
肉鸡体重对净能评定中总产热量和绝食产热量的影响
2014-03-14 03:40:42蔡辉益闫海洁刘国华杨华明
动物营养学报 2014年8期
刘 伟 蔡辉益* 闫海洁 刘国华 张 姝 杨华明
(1.中国农业科学院饲料研究所,农业部饲料生物技术重点开放实验室,北京 100081;2.吉林省农业科学院,长春 130033)
家禽饲料原料能值的评定以代谢能(metabolizable energy,ME)为主,ME包含了家禽对饲料摄食和消化过程产生的热增耗(heat increment,HI)。所以,ME并不能真实的代表可被家禽用来维持和生产利用的能量[1]。净能体系考虑了HI的损失,在评定饲料原料能值上更加准确。其中,HI是通过计算动物采食前后产热量的差值来估测的。家禽产热量受品种、年龄、性别、采食量、环境等多种因素影响。O’Neill等[2]研究表明,母鸡的绝食产热量高于公鸡。Johnson等[3]将蛋鸡的采食量分成9个梯度,结果表明产热量随着采食量的减少明显下降。王旭莉[4]研究表明,蛋鸡的产热量与采食量和产蛋率相关。Waring等[5]、O’Neill等[6]、Valencia 等[7]报道环境温度影响家禽产热量。Li等[8]报道,光照时间影响产热量。家禽的绝食产热量受绝食时间长短的影响,Shannon等[9]、O’Neill等[6]、Lundy 等[10]、Macleod 等[11]测定家禽绝食48 h后的产热量;宁冬等[12]在测定棉籽粕和玉米蛋白粉对蛋鸡的净能值中,采用蛋鸡绝食3 d后的产热量作为绝食产热量。
Close等[13]、胡琴[14]研究发现,猪的产热量与体重和绝食时间有关。而体重对肉鸡总产热量、绝食产热量以及绝食时间影响的报道较少。本研究利用禽用开放式呼吸测热装置测定4~7周龄阶段爱拔益加(AA)肉鸡不同周龄体重下的总产热量和绝食产热量,研究体重对其采食和绝食期间的耗氧量、二氧化碳生成量、呼吸熵以及总产热量的影响,研究并确定4~7周龄阶段AA肉鸡适宜的绝食时间和代谢体重绝食产热量,为以后测定肉鸡饲料原料净能值建立方法。
1 材料与方法
1.1 试验动物与试验设计
试验采用单因子试验设计,选用4~7周龄阶段AA肉鸡48只,每周第1天选取12只体重相近的肉鸡作为1组,分为 A、B、C、D 4组,每组6个重复,每个重复2只鸡,全部为公鸡。采食前和采食3 d后不同周龄阶段肉鸡之间体重差异显著(P<0.05),见表1。每周将1组肉鸡随机放入6个呼吸测热室中。第1天适应小室环境,从第2天开始连续测定3 d采食过程的总产热量。第5天09:00称量各个重复肉鸡体重后开始绝食,并连续测定72 h绝食产热量,以间隔12 h作为1个统计单位。

表1 AA肉鸡体重Table 1 The body weight of AA broilers
1.2 试验饲粮
试验饲粮采用玉米-豆粕型,营养水平参考《鸡的饲养标准》(2004),试验饲粮组成及营养水平见表2。
1.3 饲养管理
呼吸测热装置为1台气体数据采集仪和1台气体数据分析仪控制6个小室,分析仪为Qtfxy-6多功能气体分析仪,气体数据采集仪和气体数据记录分析软件型号分别为MTAC-A02和PDRLAB-12A,由中国农业科学院饲料研究所和吉林省农业科学院共同研制,委托哈尔滨物格电子技术有限公司生产。每周第2天和第5天09:00对肉鸡称重。小室温度控制在21℃左右,湿度65%左右。采食期间自由采食。整个测定期间自由饮水。
1.4 数据采集
电脑软件每3 m in自动记录每个小室2只鸡的耗氧量、二氧化碳生成量以及呼吸熵。
1.5 计算公式
1.5.1 耗氧量和二氧化碳生成量

式中:CO2,1和 CO2,2分别为户外和小室内氧气浓度;CCO2,2和 CCO2,1分别为小室内和户外二氧化碳浓度;V为进气量和排气量。
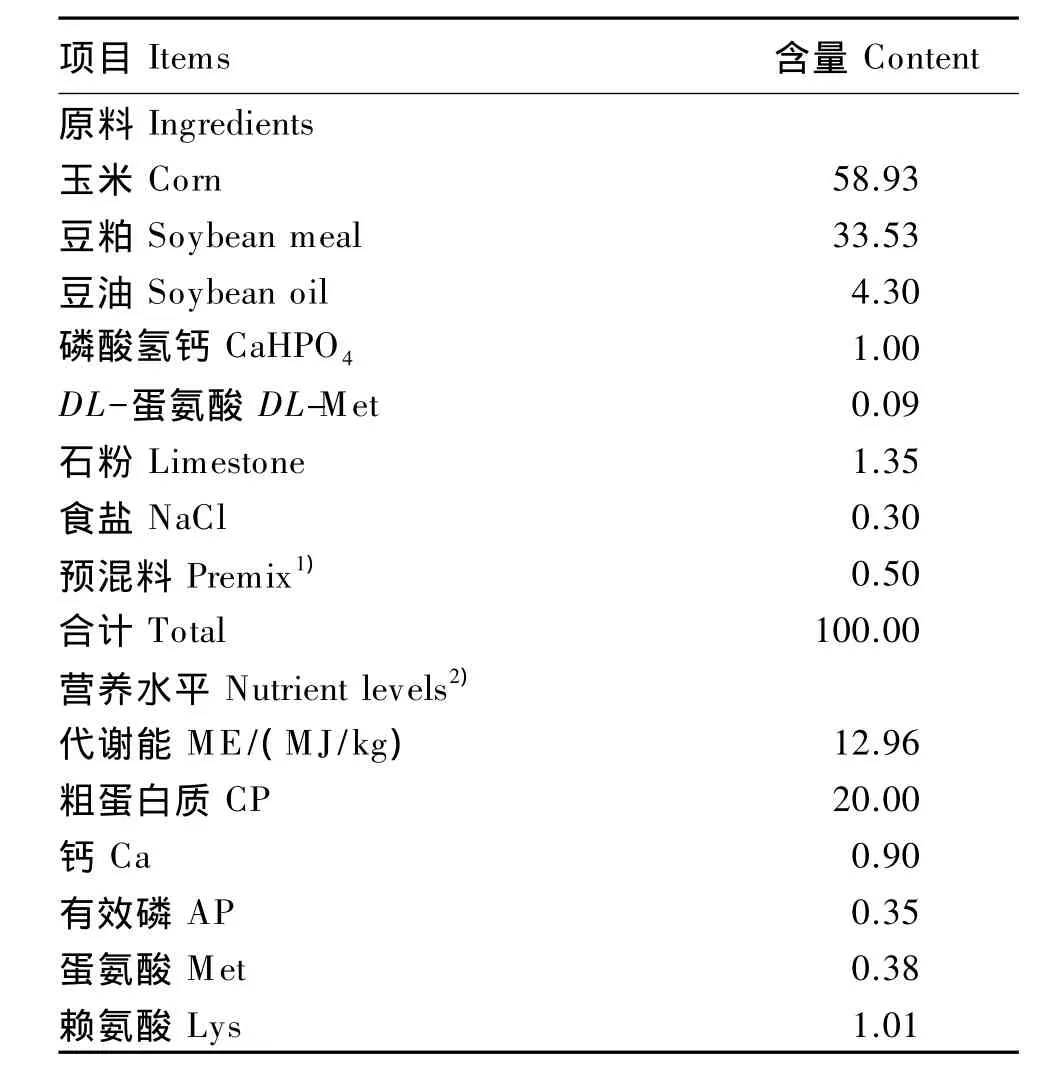
表2 试验饲粮组成及营养水平(风干基础)Table 2 Composition and nutrient levels of the experimental diet(air-dry basis) %
1.5.2 产热量
根据 Brouwer[15]的计算公式:
产热量(kJ)=16.175×VO2(L/d)+
5.021×VCO2(L/d);
呼吸熵(RQ)=VCO2(L/m in)/VO2(L/m in);代谢体重绝食产热量[FHP,MJ/(kg BWb·d)]=
aBWb。
式中:VO2和VCO2分别为耗氧量和二氧化碳生成量;a为每千克代谢体重绝食产热量;BW为体重;b为代谢体重指数。
1.6 数据统计分析
试验数据用平均值±标准差表示,采用SPSS 19.0软件中的ANOVA过程进行单因素方差分析,差异显著者采用Duncan氏法进行多重比较,P<0.05为差异显著水平。
2 结果与分析
2.1 绝食时间对不同体重肉鸡耗氧量、二氧化碳生成量和呼吸熵的影响
由表3和表4可知,除A和B组肉鸡在绝食到60 h时的耗氧量和二氧化碳生成量低于绝食到72 h时以外,不同体重肉鸡随绝食时间的延长,耗氧量和二氧化碳生成量逐渐降低。A和C组肉鸡耗氧量和二氧化碳生成量在绝食到12和24 h时差异不显著(P>0.05),与绝食到36 h时差异显著(P<0.05)。A组肉鸡绝食到48和72 h时的耗氧量和二氧化碳生成量差异不显著(P>0.05),绝食到60 h显著低于绝食到72 h(P<0.05)。B组肉鸡从绝食12到36 h的耗氧量和二氧化碳生成量显著降低(P<0.05),绝食到 36、48和 72 h时差异不显著(P>0.05),绝食到60 h时的耗氧量和二氧化碳生成量低于绝食到72 h,差异不显著(P>0.05),但显著低于绝食到 36 和 48 h(P<0.05)。C组肉鸡绝食到48、60和72 h时的耗氧量和二氧化碳生成量逐渐降低,但差异不显著(P>0.05)。D组肉鸡从绝食12到48 h的耗氧量和二氧化碳生成量显著降低(P<0.05),进一步绝食到72 h,耗氧量和二氧化碳生成量趋于平稳,无显著差异(P>0.05)。整体来看,各组肉鸡绝食到48 h以后的耗氧量和二氧化碳生成量逐渐趋于平稳。
由图1可见,从绝食12到72 h,不同体重肉鸡的呼吸熵处于0.65~0.75之间。随着绝食时间的延长,呼吸熵呈逐渐下降的趋势,但无明显规律。相同绝食时间下不同体重肉鸡之间呼吸熵也无明显规律。
2.2绝食时间对不同体重肉鸡绝食产热量的影响
由表5可见,A和B组肉鸡绝食到36和48 h的产热量差异不显著(P>0.05)。A组肉鸡绝食到72 h的产热量低于绝食到48 h(P>0.05),显著低于绝食到36 h(P<0.05),显著高于绝食到 60 h(P<0.05)。B组肉鸡绝食到72 h的产热量高于绝食到60 h,低于绝食到36和48 h,差异均不显著(P>0.05)。C和D组肉鸡绝食到48、60和72 h时的产热量逐渐降低,但差异不显著(P>0.05)。A和B组肉鸡绝食到36 h以后产热量趋于稳定,与绝食到48 h的产热量相近。C和D组肉鸡绝食到48 h以后的产热量趋于稳定。
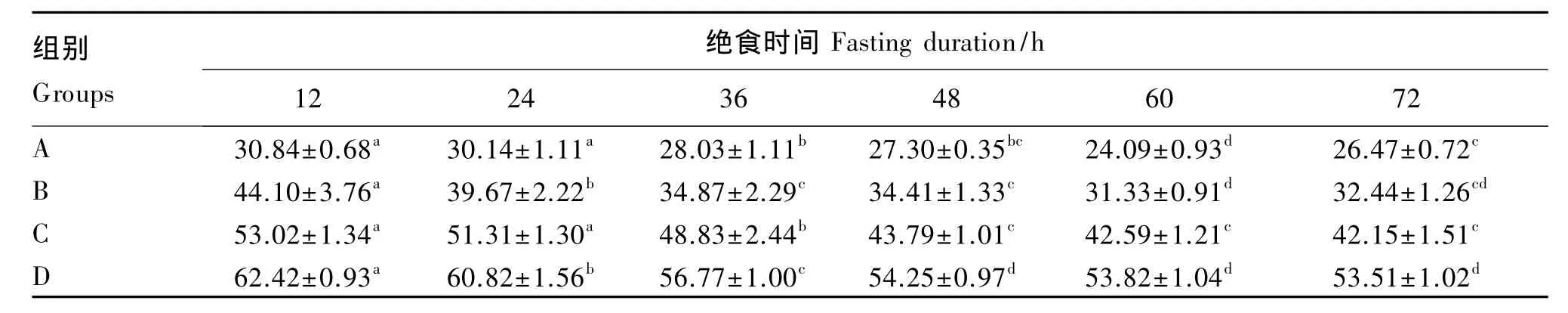
表3 绝食时间对不同体重肉鸡耗氧量的影响Table 3 Effects of fasting duration on O2 consumption of different body weight broilers L/d

表4 绝食时间对不同体重肉鸡二氧化碳生成量的影响Table 4 Effects of fasting duration on CO2 production of different body weight broilers L/d

图1 绝食时间对不同体重肉鸡呼吸熵的影响Fig.1 Effects of fasting duration on RQ of different body weight broilers
A、B、C和D组肉鸡绝食到48 h时的产热量都趋于稳定,以肉鸡采食3 d后的体重的对数为自变量,以A、B、C和D组肉鸡绝食到48 h时的产热量的对数作为因变量进行回归分析,得到回归方程:y=0.737x-0.332(R2=0.99,P<0.01)。由回归方程可知:lga=-0.332,得到4~7周龄阶段 AA 肉鸡代谢体重 绝 食 产 热 量:a=0.47 MJ/(kg BW0.74· d)(R2=0.99,P<0.01)。
2.3 体重对肉鸡耗氧量、二氧化碳生成量、总产热量、绝食产热量和呼吸熵的影响
由表6可知,4~7周龄阶段肉鸡采食3 d内的耗氧量、二氧化碳生成量和总产热量随着体重的增加显著提高(P<0.05)。从第4周到第7周,总产热量提高了92.94%(P<0.05)。体重对不同周龄肉鸡采食期间呼吸熵没有显著影响(P>0.05),呼吸熵均在0.90以上。
根据表5结果,将4~7周龄的肉鸡绝食到48 h的产热量作为各周龄阶段的绝食产热量。由表7可知,绝食到稳定水平时的耗氧量、二氧化碳生成量和产热量随绝食前体重的增加显著提高(P<0.05)。与第4周相比,第7周肉鸡产热量提高了 98.15%(P<0.05)。各周龄之间每日每千克代谢体重产热量差异不显著(P>0.05),与由回归方程得到的 0.47 M J/(kg BW0.74·d)相近。绝食到48 h时,各周龄肉鸡的呼吸熵差异不显著(P>0.05)。

表5 绝食时间对不同体重肉鸡绝食产热量的影响Table 5 Effects of fasting duration on FHP of different body weight broilers MJ/d
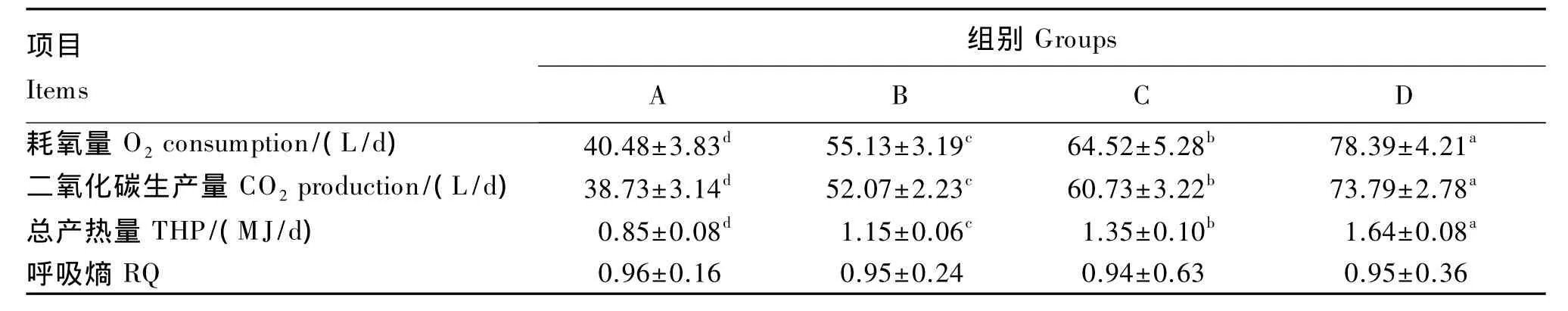
表6 体重对肉鸡耗氧量、二氧化碳生成量、总产热量及呼吸熵的影响Table 6 Effects of body weight on O2 consumption,CO2 production,THP and RQ of broilers
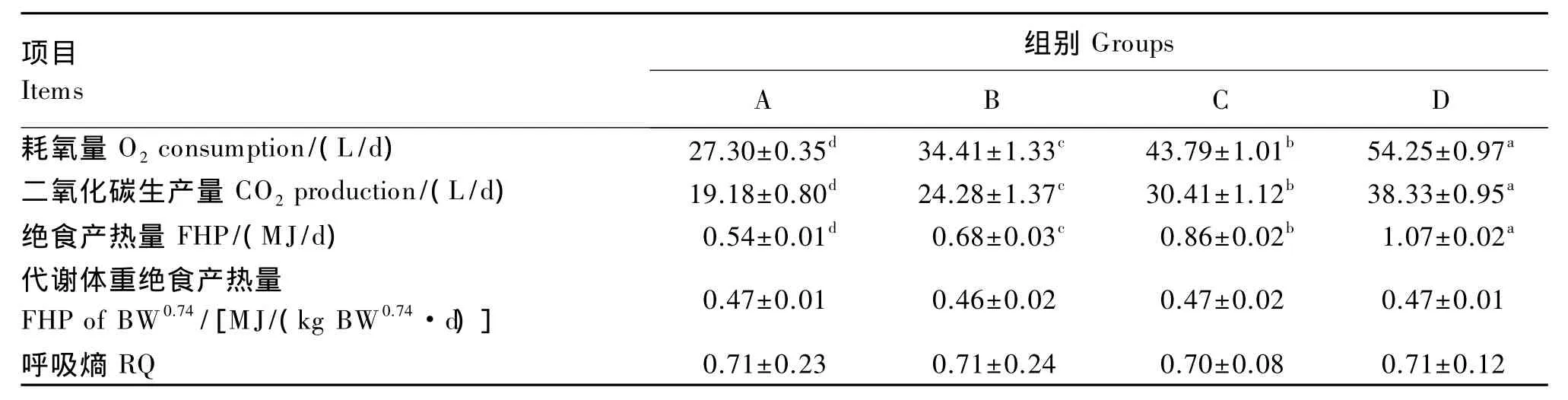
表7 体重对肉鸡耗氧量、二氧化碳生成量、绝食产热量、代谢体重产热量及呼吸熵的影响Table 7 Effects of body weight on O2 consumption,CO2 production,FHP,FHP of BW 0.74 and RQ of broilers
3 讨论
3.1 绝食时间对不同体重肉鸡耗氧量、二氧化碳生成量和呼吸熵的影响
动物采食饲粮后经过消化吸收营养物质进入体内,通过生物氧化还原反应过程进行能量代谢,实质上就是动物吸入氧气由肺进入血液进而被运送到各组织细胞参与营养物质的氧化还原反应,形成二氧化碳和水,二氧化碳经血液运输由肺排出。通过测定这种气体交换过程动物的耗氧量和二氧化碳生成量,可以间接计算出动物的产热量[16]。氧气和二氧化碳浓度的测定对传感器的准确度、精度和耐用性都有较高的要求。本试验所采用的粒子流氧分析仪和近红外二氧化碳分析仪的测定值可精确到万分之一。
本研究发现,4~7周龄阶段肉鸡耗氧量、二氧化碳生成量随绝食时间的延长而逐渐降低,只有第4周和第5周肉鸡绝食到60 h时的耗氧量、二氧化碳生成量略高于绝食到72 h,可能是由于4~5周龄阶段肉鸡体重较小,绝食到60 h体内营养物质消耗过大,晚间活动较少,较白天安静,呼吸强度低于白天。总体来看,4~7周龄阶段肉鸡绝食到48 h以后的耗氧量、二氧化碳生成量逐渐趋向平稳。
动物通过呼吸作用进行营养物质代谢时,所消耗的氧气与产生的二氧化碳之比称为呼吸熵。各种营养物质在体内氧化时,其呼吸熵均有各自特定的数值,是由各自的化学结构不同而决定的。葡萄糖氧化时呼吸熵为1.00,脂肪氧化时呼吸熵为0.70,体蛋白氧化时呼吸熵为0.809,由于动物体内各种营养物质是以不同的比例共同被氧化的,所以呼吸熵应该处于0.70~1.00之间。当动物绝食时,体内沉积的脂肪和蛋白质向碳水化合物转化合成,呼吸熵在0.70以下[16]。本研究中 4~7 周龄阶段肉鸡绝食12~72 h期间呼吸熵处于0.65~0.75之间,表明肉鸡在绝食代谢状态下,维持机体体温和基础代谢所需的能量主要来源于对体内沉积的脂肪和蛋白质的降解转化。
3.2 绝食时间对不同体重肉鸡绝食产热量的影响
Close等[13]测定 20~45 kg长白猪连续绝食4 d的产热量,发现产热量在绝食第1天迅速降低,在随后的3 d,产热量降低的速度减慢,第3天和第4天产热量差异不显著,并在第3天或第4天得到稳定的最小绝食产热量。胡琴[14]分别测定了3个体重阶段的健康去势公猪(杜×长×大)绝食96 h的产热量,发现不同体重猪绝食产热量达到稳定水平所需的绝食时间不同,体重越大,绝食产热量趋于平稳时所需的时间越长。其原因在于不同体重阶段的猪采食后的排空所需时间不同,即体重越大排空时间越长,营养物质在机体存留的时间越长,由饲粮引起的产热量降低的速度越慢,产热量达到稳定时所需的时间越长。Li等[8]研究表明,蛋鸡绝食到22和46 h时的产热量没有差异,绝食产热量达到稳定水平所需的绝食时间与绝食前采食量有关,采食量越少绝食产热量达到稳定的时间越快,因采食进入体内的营养物质引起的代谢产热在绝食到22 h以后就可能已经消失。本研究中,4~7周龄阶段肉鸡绝食到48 h以后产热量逐渐趋于平稳,因此,可将48 h作为4~7周龄阶段肉鸡最佳的绝食时间。这与 Shannon等[9]、O’Neill等[6]、Lundy 等[10]、Macleod 等[11]在家禽上所得的结果相同。
通常以单位代谢体重BW0.75表示动物的绝食代谢产热。对于动物整个生长阶段产热量,Tess等[17]、Van M ilgen 等[18]认 为 0.75 过 大。Noblet等[19]和胡琴[14]分别计算 45~150 kg 和 30~100 kg猪绝食产热量,得到适宜的系数分别为0.42和0.55。本试验通过计算4~7周龄阶段不同体重肉鸡的绝食产热量,得到适宜的系数为0.74,与0.75相近。
3.3 体重对采食代谢和绝食代谢的影响
经过遗传选育,AA肉鸡具有生长快速、饲粮转化率高等特点,每周体重都有较大变化。本研究发现4~7周龄阶段肉鸡每周采食过程的总产热量都随不同周龄体重的增加而显著升高,这主要因为体重增加,产热体表面积增加。此外,随着体重增加,肉鸡消化道体积变大,采食饲粮增多,营养物质在机体内的代谢产热增加。
胡琴[14]测定 35.6~95.6 kg 猪 7 个体重阶段稳定的绝食产热量发现,随体重增加,绝食产热量显著增加。本试验中,以48 h作为4~7周龄阶段肉鸡稳定的绝食时间,不同周龄肉鸡绝食到48 h时的耗氧量、二氧化碳生成量、产热量也随体重的增加显著升高。因此,在测定肉鸡产热量时要根据肉鸡体重分阶段进行测定。以0.74为代谢体重指数系数,将4~7周龄阶段肉鸡绝食到48 h的产热量计算为代谢体重产热量,经统计分析,不同周龄肉鸡绝食到48 h的代谢体重产热量无显著差异,与根据回归方程得到的数值0.47 MJ/(kg BW0.74·d)(R2=0.99,P< 0.01)接近。故认为在本试验条件下,可将0.74作为代谢体重系数用于计算4~7周龄阶段肉鸡绝食产热量。
4 结论
4~7周龄阶段AA肉鸡采食代谢和绝食代谢受体重影响显著。肉鸡生长后期总产热量和绝食产热量应该按周龄分别计算。4~7周龄阶段肉鸡绝食产热量在绝食48 h以后达到稳定,可将48 h作为最佳绝食时间。本试验条件下,1.21~3.10 kg肉鸡单位代谢体重系数为0.74,单位代谢体重绝食产热量为 0.47 M J/(kg BW0.74·d)。
[1] MACLEOD M G,THEODOROU M K,FRANCE J.Modelling the utilization of dietary energy and am ino acids by poultry[M]//THEODOROU M K,FRANCE J.Feeding systems and feed evaluationmodels.Wallingford:CAB International,1999:393-412.
[2] O’NEILL S JB,JACKSON N.The heat production of hens and cockerels maintained for an extended period of time at a constant environmental temperature of 23 C[J].The Journal of Agricultural Science,1974,82(2):549-552.
[3] JOHNSON R J,FARRELL D J.Energy metabolism of groups of broiler breeders in open-circuit respiration chambers[J].British Poultry Science,1983,24(4):439-453.
[4] 王旭莉.蛋鸡玉米和豆粕净能值的测定及其净能体系的应用[D].硕士学位论文.杨凌:西北农林科技大学,2010.
[5] WARING J J,BROWN W O.Calorimetric studies on the utilization of dietary energy by the laying White Leghorn hen in relation to plane of nutrition and environmental temperature[J].The Journal of Agricultural Science,1967,68(1):149-155.
[6] O’NEILL S J B,BALNAVE D,JACKSON N.The influence of feathering and environmental temperature on the heat production and efficiency of utilization of metabolizable energy by the mature cockerel[J].The Journal of Agricultural Science,1971,77(2):293-305.
[7] VALENCIA M E,MAIORINO PM,REID B L.Energy utilization in laying hens Ⅲ.Effect of dietary protein level at21 and 32 C[J].Poultry Science,1980,59(11):2508-2513.
[8] LIY,ITO T,YAMAMOTO S.Use of lim ited daily access to food in measuring the heat production associated w ith food intake in laying hens[J].British Poultry Science,1991,32(4):829-839.
[9] SHANNON D W F,BROWN W O.Calorimetric studies on the effect of dietary energy source and environmental temperature on the metabolic efficiency of energy utilization by mature Light Sussex cockerels[J].The Journal of Agricultural Science,1969,72(3):479-489.
[10] LUNDY H,MACLEOD M G,JEW ITT T R.An automated multi-calorimeter system:prelim inary experiments on laying hens[J].British Poultry Science,1978,19(2):173-186.
[11] MACLEOD M G.Effects of feeding by crop intubation on energy metabolism and physical activity in domestic cockerels[J].British Poultry Science,1991,32(5):1089-1095.
[12] 宁冬,呙于明,王永伟,等.间接测热法和回归法估测棉籽粕和玉米蛋白粉在蛋鸡中的代谢能和净能值[J].动物营养学报,2013,25(5):968-977.
[13] CLOSEW H,MOUNT L E.The rate of heat loss during fasting in the grow ing pig[J].British Journal of Nutrition,1975,34(2):279-290.
[14] 胡琴.生长肥育猪玉米和豆粕净能值的测定[D].博士学位论文.北京:中国农业大学,2012.
[15] BROUWER E.Report of sub-comm ittee on constants and factors[C]//BLAXTER K L.Energy metabolism.London:Academ ic Press,1965:441-443.
[16] 杨嘉实,冯仰廉.畜禽能量代谢[M].北京:中国农业出版社,2002:31-33.
[17] TESSM W,DICKERSON G E,NIENABER JA,et al.The effects of body composition on fasting heat production in pigs[J].Journal of Animal Science,1984,58(1):99-110.
[18] VAN M ILGEN J,BERNIER J F,LECOZLER Y,et al.Major determ inants of fasting heat production and energetic cost of activity in grow ing pigs of different body weight and breed/castration combination[J].British Journal of Nutrition,1998,79(6):509-517.
[19] NOBLET J,SHI X S,DUBOIS S.Effect of body weight on net energy value of feeds for grow ing pigs[J].Journal of Animal Science,1994,72(3):648-657.
猜你喜欢
动物营养学报(2022年11期)2022-12-08 08:23:14
特产研究(2021年6期)2021-12-08 03:16:38
河南农业科学(2019年9期)2019-09-24 11:49:32
医药前沿(2018年18期)2018-06-21 01:17:08
环球时报(2018-01-09)2018-01-09 06:56:11
海峡科学(2017年4期)2017-07-12 17:50:47
农村百事通(2016年6期)2016-05-14 00:27:38
红领巾·探索(2014年6期)2015-03-11 04:02:16
资源节约与环保(2015年8期)2015-01-27 22:38:27
铁路节能环保与安全卫生(2010年2期)2010-01-29 02:26:08
