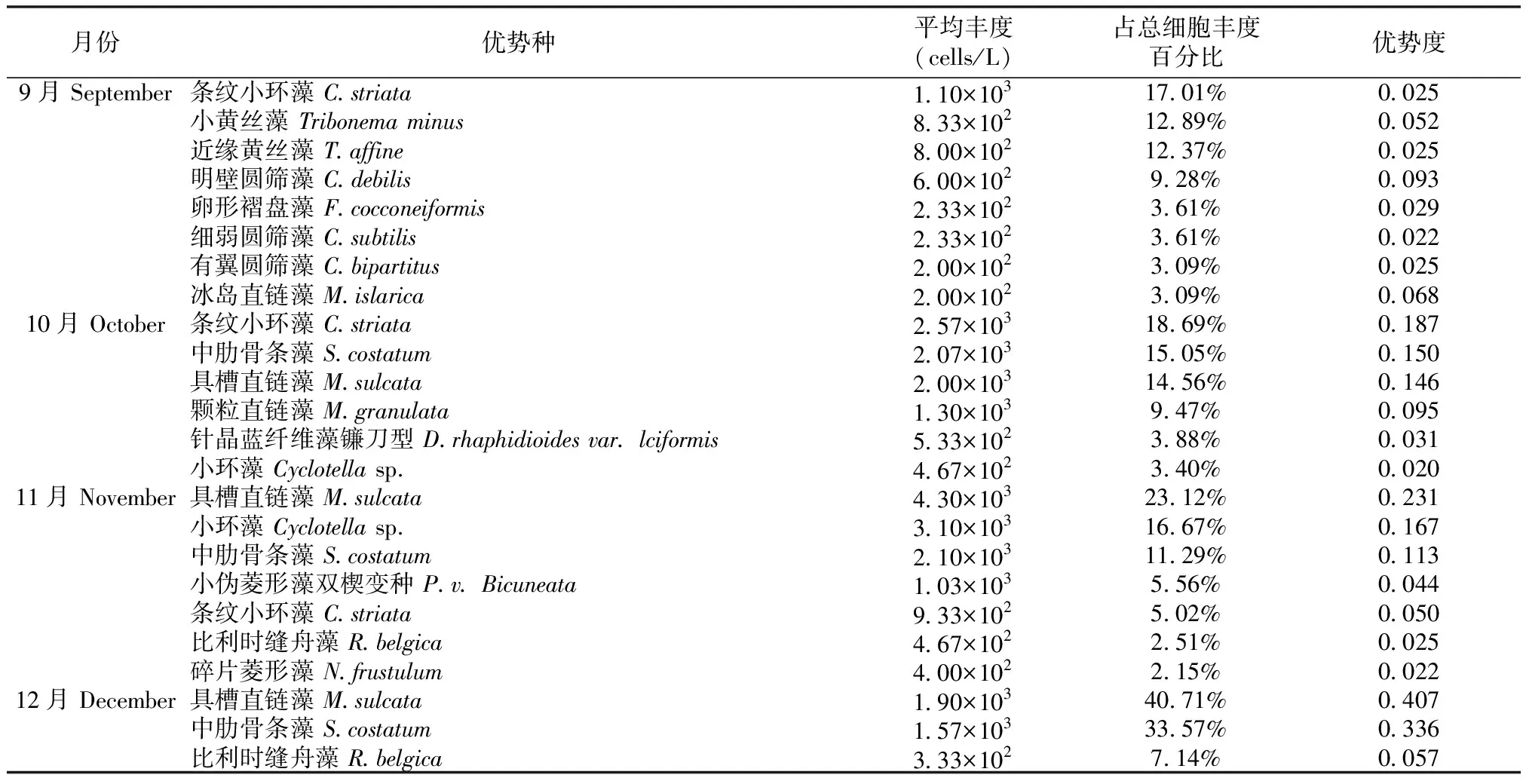
续表1(Table 1 Continued)

2.3 浮游植物丰度周年变化
长江口北支浮游植物全年丰度在2.83×103~6.18×104cells/L之间,全年浮游植物丰度的分布为单峰型,6月的丰度最高,1月的丰度最低,全年的平均丰度为1.73×104cells/L(图3)。方差分析显示,四个季节之间浮游植物的平均丰度差异不显著(P>0.05),但月份之间浮游植物的丰度的差异明显(P<0.05);10月和11月浮游植物丰度为上升趋势,但进入冬季12月和1月丰度降到了一年中的最低水平,2月至5月,浮游植物丰度有所回升,变化较为平稳,在2.00×104cells/L左右波动;进入6月,浮游植物丰度急剧上升,达到峰值并远远高于其他月的丰度,在6月和7月之间,浮游植物的丰度出现骤降,降低至冬季12月和1月的丰度水平,8月浮游植物的丰度又有了明显的回升,同5月的浮游植物丰度接近,而到了9月丰度再次下降,且在较低的水平,略高于7月。

图3 长江口北支浮游植物丰度周年变化
3 讨论
长江口北支浮游植物的丰度和多样性具有明显季节变化,主要受到温度、盐度和营养盐的影响。冬季的12月和1月,丰度和多样性指数都在下降,处于一年中最低的时期,这是因为温度成了限制因子,较低的温度抑制浮游植物的生长,这一结果同以往长江口南支和口外海域浮游植物的变化规律相似,冬季浮游植物种类数和丰度都较低[14]。进入2月,随着温度的上升,浮游植物的丰度和多样性都有所升高;本次调查硅藻所占比例最大,但在3月蓝藻成为优势种,浮游植物丰度较2月有所升高(图3),但多样性指数、丰富度和均匀度较2月都有下降(图2),表明该月虽然丰度上升,但物种数量反而下降,多样性降低,这可能是由于在3月,本次调查水域的硝酸盐和氨氮都达到了一年中的最高值,丰富的营养盐促使浮游植物大量繁殖,蓝藻相对于硅藻生长更迅速,从而取代了硅藻的优势地位。到了4月,以中肋骨条藻和具槽直链藻为代表的硅藻重新占据优势,它们对长江口北支高盐水域环境有更强的适应性。在6月浮游植物的丰度达到顶峰,丰富度指数最高,但均匀度指数和Shannon-Wiener 多样性指数相对较低,说明该月种类丰富,但伴生种较多,优势种的优势地位较大。由表3可知该月中肋骨条藻占该月总密度的41.35%,中肋骨条藻的大量繁殖导致了多样性指数较低,在长江口其他水域,浮游植物丰度在6月同样是一年中的最高[14,18,25],这一变化规律在长江口普遍适用。在7月不论是浮游植物丰度还是多样性指数都出现骤降,中肋骨条藻优势度下降明显,而淡水种颗粒直链藻成为第一优势种。本次调查中,盐度由6月的17下降到7月的10,可见该水域在在7月受到了淡水大量输入的影响,使得群落结构急剧变化,半咸水种的繁殖受到抑制,淡水的输入在7月成了关键影响因子。8月浮游植物的丰度和多样性指数较7月上升,第一优势种为具槽直链藻,到了9月浮游植物丰度再次发生明显的下降,盐度在9月由13上升到了20,高盐度海水的入侵限制了浮游植物的生长,进入10月和11月,不论是浮游植物丰度还是多样性指数都在稳步上升,这可能是由于该段时期水域环境相对稳定利于浮游植物的繁殖。通过对6月—11月浮游植物群落结构的分析可以看出,在这个时间段,浮游植物在冲淡水和盐水入侵的共同作用下,不断的发生群落结构的变动,淡水种和半咸水种交替占据上峰,这一现象在长江口其他水域未见报道,是北支特有的变化规律。
进入20世纪,国内学者对长江口浮游植物的调查增多,但都把调查水域设在了南支和口外海域。孙军等对长江口口外海域进行浮游植物季度调查,优势种主要为中肋骨条藻和具槽直链藻,浮游植物丰度在4.97×103~1.47×105cells/L之间[4,13,15-20];李俊龙等对长江口外海域的调查中,中肋骨条藻为优势种,浮游植物平均丰度为8.82×104cells/L[24];陈立婧等对长江口南支九段沙水域进行周年调查,中肋骨条藻为优势种,周年平均丰度为9.23×105cells/L[25];与上述研究相比发现,长江口及其邻近海域的优势种相似,中肋骨条藻在这一水域普遍存在,这是因为中肋骨条藻的适盐范围更大,易于在长江口这样盐度变化范围广且多变的水域建立优势种群;北支浮游植物丰度低于邻近水域,主要是因为营养盐随径流进入长江口,而北支的径流量明显少于南支,造成北支水域缺乏足够的营养盐支持浮游植物大量繁殖,本研究中,浮游植物丰度与氨氮极显著正相关(P<0.01),与硝酸盐显著正相关(P<0.05)(表3),可见氮是限制北支水域浮游植物生长的关键因素。

表2 长江口北支浮游植物丰度与环境因子Pearson相关性分析
注:* *—P<0.01 相关性极显著 (双侧); *—P<0.05 相关性显著(双侧)。
通过与以往研究的年际间比较和本次调查周年各月比较,我们可以发现,长江口水域适宜广温广盐的小型硅藻生长。浮游植物丰度受温度、盐度和营养盐的影响明显,硝酸盐和氨氮的增加可以促进浮游植物的繁殖。
参考文献:
[1]郭沛涌, 沈焕庭. 河口浮游植物生态学研究进展[J]. 应用生态学报, 2003, 14(1):139-142.
[2]田家怡. 黄河三角洲附近海域浮游植物多样性[J].海洋环境科学, 2000,19(2):38-42.
[3]戴 明, 李纯厚, 贾晓平, 等.珠江口近海浮游植物生态特征研究[J].应用生态学报, 2004,15(8):1389-1394.
[4]栾青杉, 孙 军, 宋书群, 等. 长江口夏季浮游植物群落与环境因子的典范对应分析[J]. 植物生态学报, 2007, 31(3) :445-450.
[5]Cho C H, Huh S H. Community structure and distribution of phytoplankton in the Naktong River estuary ( in Korean) [J] . Ocean Res, 1988, 10(1) : 39-45.
[6]Muylaert K, Sabbe K. Spring phytoplankton assemblages in and around the maximum turbidity zone of the estuaries of the Elbe( Germany), the Schelde ( Belgium/ The Netherlands ) and the Gironde ( France) [J]. J Mar Syst, 1999, 22(2) : 133-149.
[7]Bursa A S. Phytoplankton on coastal waters of the Arctic Ocean at point barrow, Alaska [J]. Arctic, 1963, 16(4) : 239-258.
[8]沈 竑, 徐 韧, 王桂兰, 等. 上海市海岛周围水域浮游植物的调查研究[J]. 海洋通报, 1995, 14(4):26-37.
[9]杨蕉文, 华 棣, 顾新根. 长江口羽状锋海区浮游植物的生态研究[J]. 东海海洋, 1994, 12(1):47-57.
[10]徐兆礼, 白雪梅, 袁 骥, 等. 长江口浮游植物生态研究[J]. 中国水产科学, 1999, 6(5):53-54.
[11]顾新根, 袁 骥, 沈焕庭, 等. 长江口最大浑浊带浮游植物的生态研究[J]. 中国水产科学, 1995, 2(1):16-27.
[12]王云龙,袁 骥,沈新强,等. 长江口及邻近水域春季浮游植物的生态特征[J]. 中国水产科学, 2005, 12(3): 300-311.
[13]孙 军, 赵 冉, 张利永, 等. 春末三峡大坝首次蓄水期长江口浮游植物群集[J]. 应用生态学报, 2011, 22(4): 1045-1052.
[14]林峰竹, 吴玉霖, 于海成, 等. 2004年长江口浮游植物群落结构特征分析[J]. 海洋与湖沼, 2008, 39(4): 402-407.
[15]栾青杉, 孙 军, 宋书群, 等.2004年秋季长江口及其邻近水域浮游植物群集[J]. 海洋科学进展, 2008, 26 (3) : 364-371.
[16]何 青, 栾青杉, 孙 军, 等. 长江口及其邻近水域冬季浮游植物群集[J]. 应用生态学报, 2007, 18(11): 2560-2571.
[17]栾青杉, 孙 军. 2005年秋季长江口及其邻近水域浮游植物群集[J]. 长江流域资源与环境, 2010, 19(2): 201-209.
[18]栾青杉, 孙 军. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系[J]. 生态学报, 2010, 30 (18): 4967-4975.
[19]何 青,孙 军,栾青杉,等. 冬季长江口及其邻近水域的浮游植物[J]. 海洋环境科学, 2009, 28(4):360-365.
[20]王 丹,孙 军,周 峰,等. 2006年6月长江口低氧区及其邻近水域浮游植物[J]. 海洋与湖沼, 2008, 39(6): 619-631.
[21]李 云,李道季,唐静亮,等. 长江口及毗邻海域浮游植物的分布与变化[J]. 环境科学, 2007, 28(4):719-728.
[22]王云龙,袁 骥,庄 平,等. 长江口中华鲟自然保护区附近水域浮游植物分布[J]. 长江流域资源与环境, 2008, 17(1) :72-81.
[23]吴玉霖,傅月娜,张永山,等. 长江口海域浮游植物分布及其与径流的关系[J]. 海洋与湖沼, 2004, 35 (3): 246-257.
[24]李俊龙,郑丙辉,刘录三,等. 长江口浮游植物群落特征及其与环境的响应关系[J]. 环境科学研究,2013, 26 (4) :34-54.
[25]陈立婧,蒋 琰,龚小玲,等. 长江口九段沙湿地近岸水域浮游植物群落结构的特征[J]. 上海海洋大学学报, 2013, 22 (1): 74-79.
[26]陈吉余,陈沈良. 长江口生态环境变化及对河口治理的意见[J]. 水利水电技术, 2003, 34(1): 19-25.
[27]国家质量技术监督局.海洋监测规范[S].2007.
[28]Shannon C E, Weaver W. The mathematical theory of communication[M]. Urbana L: University of Illinos Press, 1949.
[29]Belaoussoff S, Kevan P G, Murphy S, et al. Assessing tillage disturbance on assemblages of ground beetles by using arrange of ecological indices[J]. Biodiversity and Conservation, 2003, 12: 851-882.
[30]Pielou E C. An introduction to mathematical ecology[M]. New York: Wiley-Interscience, 1969.
[31]Habib O A, Tippett R, Murphy K J. Seasonal changes in phytoplankton community structure in relation to physic-chemical factors in Loch Lomond [J]. Hydrobiologia, 1997, 350: 63-79.
[32]王 奎,陈建芳,金海燕,等.长江口及邻近海区营养盐结构与限制[J]. 海洋学报, 2013, 35(3):128-136.
[33]黄自强, 暨卫东. 长江口水中总磷、有机磷、磷酸盐的变化特征及相互关系[ J] .海洋学报, 1984, 16 (1) : 51-60.
[34]李 峥, 沈志良, 周淑青, 等. 长江口及其邻近海域磷的分布变化特征[ J] . 海洋科学, 2007, 31 (1) : 28-42.
[35]孟 伟, 秦延文, 郑丙辉, 等.长江口水体中氮、磷含量及其化学耗氧量的分析[ J] .环境科学, 2004, 25 ( 6) : 65-68.
猜你喜欢
水上消防(2021年3期)2021-08-21
水上消防(2020年5期)2020-12-14
文史春秋(2019年11期)2020-01-15
智能城市(2018年7期)2018-07-10
海洋渔业(2016年6期)2016-04-16
海洋渔业(2016年6期)2016-04-16
Coco薇(2015年5期)2016-03-29
中国当代医药(2015年36期)2015-03-11
小学生时代·综合版(2014年12期)2015-01-17
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
