
基于几何形态测量学的4种金线鱼矢耳石识别研究*
2014-03-09 03:40王学锋朱立新卢伙胜
海洋与湖沼 2014年3期
侯 刚 王学锋 朱立新 卢伙胜 冯 波①
(1.广东海洋大学 水产学院 湛江 524088;2.南海渔业资源监测与评估中心 湛江 524025;3.山东大学(威海) 海洋学院 威海 264209)
耳石由于具有代谢上的惰性,一旦沉积,材料很难被重新吸收或者改变,因而在鱼类生活史履历(通过元素指纹分析)和种类、种群识别(通过形态分析)研究中得到广泛应用(Campanaet al,1993)。由于矢耳石(sagitta)的形态分析应用效果要好于微耳石(lapillas)和星耳石(asteriseus),因而应用矢耳石进行相关研究的案例较多(叶振江等,2007;郭弘艺等,2007;王英俊等,2010)。在矢耳石形态分析中,目前主要存在两种类型方法,一类是以框架分析法(framework method)为代表的传统形态方法,通过线性测量等获得耳石的表征性变量,或获得面积指标,并结合一些耳石形状系数方面的信息(窦硕增等,2012);第二类型是几何形态测量学(geometric morphometrics)方法,又名新形态测量学,是20世纪90年代生物形态学研究领域中新发展起来的一种形态测量学方法,由于它在形态结构数值化和数据分析上具有重大突破,因而被称为形态测量学的革命(Rohlfet al,1993)。几何形态测量学主要有轮廓法(outline methods)和地标点法(landmark methods)两类方法,其中轮廓法,是对耳石二维投影后通过傅里叶分析提取表达轮廓信息的傅里叶系数(Campanaet al,1993),然后通过主成分分析和判别分析等数学统计方法区分鱼类种类或种群。一般来说,低谐次的傅里叶谐值在种类区分中作用较大(1—11),高谐次的傅里叶谐值在种群区分中作用较大(11—20)(Campanaet al,1993)。这种方法在耳石外部轮廓较为规则、轮廓变异不是很剧烈的鱼类研究案例中应用较多,但是如果耳石轮廓不规则,外形突起较多变异较大,或有所缺损,则可能会导致较大的分析偏差,而会表现一定的不适宜性。目前所讲的几何形态测量学方法,多侧重于地标点法,是基于笛卡尔地标的形状统计方法,通过获取二维影像上的地标点x、y坐标数据,将其进行相对扭曲(Relative Warp,RW)和薄板样条分析,绘制网格变形图,分析耳石形态变异。地标点方法由于将形态差异与变化定量描述和可视化,能够从数量和形状上来总结形态数据,且可以利用多变量技术来研究多维关系,具有目前其它形态学方法所无法达到的分析效果(Rohlfet al,1993),因而受到广泛关注,在鱼类的功能性形态学、生物形态学、形态进化学及系统分类学,以及考古学等方面得到广泛应用(Bookstein,1990;白明等,2007;闫宝荣等,2010)。
近年来国内学者开展了诸多耳石形态学的研究,较多侧重于传统形态学方法与傅里叶方法(张国华等,1999;潘晓哲等,2010;窦硕增等,2012),而关于地标点方法的应用研究相对较少。本研究以北部湾四种金线鱼属(Nemipterus,隶属于鲈形目(Perciformes)、金线鱼科(Nemipteridae))鱼类矢耳石为例,利用二维图像分析技术,研究几何形态测量学方法在四种金线鱼的耳石形态变异和耳石识别应用上的有效性,以期为进一步开展金线鱼属鱼类系统分类与进化,以及群体识别研究提供科学依据。
1 材料与方法
同一种类在不同地理海域,因栖息环境不同或者遗传因素,可能会产生鱼体或耳石形态在种群(population)、群体(stock)或更小进化尺度上的性状趋异(Klingenberget al,2003;Paulet al,2013)。为避免这种趋异引起的种间判断的潜在干扰,本研究中采用了四种金线鱼的小尺度同域性样本。红棘金线鱼(Nemipterus nemurus)采自拖网生产渔获,日本金线鱼(N.japonicus)采自流刺网生产渔获,金线鱼(N.virgatus)与深水金线鱼(N.bathybius)来自流刺网按站点渔业资源调查取样,基本情况见表1。样本按照海洋调查规范(GB12763.6)(国家技术监督局,2007)进行常规生物学测定后,摘取耳石,清洗、拭干后装入 2mL离心管。为减少体长效应引起的形态变化干扰,四种金线鱼耳石选取了性成熟样本的耳石(侯刚等,2012)。
1.1 形态特征测定
本研究采用左侧矢耳石为材料,耳石前期拍摄与处理参见文献(侯刚等,2012),耳石形态如图1所示。为描述矢耳石主要形态特征,选取测量了7个性状指标,分别为耳石长轴长、耳石短轴长、听沟周长、听沟面积、耳石周长、耳石面积、耳石最小外切矩形面积。耳石各性状名称与量度指标参照叶振江等(2007)和郭弘艺等(2007)的文献。
1.2 几何形态测量分析
1.2.1 地标点建立与提取 地标点在生物学应用上分为三大类(Bookstein,1990),第一型地标点(TypeⅠlandmark)是指不同组织间的交点,如肌肉与骨骼的接触点、鱼鳍与鱼体的接触点、昆虫翅脉的交点、树枝的分叉点等,能被精确定位,可能具有进化学上“同源”的意义;第二型地标点(Type Ⅱ landmark)是指结构中的凹陷或突起点,如牙齿或爪的尖端、骨头的突起或其它结构突出可以明确辨认的点,对测量有帮助,可以用来推论功能及作用;第三型地标点(Type Ⅲ landmark)是指结构的极点,如最长点、最宽点等,测量中常见,但取点有主观性,不见得可靠(Bookstein,1990;萧旭峰等,1998;冯波等,2011)。
根据操作性同源与信息含量的要求,本文选取了11个地标点,对其进行了定义:6、10、11点为Ⅰ型地标点,2、4、9为Ⅱ型地标点,1、3、5、7、8为Ⅲ型地标点(图2,表2)。地标点7由于在种类间和种间内变异较大,取点具有一定主观性,未定义为Ⅰ型地标点,而定义为Ⅲ型地标点。利用Tpsdig2软件在拍摄好的金线鱼左耳石数码图片上建立这11个地标点,并以第1个和第5个坐标点连线(长轴方向)作为基准线,获取1—11地标点的x、y坐标值(2D)。建立坐标点数据文件。

表1 四种金线鱼的样本信息Tab.1 Specification of samples of four Nemipterus species
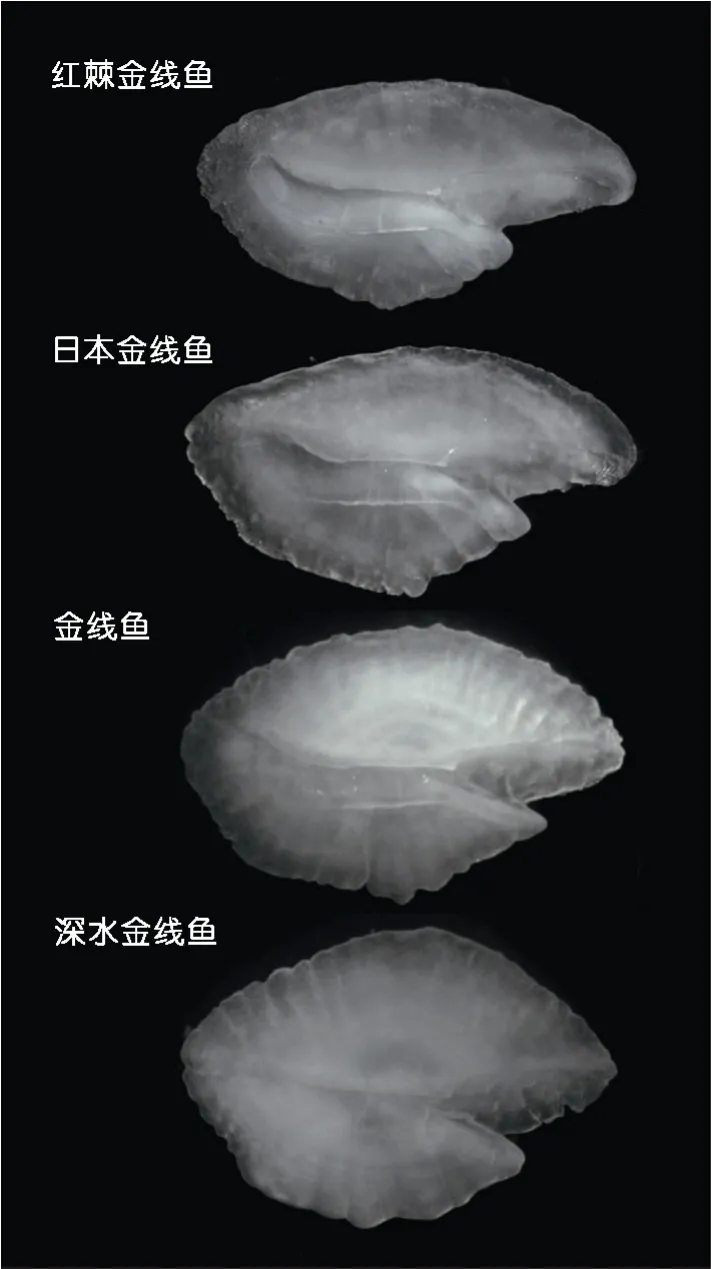
图1 四种金线鱼的左耳石形态Fig.1 Shapes of the left otolith of four Nemipterus species

表2 地标点类型与定义Tab.2 The landmark types and definition
1.2.2 平均形与薄板样条分析 利用 Tipsmall软件进行最小二乘准则(Least-squares criterion)回归分析,检验地标点有效性。用TpsRelw软件经普氏叠加出地标点,对每个样本的地标点进行平移置中、旋转和缩放,计算出质心距离,求出平均形(mean shape) ,据此进行局部扭曲(partial warps)和相对扭曲主成分分析(relative warps),保存软件生成的相对扭曲指数(relative warps scores)矩阵和分析报告。利用TpsRegr进行薄板样条分析,绘出四种金线鱼耳石的网格变形图,比较形态差异。

图2 以红棘金线鱼为例的11个地标点位置Fig.2 Positions of 11 landmarks in N.nemurus
1.2.3 判别分析 利用各样本的相对扭曲得分(relative warps scores)通过逐步判别分析法和交互验证法进行判别分析,判别方法采用 Bayes方法,用SPSS17.0进行统计分析。
2 结果
2.1 耳石形态描述
在4种金线鱼矢耳石样本的表征性变量中,长度性状上,以金线鱼长轴最长(10.50mm),深水金线鱼最小(5.23mm);宽度性状上以金线鱼短轴最长(6.93mm),红棘金线鱼最小(3.38mm)。耳石长宽关系上,深水金线鱼长宽比斜率最高,红棘金线鱼斜率最低,四种金线鱼在耳石长度性状相同下区别明显,重叠很少,表明耳石种的特征明显。形态系数上,红棘金线鱼和深水金线鱼在圆形近似率、矩形近似率、纵横比上变化不明显,形态稳定,种类特征较为明显,而金线鱼和日本金线鱼形态有所变化,重叠明显,不易区分;在成形系数、耳石听沟面积与总面积比两个量度性状上,四种金线鱼样本重叠明显,不能明显区分开(图3)。
2.2 几何形态分析
利用 TpsRelw软件根据地标点数据文件计算的四种金线鱼矢耳石平均形如图4所示,所有地标点重叠后的效果如图5所示。据此进行相对扭曲主成分分析,共提取了 18个主成分,其中第 1主成分贡献了30.78%,第2主成分贡献了28.44%,第3个主成分贡献了 11.75%,前三个主成分累积贡献了 70.97%。第1、2主成分的散点图见图6所示。11个地标点中在相对扭曲时贡献率较大的是Ⅰ型地标点6、10、11,累积贡献率59.63%;Ⅱ型地标点2,贡献率6.06%;Ⅲ型地标点7,贡献率29.67%(表3);表明Ⅰ型地标点和Ⅲ型地标点在四种金线鱼矢耳石形态区分中作用较大。对四种金线鱼矢耳石网格进行扭曲变形,相对于网格平均形,地标点 2、6、7、10、11变形较大,即表明四种金钱鱼的形态差异主要表现在听沟的头颈部(地标点6、7和11)和尾部上端(地标点10),以及基于尾部上端的耳石腹侧(ventral side)外部轮廓垂直点(地标点 2)(图6)。

图3 长度与宽度、成形系数、圆形近似率、矩形近似率、纵横比、听沟面积与总面积比率Fig.3 The scatter diagrams of length vs width,form factor,roundness,rectangular ratio,aspect ratio,sulcus and total area ratio

图4 矢耳石的平均形Fig.4 Mean shape of sagittal otolith

图5 重叠的地标点Fig.5 The superimposed landmarks

表3 不同地标点在相对扭曲分析时的贡献率Fig.3 Relative contribution of each landmark on relative warps

图6 相对扭曲第1、第2主成分散点图(主成分得分×102)Fig.6 Scatterplots of relative warp scores on the 1st and 2rd(Principal component scores are enlarged 102 times)
2.3 判别分析
将18个相对扭曲得分(Relative warp scores,RW)建立Bayes判别函数,通过逐步判别分析和交互验证法进行判别分析。利用 SPSS进行逐步法判别时,RW1、2、3、6、7、10、11、12、17 共计 9 个变量纳入到判别函数。但各组均值相等检验(one-way ANOVA)显示,RW6、7、10、12共4个变量均值相等(P>0.05),但未被剔除,如不剔除这 4个变量,将对模型的预测能力和分类效果产生一定影响。人工将其剔除后重新进行判别分析,共计5个变量纳入最终的判别分析,判别方程系数见表4。残余区别力分析表明,筛选后的 5个变量建立的预测模型效果较好(Wilks′ Lambda 检验,P<0.001,表5),可以作为判别分析的依据。结果表明,日本金线鱼 35个矢耳石样本中,错判入红棘金线鱼3条,判别成功率为91.4%;金线鱼32尾样本中,错判入深水金线鱼1条,判别成功率为96.9%;深水金线鱼36尾样本中,错判入金线鱼 4条,错判入红棘金线鱼 1条,判别成功率为86.1%;红棘金线鱼24尾样本中,错判入日本金线鱼2条,判别成功率 91.7%(表6,图7)。与逐步判别相比较,交互验证中深水金线鱼和红棘金线鱼判别结果与其相同,日本金线鱼和金线鱼的则略低。逐步判别分析和交互验证分析的判别正确率分别为91.3%和89.8%。因此,利用几何测量学方法进行四种金线鱼矢耳石判别分析是可行的,判别效果较好。

图7 4种金线鱼耳石典型判断分析散布图Fig.7 Scatterplots of canonical discriminant principle functions for four Nemipterus species

表4 4种金线鱼耳石性状的典型判别方程系数Tab.4 Fisher’s linear discriminant functions coefficients of sagitta shape variables for four Nemipterus species

表5 残余区别力分析Tab.5 The residual difference force analysis

表6 4种金线鱼耳石判别结果Fig.6 Otolith discrimination results of four Nemipterus species
3 讨论
3.1 耳石形态分析方法的选择与比较分析
本文采用几何形态测量学方法对北部湾四种金线鱼矢耳石进行了识别分析。结果表明,判别分析中有 89.8%—91.3%的判别成功率,识别效果较好,可以将这四种金线鱼矢耳石有效区分。在相同海域的三种金线鱼矢耳石研究中,基于傅里叶分析的轮廓学方法的判别成功率为 97.9%—98.6%(侯刚等,2012),比本研究中所采用的基于地标点的几何形态测量学方法的判别效果要高一些。对这三种金线鱼矢耳石判别成功率进行配对样本t检验,结果表明,在逐步判别分析和交互验证分析中,傅里叶方法与几何形态测量学方法判别效果均没有统计学意义上的显著性差异(t1=-3.475,P=0.074>0.05;t2=-3.311,P=0.080>0.05),即表明两种方法在矢耳石识别效果上接近。若对耳石形态差异进行描述,则几何形态测量学方法具有明显的优势。另外,由于本研究中的矢耳石种类特征明显,形态稳定,轮廓线上的叶形品状突(Leafed aragonite)变异不剧烈,因而提取的椭圆傅里叶标码能够较好的表达轮廓信息,77个谐值中有15个谐值经筛选纳入判别分析,贡献率作用较大,信息有效占比 19.5%(侯刚等,2012)。而几何形态测量学方法 18个相对扭曲得分中有5个纳入最终判别分析,信息有效占比 27.8%,明显高于傅里叶方法,表明地标法信息冗余要少一些。关于耳石形态方法比较方面的研究相对报道较少,Dominique(2006)比较了耳石形状系数、几何形态测量学和椭圆傅里叶方法在三种半棱属(Encrasicholina)鱼类和印度小公鱼(Stolephorus indicus)矢耳石种类区分的有效性,其结果表明,几何形态测量学法比另两种方法更有效一些,也能更加可视化地表达出耳石随着生长形态变异的趋向。李辉华等(2013)比较了传统形态测量法和傅里叶法在长江刀鲚、凤鲚种间以及刀鲚洄游生态型和淡水定居型的识别效果,认为两种方法在种间判别效果均较好,但是在种群区分上,傅里叶方法要更好些。而关于几何形态测量学和傅里叶法在鱼类耳石群体区分效果方面的比较研究,则报道较少(Cadrin,2000)。
在识别耳石形态方法选择时,可以采用耳石大小变量、耳石轮廓变化和地标点变量中的一种变量,也可以将其中两个变量或多种变量结合在一起来表征耳石形态特征,并且结合多种变量表征耳石形态时往往能提高种类和群体识别的成功率(窦硕增等,2012)。但值得注意的是,耳石大小变量、耳石轮廓变量和地标点变量是三种不同意义上的变量,其性质和单位不同,数据差异较大。无论是“降维”进行主成分分析,还是筛选“有效变量”进行判别分析,将多种变量结合在一起都有可能导致信息遗漏,从而影响识别效果。主成分分析是尽可能多地提取各变量的变异,且数据越相关,降维效果越好,而判别分析则是将参数层层筛选,剔除无效变量,提高模型预测能力和分类效果。因此,将多种变量参数结合在一起时,如果出现变量性质和数据差异太大、数据相关性降低、变量有效性互相影响等情况,则会降低数理分析的可信度或引起结果不良。因而,最好对同一性质的耳石变量进行数理分析,结合分析结果和耳石其他变量的特征来识别耳石形态,而不易将不同性质的耳石变量结合在一起进行数理分析。至于选取哪种耳石变量或方法,则应根据耳石材料特质或分析需求进行选择。
3.2 地标点的选择及其对耳石识别效果的影响
本研究中I型地标点6、10、11在相对扭曲主成分中贡献率最大,累计贡献率为59.53%,表明了I型地标点在 4种金线鱼矢耳石形态区分中有着重要作用;但是Ⅲ型地标点7贡献率达29.67%,表明该类地标点亦有重要作用。I型地标点6是基叶和翼叶的交点,Ⅲ型地标点 7为翼叶端点,二者贡献率合计为75.35%,表明4种金线鱼矢耳石的基叶和翼叶部分是重要的区分特征信息所在之处,这里地标信息不可忽略。倘若因避免主观性忽略了Ⅲ型地标点 7,则可能造成了信息丢失。因而,对地标点的选取,既要尽可能涵盖耳石形态所需的信息,选择合适的地标点数量,又要避免因无效地标点过多而造成信息冗余,降低数据彼此间相关性。
3.3 几何形态测量学方法在耳石形态分析中的应用展望
本研究中采用的几何形态测量学方法能够很好地区分 4种金线鱼属鱼类矢耳石,判别成功率为89.8%—91.3%,因而应用该方法对4种鱼类矢耳石进行种类识别具有很好的应用前景。由于耳石的形态变化可能受到遗传控制(Lombarteet al,1991;L’Abée-Lundet al,1993),因而基于几何形态测量学等方法所获得的耳石形态表型特征参数,亦可作为鱼类系统分类与进化研究的重要手段。另外,鱼类群体区分作为渔业资源评估与管理中的重要工作,几何测量学方法作为表型群体区分的现代形态学方法之一,较傅里叶方法在阐释形态变异和给出差异的生物学解释上更具有优势(Cadrin,2000),在鱼类群体区分中具有广阔的应用前景。
致谢感谢审稿人对本稿件提出的宝贵意见与建议。海渔1081班史亚峰、宋一平、李强、魏法俊等同学在野外采样与解剖实验中给予大力支持,在此谨表谢意!
王英俊,叶振江,刘 群等,2010.细条天竺鱼与黑鳃天竺鱼耳石形态识别的初步研究.海洋与湖沼,41(2):282—285
叶振江,孟晓梦,高天翔等,2007.中日两种花鲈(Lateolabraxsp.)耳石形态的地理变异.海洋与湖沼,38(4):356—360
白 明,杨星科,2007.几何形态测量法在生物形态学研究中的应用.昆虫知识,44(1):143—147
冯 波,程罗妹,2011.几何形态学方法区分短吻鲾地方种群的研究.上海海洋大学学报,20(5):677—681
闫宝荣,花保祯,2010.几何形态测量学及其在昆虫分类学和系统发育中的应用.昆虫分类学报,30(4):314—320
李辉华,郭弘艺,唐文乔等,2013.两种耳石分析法在鲚属种间和种群间识别效果的比较研究.淡水渔业,43(1):14—18
张国华,但胜国,苗志国等,1999.六种鲤科鱼类耳石形态以及在种类和群体识别中的应用.水生生物学报,23(6):683—688
国家技术监督局,2007.海洋调查规范-海洋生物调查.北京:中国农业出版社:57—59
侯 刚,冯 波,颜云榕等,2012.北部湾金线鱼、深水金线鱼与日本金线鱼矢耳石形态识别的初步研究.中国海洋大学学报,42(3):027—035
郭弘艺,唐文乔,魏 凯等,2007.中国鲚属鱼类的矢耳石形态特征.动物学杂志,42(1):39—47
萧旭峰,吴文哲,1998.生物形状的科学—浅谈几何形态测量学之发展与应用.科学月刊,29:624—633
窦硕增,于 鑫,曹 亮,2012.鱼类矢耳石形态分析及其在群体识别中的应用实例研究.海洋与湖沼,43(3):702—712
潘晓哲,高天翔,2010.基于耳石形态的属鱼类鉴别.动物分类学报,35(4):799—805
Bookstein F L,1990.Introduction to methods for landmark data.Proceedings of the Michigan Morphometrics Workshop.Spec Pub 2.University of Michigan Museum of Zoology,Ann Arbor,Michigan,USA:215—226
Cadrin S X,2000.Advances in morphometric identification of fishery stocks.Reviews in Fish Biology and Fisheries,10(1):91—112
Campana S E,Casselman J M,1993.Stock discrimination using otolith shape analysis.Canadian Journal of Fisheries and Aquatic Sciences,50(5):1062—1083
Dominique P,2006.Is geometric morphometrics efficient for comparing otolith shape of different fish species? Journal of Morphology,267(6):750—757
Klingenberg C P,Barluenga M,Meyer A,2003.Body shape variation in cichlid fishes of theAmphilophus citrinellusspecies complex.Biological Journal of the Linnean Society,80(3):397—408
L’Abée-Lund J H,Jensen A J,1993.Otoliths as natural tags in the systematics of salmonids.Environmental Biology of Fishes,36(4):389—393
Lombarte A,Rucabado J,Matallanas Jet al,1991.Taxonomía numé rica de Notothen Ⅱdae en base a la forma de los otolitos.Scientia Marina,55(2):413—418
Paul K,Oeberst R,Hammer C,2013.Evaluation of otolith shape analysis as a tool for discriminating adults of Baltic cod stocks.Journal of Applied Ichthyology,29(4):743—750
Rohlf F J,Marcus L F,1993.A revolution in morphometrics.Trends in Ecology &Evolution,8(4):129—132
猜你喜欢
大学教育(2022年3期)2022-05-16
小学生学习指导(低年级)(2021年12期)2021-12-31
辽宁省博物馆馆刊(2021年0期)2021-07-23
中华养生保健(2020年7期)2020-11-16
小学生学习指导(低年级)(2020年9期)2020-11-09
老友(2018年7期)2018-08-08
科学生活(2017年9期)2017-10-16
作文大王·低年级(2016年1期)2016-02-29
中国教育技术装备(2015年6期)2015-03-01
测绘学报(2015年8期)2015-01-14
