
高温短程反硝化菌Brevibacillus sp.XF-03特性及其降解动力学
2014-03-17 11:46郝敏娜,杨云龙,葛启隆
生物技术通报 2014年1期
高温短程反硝化菌Brevibacillus sp.XF-03特性及其降解动力学
郝敏娜 杨云龙 葛启隆
(太原理工大学环境科学与工程学院,太原 030024)
采用梯度驯化方法,使得菌株Brevibacillus sp.XF-03在高温(50℃)条件下,能够降解1 000 mg/L亚硝态氮,并通过单因素试验对其生长碳源和C/N进行优化,结果显示,菌株Brevibacillus sp.XF-03短程反硝化最适碳源为琥珀酸钠,C/N为12∶1。在此最佳条件下,42 h对初始浓度为100 mg/L亚硝态氮去除率为95.1%。对该菌株亚硝态氮降解动力学过程进行模拟,符合基质抑制型的 Haldane 模型,各参数分别为:最大比降解速率(μmax)=1.28 h-1,半饱和常数(KS)=451.42 mg/L,底物抑制常数(Ki)=176.77 mg/L。初步探讨了亚硝酸盐还原酶(NIR)活性,在该菌株生长指数期的后期,亚硝酸盐还原酶比活力达0.279(U/ mg protein)。
Brevibacillus sp.XF-03 高温 短程反硝化 亚硝态氮降解动力学 酶活性
随着工业化的发展,水体富营养化现象日趋严重,去除水中氮素污染已成为当今水污染防治领域的一个热点问题[1,2]。目前普遍认为生物脱氮是从污水中去除氮素污染的经济有效的方法之一[3,4]。在诸多污水脱氮新技术中,应用广泛、发展较快及最经济高效的有短程硝化反硝化、同步硝化反硝化及厌氧氨氧化等,它们因具有节省曝气量、提高反应速率、节约碳源、缩短反应时间等优点[5],越来越受到人们的重视[6,7],实现这些新技术的关键是获得具有较高脱氮能力的微生物。此外,以上新技术多适用处理中低温环境的污水[8,9],若处理高温污水则需对污水进行降温,这给生物法处理高温污水带来较大困难。
本文研究菌株Brevibacillussp.XF-03的高温短程反硝化特性及其亚硝态氮降解动力学,以期为短程硝化反硝化工艺的应用以及传统反硝化过程中亚硝
态氮积累问题的解决提供技术参数与新的菌源。
1 材料与方法
1.1 材料
1.1.1 供试菌株 供试菌株:Brevibacillussp.XF-03,来源于太原理工大学环境科学与工程实验室,经鉴定为短芽孢杆菌属(Brevibacillussp.),GenBank登录号为 KF052620。试验初步证明在高温条件(50℃)下,该菌具有较好的短程反硝化能力。
1.1.2 培养基 LB富集培养基(g/L):胰蛋白胨10;酵母浸出粉 5;NaCl 10;pH7.0-7.5。
短程反硝化培养基(g/L):NaNO2适量;KH2PO41.5;Na2PO4·7H2O 7.9;MgSO4·7H2O 0.1;琥珀酸钠适量;微量元素溶液2 mL;pH 7.0-7.5。
微量元素溶液成分(g/L):EDTA 50.0;ZnSO42.2;CaCl25.5;MnCl2·4H2O 5.06;FeSO4·7H2O 5.0;(NH4)6Mo7O2·4H2O 1.1;CuSO4·5H2O 1.57;CoCl2·6H2O 1.61;pH7.0-7.5。
以上培养基121℃高压灭菌20 min,固体平板和斜面培养基均在上述培养基中加入1.8%的琼脂粉。
1.2 方法
1.2.1 菌株Brevibacillus sp.XF-03的短程反硝化驯化 将本实验室保藏的菌株Brevibacillussp.XF-03接种至装有LB培养液的三角瓶中,30℃,160 r/min摇床振荡活化2代,并将次级培养物活化至细菌对数生长期,作为接种体使用。取活化后的菌液10 mL,接入装有100 mL培养液的三角瓶中,摇床恒温振荡。以琥珀酸钠作为唯一碳源和能源,利用NO2
--N作为唯一氮源,采用梯度驯化的方法首先对细菌进行升温培养。驯化培养过程按10%梯度接种的方法,温度梯为:30℃→35℃→40℃→43℃→46℃→48℃ →50℃。菌株在50℃下生长稳定后,再对其进行亚硝态氮浓度梯度驯化。亚硝态氮浓度梯度增加依次为:50、100、150、 200 、300、400、500、600、700、800、1 000 mg/L。同时,在驯化的过程中定期监测水质并观察微生物的生长情况。
1.2.2 菌株Brevibacillussp.XF-03短程反硝化条件优化 将处于对数生长期的菌液接入100 mL短程反硝化培养基中,通过调节碳源(葡萄糖、苯酚、酒石酸钾钠、柠檬酸钠、乙醇、琥珀酸钠)和 C/N(3∶1,6∶1,9∶1,12∶1,15∶1,20∶1),50℃、160 r/min摇床振荡培养,定期测定培养液中菌体生长量与NO2
--N含量。考察碳源与碳氧化(C/N)对菌株Brevibacillussp.XF-03生长及短程反硝化能力的影响,每组试验设3个重复。
1.2.3 短程反硝化动力学研究 在最佳试验条件下,应用Haldane方程对菌株生长和亚硝态氮降解特性进行动力学研究[10]。为用回归曲线求出准确的动力学方程,动力学试验在较宽的NO2--N含量范围内进行,即0-1000 mg/L。NO2--N浓度选择如下:0-200 mg/L之间,每40 mg/L一个浓度梯度;200-600 mg/L之间,每100 mg/L一个浓度梯度;600-1000 mg/L之间,每200 mg/L一个浓度梯度。将菌悬液分别接种到装有上述不同初始NO2--N浓度的短程反硝化培养基中,摇瓶振荡培养,每隔适宜时间取样检测培养液中的菌体含量和残余NO2--N含量。
1.2.4 粗酶液的提取 将处于不同生理时期的菌液离心(8 000 r/min,10 min),弃去上清液,收集菌体,用0.01 Na2HPO4-NaH2PO4缓冲液(pH7.2)清洗后重悬细胞,再次离心,重复3次,留取菌体沉淀。按原菌液体体积的1/5-1/10加入裂解液,制成悬浮菌液,裂解液的成分为:pH8.0的50 mmol/L Tris-HCI缓冲液、2 mmol/L EDTA、100 mmol/L NaCl,置于冰水浴中超声破碎。破碎液于12 000 r/min,4℃离心20 min,收集上清液即为粗酶[11],低温保存待用。
1.2.5 菌体OD600测定 可见分光光度法,细胞干重采用干燥恒重法测量,根据细胞干重标准曲线将吸光度转换为细胞干重[12];亚硝态氮测定:N-(1-萘基)-乙二胺光度法[13];亚硝酸盐还原酶(NIR)活力测定:测定系统总体系3 mL,内含1.0 mL Na2HPO4-NaH2PO4缓冲液(pH7.2),1 mL粗酶液以及1 mL 0.01 mol/L的亚硝酸钠溶液,试剂全部加入后立即取适量上述混合液,测340 nm处的吸光度,剩余待测液在常温静置20 min后测其340 nm处的吸光度[14]。定义亚硝酸盐还原酶(NIR)活力单位(U)为每分钟转化1 μmol亚硝酸盐氮所需酶量。总蛋白含量用Bradford法测定[15],酶的比活力以每毫克蛋白质中所含酶的活力单位数计算。试验数据分析借助Matlab软件。
2 结果
2.1 碳源对菌株Brevibacillus sp.XF-03短程反硝化影响
图1为不同碳源情况下,42 h后菌株各自生长及NO-
2-N降解情况。由图1可以看出菌株Brevibacillussp.XF-03短程反硝化的顺利进行对碳源有一定的选择性。该菌几乎不能以乙醇、苯酚、酒石酸钾钠作为唯一碳源生长;而对葡萄糖而言,菌体在其中所需的适应阶段较长,亚硝态氮降解也不充分,可能是由糖类物质降解过程的复杂性引起的。分别以柠檬酸钠和琥珀酸钠为碳源时,菌株Brevibacillussp.XF-03均能较好地生长,与以柠檬酸钠为碳源相比,当以琥珀酸钠为碳源时,亚硝态氮降解率较高42 h亚硝态氮由100 mg/L降解到5.9 mg/L,降解率达94.1%,说明琥珀酸钠为该菌株的最适碳源。
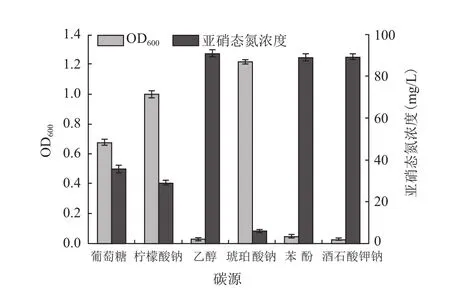
图1 碳源对菌体生长及NO-2-N降解的影响
2.2 碳氮化对菌株Brevibacillus sp.XF-03短程反硝化影响
以琥珀酸钠为唯一碳源,亚硝酸钠为唯一氮源,固定碳源含量,通过调节亚硝酸钠含量控制碳氮比。由图2可知,当碳氮化(C/N)小于12时,亚硝态氮去除率及菌株生长情况随着C/N增加而提高,C/N大于12时,亚硝态氮去除率及菌株生长情况趋于平缓,提高不明显。综合考虑,确定菌株Brevibacillussp.XF-03的最适碳氮比为12∶1。
2.3 菌株Brevibacillus sp.XF-03短程反硝化特性
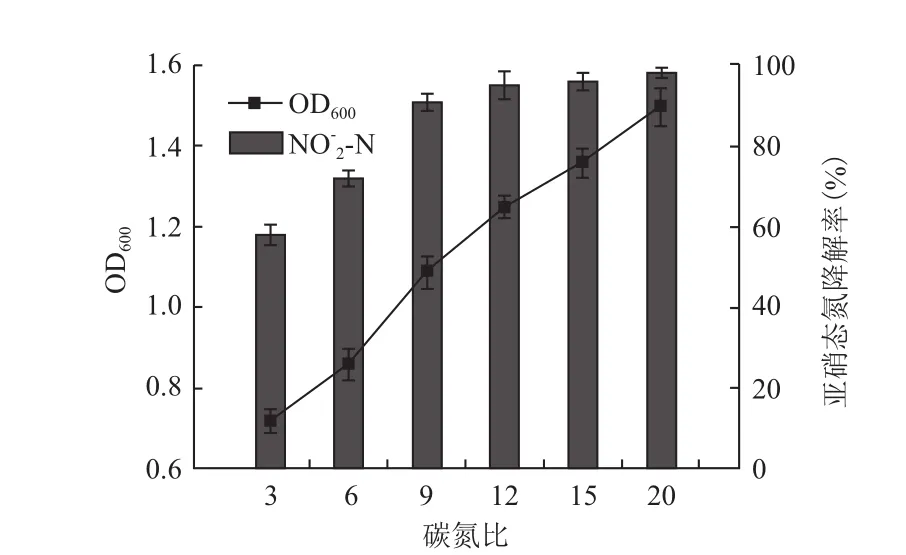
图2 碳氮比(C/N)对菌体生长及NO2--N降解的影响
在最适碳源及碳氮比,培养液初始NO-2-N浓度为100 mg/L条件下,菌株Brevibacillussp.XF-03生长及亚硝酸盐降解曲线如图3。经18 h的适应期后菌株Brevibacillussp.XF-03进入对数生长期,在该时期内,短程反硝化作用明显加强,因为这个时期(18-36 h)是细菌生长和繁殖最旺盛的时期,细胞合成所需要的能量和还原力主要在这一阶段被消耗,因此反硝化作用主要在这一时段内完成。培养42 h,NO-
2-N从100 mg/L降 至4.90 mg/L, 去 除 率 为95.1%。在对数期后期,有效细菌的数目虽逐渐减少,而亚硝酸盐仍有少量降低,此时很可能是细菌的同化起主要作用。

图3 菌株Brevibacillus sp.XF-03生长曲线及亚硝态氮变化
2.4 亚硝态氮降解动力学
短程反硝化菌在以亚硝酸盐氮为底物进行反硝化的过程中,亚硝态氮对微生物有一定的毒害作用,当其浓度达到一定值时,也会对反硝化过程产生抑制,导致反应速率下降,因此建立动力学模型时,需要选择一种非竞争性底物抑制模型,已经有多种底物抑制模型被建立,其中Haldane模型是最常见的模型,定义如下式:
μ=μmaxC/(Ks+C+C2/Ki) (1)
式中,μ为底物比降解速率(h-1);μmax为最大比降解速率(h-1);C为底物初始浓度(mg/L);KS
为半饱和常数(mg/L);Ki为底物抑制常数(mg/L)。
试验假定亚硝态氮是菌株Brevibacillussp.XF-03比生长速率的单一限制性底物,摇瓶中的溶氧为常数且为非限制性因素。对于每一个初始亚硝态氮浓度(C),比生长速率(μ) 由指数生长期决定。对每一个摇瓶指数生长期的菌体浓度和时间的半对数图做线性最小二乘拟合得,在指数生长期,μ 约为一个常数。
运用Matlab软件处理试验数据,得到亚硝酸盐抑制动力学方程式(2)。模型相关性系数R2为0. 992,说明拟合曲线与试验实测值相关性良好。
μ=1.28C/(451.42+ C+C2/176.77) (2)
模拟曲线如图4所示。底物比降解速率(μ)随着亚硝态氮初始浓度的增加呈现先增大后减小的趋势,这是典型的抑制生长模式,最大比降解速率发生在亚硝态氮浓度为282.48 mg/L处,随着初始亚硝态氮浓度的继续增加,其比降解速率逐渐减小,在这一过程中,亚硝态氮抑制作用占主导。此外,反应体系中代谢物的积累也是造成亚硝态氮比降解速率下降的原因之一。

图4 亚硝酸盐氮降解动力学拟合曲线
2.5 酶活性的测定
以琥珀酸钠为碳源,C/N=12,初始亚硝态氮浓度为282.48 mg/L,研究菌株Brevibacillussp.XF-03亚硝酸盐还原酶活性。由图5可知,菌株Brevibacillussp.XF-03在生长停滞期,亚硝酸盐还原酶活性较低,亚硝态氮降解率仅有12.56%,在其生长指数期的后期,产生大量亚硝酸盐还原酶,且亚硝酸盐还原酶的比活力达到最大值0.279(U/mg protein),亚硝态氮降解速率提高,说明脱氮过程中菌株不同生长阶段亚硝态氮还原酶的活力不同,亚硝酸盐还原酶是短程反硝化菌降解亚硝态氮的关键酶。
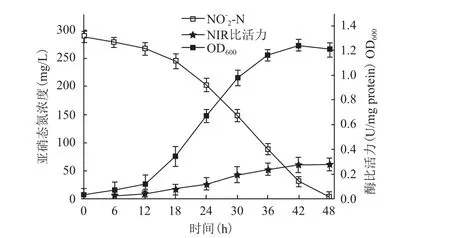
图5 菌体生长,亚硝态氮降解及亚硝态氮还原酶比活力变化曲线
3 讨论
反硝化微生物大多数需要碳源作为电子供体,具有较广泛的碳源谱,碳源的种类影响到反硝化强度[16]。Martienssen等[17]认为有机碳源的数量和种类是影响反硝化微生物菌落结构的重要因素之一,这与本研究结论一致。同时,研究表明碳氮比也是影响微生物脱氮效果的重要因素之一,低碳氮比条件下,NO-2和N2O均可利用内碳源进行反硝化,氧化亚氮还原酶竞争电子的能力较弱,反硝化过程出现N2O积累。碳源充足,亚硝态氮还原酶和氧化亚氮还原酶同时利用外碳源进行反硝化,初始阶段产生的N2O在接下来的反硝化过程中被迅速还原为N2[18]。碳源不足,反硝化速率降低;碳源过量虽可提高系统反硝化脱氮能力,但增加了碳源消耗,同时也增加了耗氧量。
目前,已发现多种反硝化菌,主要分布于Pseudomonas、Alcaligenes和Bacillus三个属[19]。对反硝化微生物的研究主要集中于以硝酸盐氮为电子受体的反硝化菌,对亚硝酸型反硝化菌的研究集中在混合菌反应器运行上[20,21],而对在温度与亚硝态氮含量均较高的条件下短程反硝化单菌种的研究鲜见报道。亚硝酸型反硝化是一个复杂生化过程,是短程硝化反硝化的重要过程。本研究采用梯度驯化得到高效的短程反硝化菌,优化影响短程反硝化的环境因子,构建亚硝酸盐氮降解动力学模型及初步探讨其酶活,以期为短程硝化反硝化工艺的应用以及传统反硝化过程中亚氮积累问题的解决提供技术参数。同时短程反硝化微生物的特性、亚硝酸盐还原酶的
表达及其应用有待于进一步研究。
4 结论
驯化得到高温短程反硝化菌Brevibacillussp.XF-03,该菌株最佳生长碳源为琥珀酸钠,碳氮比(C/N)为12。
动力学模型具有良好的有效性,相关系数(R2)为0.992。
亚硝酸盐还原酶是短程反硝化菌降解亚硝态氮的关键酶,在菌株Brevibacillussp.XF-03生长指数期的后期,产生大量亚硝酸盐还原酶,且亚硝酸盐还原酶的比活力达到最大值0.279(U/mg protein)。
[1] Walter KD· Availability, uptake and regeneration of phosphate in mesocosms with varied levels of P deficiency[J]. Hydrobiologia, 1995, 297:1-9.
[2] 周康群, 刘晖, 崔英德.一株新的厌氧除磷功能菌株的鉴定与活性[J].化工学报, 2008, 59(6);1522-1530.
[3] Potter TG, Tseng CC, Koopman B. Nitrogen removal in partial nitrification/complete denitrification process[J].Water Environment Research, 1998, 70(3);334-342.
[4] Ladan H, George N, Amarjeet SB, et al. Long term performance of MBR for biological nitrogen removal from synthetic municipal wastewater[J].Chemosphere, 2007, 66(5);849-857.
[5] 陈英文, 陈徉.膜生物反应器同步硝化反硝化系统的研究[J].环境工程学报, 2008, 2(7);902-905.
[6] Ruiz G, Jeison D, Rubilar O, et al. Nitrification-denitrification via nitrite accumulation for nitrogen removal from wastewaters[J]. Bioresource Technology, 2006, 97(2);330-335.
[7] Turk O, Mavinic DS. Maintaining nitrite build-up in a system acclimated to free ammonia[J]. Water Research, 1989, 23(11):1383-138.
[8] 尚会来, 彭永臻, 张静蓉, 等.温度对短程硝化反硝化的影响[J].环境科学学报, 2009, 29(3);516-520.
[9] 刘超翔, 胡洪营, 彭党聪.短程硝化反硝化工艺处理焦化高氨废水[J].中国给水排水, 2003, 19(8);11-14.
[10] 戚以政, 汪叔雄. 生化反应动力学与反应器[M]. 北京:化学工业出版社, 1992
[11] Dong XJ, Qing H, He LY, Xin J. Characterization of phenoldegrading bacterial strains isolated from natural soil[J]. International Biodeterioration & Biodegradation, 2008(3);62:257-262.
[12] Jiang Y, Wen JP, Bai J. Biodegradation of phenol at high initial concentration by Alcaligenes faecalis[J].Journal of Hazardous Materials, 2007, 147(1-2);672-676.
[13] 国家环保总局.水和废水监测分析方法. [M].第4版, 北京:中国环境科学出版社, 2002.
[14] Padhi SK, Tripathy S, Sen R, et al. Characterisation of heterotrophic nitrifying and aerobic denitrifying Klebsiella pneumoniae CF-S9 strain for bioremediation of wastewater[J]. International Biodeterioration & Biodegradation, 2013, 78;67-73.
[15] Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding[J]. Anal Biochem, 1976, 72(1);248-254.
[16] Blaszczyk M. Effect of medium composition on the denitrification of nitrate byParacoccus denitrificans[J].Applied Environmental Microbiology, 1993, 59(11);3951-3953.
[17] Martienssen M, Schops R. Population dynamics of denitrifying bacteria in a model biocommunity[J].Water Research, 1999, 33(3);639-646.
[18] 巩有奎, 王淑莹, 王莎莎, 等.碳氮比对短程反硝化过程中N2O产生的影响[J].化工学报, 2011, 62(7);2049-2054.
[19] Cheneb YD, Philoppot L, Hartmann A, et al. 16S rDNA analysis for characterization of denitrifying bacteria isolated from three agricultural soils[J].FEMS Microbiology Ecology, 2000, 34(2);121-128.
[20] 高景峰, 彭永臻, 王淑莹.温度对亚硝酸型硝化/反硝化的影响[J].高技术通讯, 2002, 12:88-93.
[21] 巩有奎, 王淑莹, 王莎莎, 等. DO 对短程反硝化过程中 N2O产量的影响[J].中南大学学报:自然科学版, 2012, 43(1);395-400.
(责任编辑 李楠)
Characteristics and Degradation Kinetics of Thermophilic Shotcut Denitrifier Brevibacillus sp.XF-03
Hao Minna Yang Yunlong Ge Qilong
(College of Environmental Science and Engineering,Taiyuan University of Technology,Taiyuan 030024)
The denitrifier Brevibacillus sp. XF-03 could reduce high nitrite concentration(1 000 mg/L)under the condition of high temperature(50℃)by the method of gradient domestication. The effects of carbon source and C/N on growth and denitrification of the denitrifier were optimized by single factor experiments. The results showed that the optimum shortcut denitrification carbon source of Brevibacillus sp. XF-03 was sodium succinate, optimum C/N ratio 12∶1. Under the optimal condition, the nitrite removal efficiency reached 95.1% within 42 hours when the original concentration of NO-2-N was 100 mg/L. Nitrite degradation kinetic studies indicated that the strain followed Haldane’s model, and the parameters were:μmax(maximum specific rate)=1.28 h-1, Ks(half-satruration constant)=451.42 mg/L, Ki(inhibition constant)= 176.77 mg/L. Nitrite reductase(NIR)activity was preliminary discussed. The NIR specific activity reached 0.279(U/mg protein)at the lag phase of exponential growth.
Brevibacillus sp.XF-03 High temperature Shortcut denitrification Nitrite degradation kinetics Enzyme activity
2013-07-29
郝敏娜,女,硕士研究生,研究方向:水污染控制;E-mail:tyuthaominna@163.com
猜你喜欢
中国民间疗法(2021年18期)2021-11-02
中国民间疗法(2021年9期)2021-07-22
昆明医科大学学报(2021年5期)2021-07-22
浙江工业大学学报(2017年5期)2018-01-22
国外医药(抗生素分册)(2016年3期)2016-07-12
环境污染与防治(2016年12期)2016-03-13
中国洗涤用品工业(2015年9期)2015-02-28
中国酿造(2014年9期)2014-03-11
中国当代医药(2012年16期)2012-01-23
植物营养与肥料学报(2011年4期)2011-10-26
