
几种常见保存方法对浮游动物生物量的影响*
2014-03-09 03:40冯秋园孙晓霞任琳琳
海洋与湖沼 2014年3期
冯秋园 孙晓霞 任琳琳
(1.中国科学院海洋研究所 山东胶州湾海洋生态系统国家野外科学观测研究站 青岛 266071;2.中国科学院大学 北京 100049)
浮游动物是海洋生态系统的重要组成部分,是海洋生物生产过程的核心之一,是调节海洋生态系统物质流和能量流的关键环节,是诸多国际研究计划的重要内容(孙晓霞,2011)。生物量的估算是研究浮游动物现存量和生产力的中心环节(Gaston,1996)。目前国际上对于浮游动物的研究常基于丰度这一概念,但是由于不同种类的浮游动物之间存在较大的差异,即使是同一类群的浮游动物,体型大小不同其生物量也存在差异,仅仅基于浮游动物的丰度难以实际地反映出浮游动物在生态系统中的地位和作用,因此,对浮游动物生物量的估算是海洋生态学研究的重要内容。5%的甲醛,-20°C冰冻以及液氮是几种常用的浮游动物保存方法。由于不同的保存方法会引起浮游动物生物量不同程度的变化,了解不同保存方法对浮游动物生物量的影响对于提高浮游动物生物量的估算精度非常关键。国际上针对福尔马林保存对浮游动物生物量的影响开展了一系列研究(Donaldet al,1977;Gastonet al,1996;Leuvenet al,1985;Dumontet al,1975;Kuhlmannet al,1982;Ajahet al,2003;Williamset al,1982;Giguereet al,1989),但是对液氮和冰冻保存对浮游动物样品所产生的影响研究较少,国内对这方面的研究则更为有限。由于生物量的损失程度与浮游动物的种类、保存方法和保存时间等具有很大的关系(Donaldet al,1977;Morris,1972),针对中国近海浮游动物优势种类,研究中国近海浮游动物生物量在不同保存条件下的变化规律对于估算中国近海浮游动物生物量非常必要。
中华哲水蚤(Calanus sinicus)和强壮箭虫(Sagitta crassa)是黄东海海域的浮游动物优势种(杨波等,1988;陈亚瞿等,1980),且在胶州湾全年都有分布(孙松等,2008)。在黄海海区,中华哲水蚤生物量较高,在5月生物量高达55.5mg/m3,占总生物量的29.6%(杨波等,1988)。春季,在东海赤潮高发区,中华哲水蚤作为饵料生物,优势度达 0.68,处于绝对优势地位(徐兆礼等,2003)。在胶州湾,强壮箭虫在数量上占有优势,年平均密度达45.3ind/m3(王倩等,2010)。湿重(WW)、干重(WD)和碳(C)、氮(N)含量是生物量的重要参数。本文以中华哲水蚤和强壮箭虫为研究对象,以湿重、干重和C、N含量为生物量参数,研究了上述几种保存方法对浮游动物生物量的影响,为直接测量生物量或通过体型-生物量方程间接估计生物量提供校正因子。在上述研究的基础上,利用体型-生物量间接估算法,结合图像自动识别技术,可以快速自动得到不同种类的浮游动物在较大时空范围内的生物量,为认识海洋食物网各营养级之间的物质循环和能量流动奠定基础。
1 材料和方法
1.1 样品采集
2012年3月在胶州湾进行采样,采样站位见图1,A1-D7表示在胶州湾进行常规监测的采样点位置,由于C3和D7站浮游动物的数量相对较大,因此本实验所用的浮游动物样品主要采于C3和D7两站。使用浮游生物浅水I型网(网口直径50cm,网长145cm,网孔 500μm)由底到表垂直拖网采集,采到的鲜活样品置于冰桶中带回实验室处理。

图1 胶州湾采样站点Fig.1 Sampling stations in the Jiaozhou Bay,the Yellow Sea
1.2 实验方法
本实验采用5%甲醛过滤海水溶液,-20°C冰冻和液氮保存3种保存方法。甲醛保存法,用GF/F玻璃纤维滤膜过滤的海水与分析纯的甲醛配制成体积比为95︰5的甲醛过滤海水溶液。将已经测量完鲜重的浮游动物样品用镊子轻轻地转移到容积为10ml的小试剂瓶中,加入 5ml浓度为 5%的甲醛过滤海水溶液在阴凉处保存。-20°C冰冻保存法,实验中将已测量完鲜重的样品平铺于筛绢上,将筛绢放于5ml的冻存管中于-20°C条件下进行保存。液氮保存法与-20°C冰冻保存法的处理方法相同。
每种保存方法的最长保存时间为60 d,分别设置0、1、3、5、7、10、15、20、25、30、35、40、45、50、60 d共15个测量时间点,每个时间点设置a,b两个平行样。采集回来的样品立即进行分类,挑取完整、健壮、活的个体,每个样本大约20—30个个体(保证样品干重至少达到 2mg)。用少量蒸馏水快速轻轻冲洗,去除生物体表的杂质和盐分,再用吸水纸吸取生物体表面的水分,用 METTLER TOLEDO XS105精密天平测量鲜重(WF)。称量后的样品分别用甲醛,冰冻和液氮保存,在到达设定的时间点以后取出对应的样品,其中甲醛保存的样品采用上面同样的方法清洗,称量湿重(WW)。称量后的个体置于事先处理好的称量瓶中,放于烘箱中于60°C下干燥24h,然后于干燥器中冷却,待样品冷却到室温以后测量干重(WD),再用德国Vario EL Ш元素分析仪测定C、N含量W(C)、W(N)。得到WF、WW、WD和W(C)、W(N)分别与样品初始的WF作比,用WW/WF,WD/WF,W(C)/WF和W(N)/WF表示,与保存时间作图分析其变化。
2 结果
2.1 中华哲水蚤在几种保存条件下的生物量变化
中华哲水蚤在甲醛、冰冻和液氮3种保存条件下的生物量变化如图2所示。中华哲水蚤在上述3种保存条件下,WW呈现出不同的变化趋势。在甲醛保存条件下,其WW在保存的前几天急剧地上升,达到了WF的127.6%,之后不断下降,在2个月后下降为WF的81%;在冰冻保存条件下,其WW呈一直下降的趋势,2个月后下降为WF的66.6%;在液氮保存条件下,中华哲水蚤的WW呈现先下降后上升的趋势,在保存 2个月后为WF的91.9%。
中华哲水蚤在甲醛保存条件下,其WD在保存初期呈现出 1个小的上升,之后下降,后趋于稳定,在保存2个月后变为WF的14.2%;在冰冻保存条件下,WD变化趋势与在甲醛保存条件下的变化趋势相同,但是下降幅度比在甲醛保存条件下要小,在保存2个月后,WD变为WF的22.34%;在液氮保存条件下,WD呈现上升的趋势,保存2个月后为WF的26.7%。
对于中华哲水蚤,其W(C)在 3种保存条件下的变化趋势与W(N)相同。在5%的甲醛保存条件下,中华哲水蚤的W(C)、W(N)含量先下降后趋于稳定,保存 2个月后W(C)/WF和W(N)/WF分别为 7.03%和1.20%;冰冻保存条件下,W(C)、W(N)先短暂上升后下降,再趋于稳定,保存 2个月后W(C)/WF和W(N)/WF下降为10.3%和2.15%;在液氮保存条件下,W(C)、W(N)呈现出稳定上升的趋势,在保存 2个月后W(C)/WF和W(N)/WF分别为11.3%和2.41%。
综合分析发现,中华哲水蚤的WW在3种方法保存2个月后都有不同程度的下降,其下降程度分别为冰冻>甲醛>液氮。与新鲜样品相比,甲醛保存2个月后的样品,其WD和W(C)、W(N)都降低了,分别下降了 29.4%、23.01%和43.3%。而冰冻和液氮保存2个月后的样品,其WD和W(C)、W(N)有所上升。WD、W(C)、W(N)在冰冻保存条件下,分别上升了11.14%、12.88%、1.94%;在液氮保存条件下,分别上升了32.59%、23.26%、14.65%。

图2 中华哲水蚤各生物量指标在5%甲醛、-20°C冰冻和液氮保存条件下的变化Fig.2 Changes in biomass of Calanus sinicus preserved in 5% formalin,-20°C in liquid nitrogen
利用SPSS统计软件中的Duncan法,对中华哲水蚤在5%甲醛,-20°C和液氮3种保存条件下的的生物量变化进行方差分析和多重比较,不同处理结果的差异显著性用标记字母法表示,如表1所示,相同的字母表示不同的处理之间的差异性不显著,不同的字母表示不同的处理之间具有极显著的差异,如果某一处理的结果是两个字母的组合,例如AB,而另2组处理的结果分别是组成上述字母组合的某一个字母,即分别为A和B,则表明前一处理分别与后两种处理之间均没有显著性差异。经Duncan检验,上述3种保存方法对中华哲水蚤的WW和WD的影响,存在极显著性差异(P<0.01),对中华哲水蚤W(C)和W(N)的影响无显著性差异(P>0.05)。5%的甲醛保存对中华哲水蚤WW和WD的影响与-20°C冰冻和液氮保存之间存在显著性差异,而后两种保存方法之间无显著性差异。5%甲醛保存导致中华哲水蚤WW的下降程度比-20°C冰冻和液氮保存小,5%甲醛导致中华哲水蚤WD的下降程度比-20°C冰冻和液氮保存大。
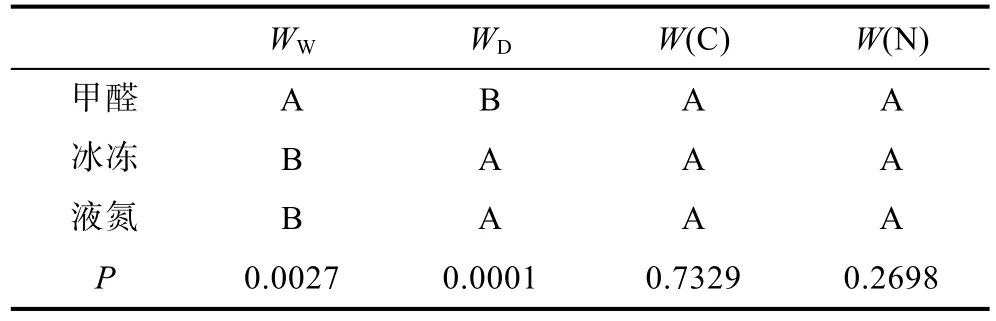
表1 中华哲水蚤在5%甲醛、-20°C、液氮3种保存条件下生物量变化的多重比较Tab.1 Changes in biomass of Calanus sinicus preserved in 5%formalin,-20°C in liquid nitrogen
2.2 强壮箭虫在几种保存条件下的生物量变化
在甲醛保存条件下,强壮箭虫的WW、WD、W(C)和W(N)呈现出先上升再下降后上升的趋势(图3),在保存到大约第3—5d的时候,各生物量指标有一个小的上升,之后下降,在第7—10d的时候下降到最低点,然后再上升,在保存2个月后WW较WF上升了4%,与新鲜样品相比,WD、W(C)和W(N)分别下降了44.3%、44.4%和51.5%。在冰冻保存条件下,WW、WD、W(C)和W(N)一直呈现下降的趋势,WW起初变化比较缓慢,在第10d左右开始急剧下降,WD、W(C)和W(N)从保存的第1d左右开始急剧下降,之后缓慢下降,在保存 2个月后,与新鲜样品的各生物量指标相比,WW、WD、W(C)和W(N)分别下降了的29.3%、23.5%、23.1%和29.1%;在液氮保存条件下,WW、WD、W(C)和W(N)最初都呈现急剧下降的趋势,WW和WD在保存到大约40d的时候下降到最低点,W(C)和W(N)在大约第7d的时候下降到最低点,之后各生物量指标又略有上升的趋势,在保存 2个月后,与新鲜样品的各生物量指标相比,WW、WD、W(C)和W(N)分别下降了26.85%、 30.74%、 16.74%和20.97%。综合分析发现,强壮箭虫的WW在甲醛保存2个月后上升为WF的104%,而在冰冻和液氮保存2个月后分别下降为WF的70.7%和73.2%。WD在3种保存条件下保存2个月后的下降幅度大小为:5%的甲醛>液氮保存>-20°C冰冻。W(C)和W(N)的下降幅度大小为:5%的甲醛>-20°C冰冻>液氮。
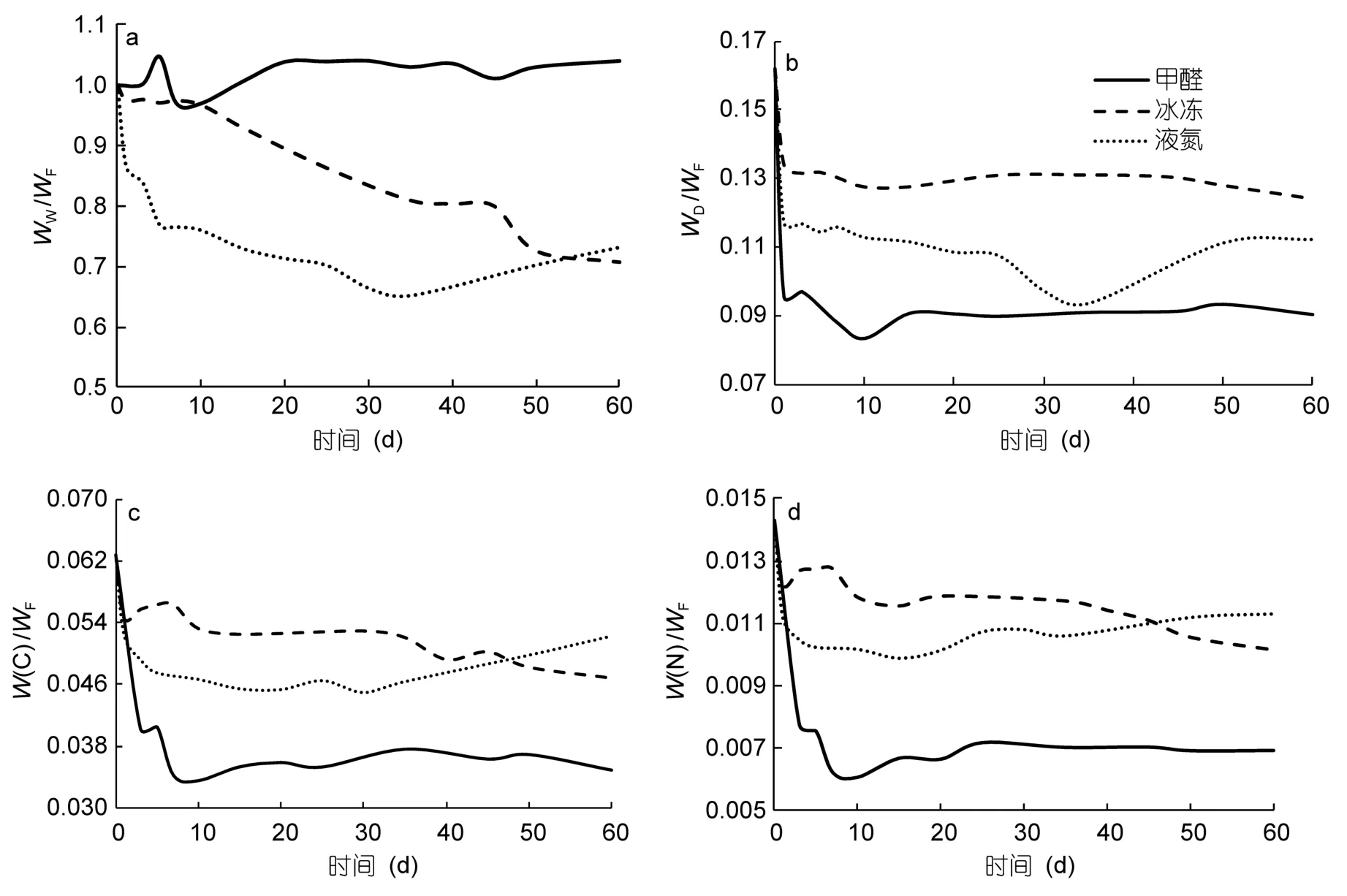
图3 强壮箭虫生物量在5%甲醛、-20°C冰冻和液氮保存条件下的变化Fig.3 Changes in biomass of Sagitta crassa preserved in 5% formalin,-20°C in liquid nitrogen
利用 Duncan法,对强壮箭虫在 5%甲醛,-20°C和液氮 3种保存条件下的生物量变化进行方差分析和多重比较,结果如表2所示。经Duncan检验,上述3种保存方法对强壮箭虫生物量的影响存在显著性差异(P<0.05),其中对WD的影响存在极显著性差异(P<0.01)。经过多重比较,-20°C冰冻保存导致强壮箭虫WW的损失程度比液氮小,与5%甲醛相比,2种保存方法之间无显著性差异,5%甲醛保存和液氮保存对强壮箭虫WW的影响不存在显著性差异。5%甲醛保存导致强壮箭虫WD的损失程度比-20°C冰冻保存和液氮保存大,-20°C冰冻保存和液氮保存对强壮箭虫WD的影响不存在显著性差异。5%甲醛导致强壮箭虫W(C)的损失程度比-20°C 冰冻大,与液氮保存相比,2种方法之间无显著性差异。-20°C冰冻保存和液氮保存对强壮箭虫W(C)的影响不存在显著性差异。5%甲醛保存导致强壮箭虫W(N)的损失程度比液氮保存大,与-20°C冰冻相比,2种方法之间无显著性差异,-20°C冰冻保存和液氮保存对强壮箭虫W(N)的影响差异性不大。
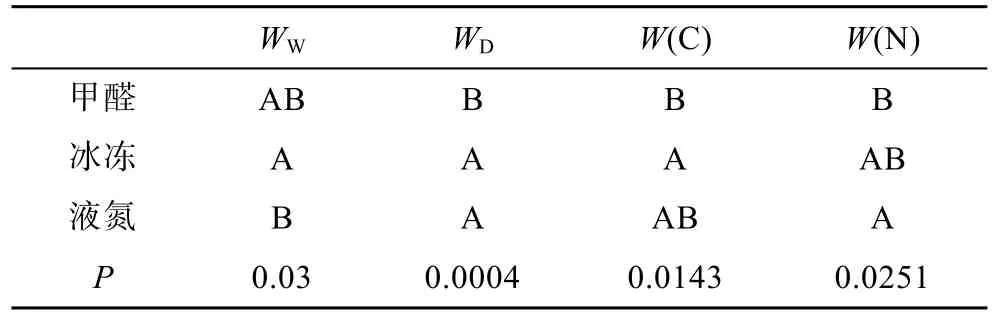
表2 强壮箭虫在5%甲醛、-20°C、液氮三种保存条件下生物量变化的多重比较Tab.2 Changes in biomass of Sagitta crassa preserved in 5%formalin,-20°C in liquid nitrogen
3 讨论
本文采用胶州湾的优势浮游动物中华哲水蚤和强壮箭虫为研究对象,以WW、WD、W(C)和W(N)为生物量指标,研究了5%甲醛,-20°C冰冻和液氮3种保存条件对其生物量的影响。对于甲壳类中华哲水蚤而言,保存2个月后,3种保存方式都会导致其下降,但是甲醛保存的中华哲水蚤在保存之初会有一个上升,之后迅速下降,Mills(1982)在“保存方法对海洋底栖无脊椎动物的影响”一文中也指出大多数生物在甲醛保存时最初生物量会上升,之后下降。研究发现 3种保存条件下WW的下降幅度分别是冰冻>甲醛>液氮。中华哲水蚤在甲醛保存条件下,在保存之初WW的上升可能是由中华哲水蚤体表的几丁质保护层等结构在甲醛的作用下通透性增加,甲醛保存液进入生物体内所导致的。之后WW下降可能是由中华哲水蚤的细胞膜等结构被破坏,体液流失量增加所导致的。而在冰冻和液氮保存条件下,WW一直下降的原因,推测是在保存过程中细胞膜受到损伤,体液流失造成的。导致中华哲水蚤WW出现以上变化趋势的具体原因,还需要进一步的生理生化实验验证。与新鲜样品相比,甲醛保存 2个月后,WD、W(C)和W(N)下降了,冰冻和液氮保存2个月后,其WD、W(C)和W(N)有所上升,增加幅度为液氮>冰冻。从实验结果分析发现不同的保存方法会导致浮游生物的生物量有不同的变化趋势。Mills(1982)研究发现在10%的甲醛保存液中,只有双壳类生物会出现明显的生物量损失,而其他不同的生物则会出现生物量不同程度的上升或下降。Kalevi等(1979)研究了一种剑水蚤(Megacyclops gigas)在 4%甲醛、酸性鲁格试剂、带水在-20°C冰冻和用稀释的甲醛溶液在-20°C保存后生物量的变化,研究表明,-20°C条件下用稀释的甲醛溶液保存剑水蚤对生物量的影响最小,几乎没有损失,接近新鲜样品的生物量,而4%甲醛和酸性鲁格试剂,以及-20°C条件下用水保存的样品则生物量损失较大。Fudge(1968)研究了新糠虾类的Neomysis integer在保存后的生化成分变化,研究发现三氯乙酸(TCA)保存下的干重较新鲜样品的WD要大,而蛋白质、脂质、碳水化合物等成分则有所下降。而冰冻保存后的Neomysis integer各种化学成分都有所上升,WD较新鲜样品上升了13%,与本实验的实验结果较为相似。分析甲醛保存的样品WD和W(C)先上升后下降,而W(N)一直下降的原因,可能是由于在保存初期,几丁质保护层在甲醛的作用下通透性增加,甲醛保存溶液进入生物体内,同时体液流失导致小部分蛋白质或是氨基酸也通过细胞膜和体壁渗透到体外,保存初期甲醛保存液的渗透作用大于体液的流失作用,由于甲醛含有丰富的C元素,因此使得WD和W(C)出现一个上升趋势,而W(N)则一直呈现下降趋势,而保存后期,由于细胞结构的完全被破坏,体液的流失作用增强,因此WD和W(C)开始呈现下降趋势。分析在冰冻和液氮保存下,WD、W(C)和W(N)上升的原因,结合文献资料(Fudge,1968)发现,冰冻保存条件下,浮游动物的蛋白质、碳水化合物和脂质等成分都有所增加,导致WD的增加,具体原因尚不明确。而随着保存时间的增加,冰冻条件下WD、W(C)和W(N)有所下降,推测可能是由某些耐冷微生物的繁殖消耗了部分的有机体导致的。而液氮条件下,极低的温度有效地抑制了微生物的繁殖,因此液氮保存下的浮游动物WD、W(C)和W(N)没有出现下降趋势。以上是作者依据实验结果,结合知识背景和查阅的资料,做出的推测和设想,导致中华哲水蚤WD、W(C)和W(N)出现以上变化的具体原因如何,还需要进一步的生理生化分析进行验证。单纯从生物量损失的角度考虑,若是测量WW,推荐使用液氮保存的方法,相较于另外2种方法,这种方法导致WW的损失最小。若是测量WD、W(C)和W(N),则推荐使用冰冻保存方法,因为冰冻保存后的样品其生物量最接近真实的生物量,而且在最初的下降之后,其变化逐渐趋于稳定,波动幅度较小。若通过体型-生物量方程来间接估算生物量时,还需要考虑各种保存方法对浮游动物体型的影响,以及操作可行性。
对于毛颚类的强壮箭虫而言,甲醛保存2个月以后,其WW大于WF。冰冻和液氮保存2个月后,其WW小于WF,其下降幅度为液氮>冰冻。分析其原因,可能是由于强壮箭虫其生理结构不存在几丁质层的保护,而且在甲醛保存条件下强壮箭虫的细胞膜被破坏,渗透调节功能消失,箭虫体液的渗透压大于甲醛溶液,导致大量水分进入箭虫体内,以致出现WW大于WF的现象。而在冰冻和液氮保存条件下,强壮箭虫的细胞膜被破坏,体液大量流失,因此使得WW下降。强壮箭虫的WD、W(C)和W(N)在3种保存方式下保存2个月后,都分别小于新鲜样品的各生物量指标,下降幅度分别为甲醛>液氮>冰冻,但是在液氮保存的后期,强壮箭虫的WD、W(C)和W(N)略有上升。分析其原因,由于在3种保存条件下强壮箭虫的细胞膜被破坏,体液大量流失导致WD、W(C)和W(N)出现了明显的下降,而在液氮保存后期,WD、W(C)和W(N)的增加可能是由生物体和液氮及周围环境之间发生了某些反应造成的,对导致此现象产生的原因还需要进一步的生理生化实验来确定。关于保存方法对毛颚类动物生物量影响的相关报道较少,Makoto Omori(1978)用以硼砂为缓冲剂的甲醛溶液和以四氮六甲圜为缓冲剂的甲醛溶液对拿卡箭虫(Sagitta nagae)进行保存,研究其对拿卡箭虫生物量的影响,研究发现在以硼砂为缓冲剂的甲醛溶液中,保存前2.5天箭虫的有机质和化学成分含量下降剧烈,之后变缓,保存1周后箭虫有机质的损失达到21%,C损失34%,N损失49%,W(C):W(N)由8.5变为11.0,保存1个月后生物量达到稳定,损失速率几乎变为0。以四氮六甲圜为缓冲剂的甲醛溶液中,在保存的前0.5 d,生物量和各化学成分迅速下降,之后的2d,生物量又有所上升,之后生物量缓慢下降,保存1周后,箭虫损失了13%的C和26%的N,但是有机质的损失非常的小。浮游动物在以四氮六甲圜为缓冲剂的甲醛溶液中比在以硼砂为缓冲剂的甲醛溶液中的生物量损失要小。单从生物量方面考虑,若是测量强壮箭虫的WW,推荐使用甲醛保存方法,因为相较于另外2种保存方法,甲醛保存下的强壮箭虫,其WW最接近其真实值,而且变化也最稳定,若是测量WD、W(C)和W(N),推荐使用冰冻保存方法,因为其WD、W(C)和W(N)损失最小,其次是液氮保存。但是同样,若采用体型-生物量方程间接估算其生物量,则需要综合考虑其各种保存条件对体型的影响。在实验过程中发现,甲醛保存对强壮箭虫体型的影响最小,也最稳定,而且操作起来也最简单,适合大样本的研究。因此在选取一定的保存方法对强壮箭虫的生物量进行研究时,要根据实验目的和实验可操作性等因素进行综合考虑和选择。
查阅国内外的相关资料并结合本实验综合分析可以得出,不同的保存方法会导致浮游动物生物量出现增加和降低不同的变化趋势,而且增加和降低的幅度也有所差异,这与采用的保存剂,保存条件、保存溶液的量,以及不同的保存方法对不同的浮游动物作用机理不同等原因有关,而且在操作过程中不同的操作方法,也会导致实验结果存在一定的差异。实验发现浮游动物的生物量在保存的初期变化比较明显,之后逐渐趋于稳定,这与 Beers等(1976)的实验结果相似。各种保存方法对中华哲水蚤生物量的影响要小于强壮箭虫,这可能与甲壳类生物表面的几丁质保护有关,有效地抑制了体液的流失。经各种保存方法处理后,浮游动物 N的损失量要大于 C,这与许多已发表的研究结果类似(Williamset al,1982,Champalbertet al,1979)。据推测,保存样品中N的流失主要是由于一些自由氨基酸和小分子的含 N分子造成的,这些富含 N的物质用来维持细胞内的渗透压(Beers,1976)。
本文选取中国近海两种重要的浮游生物中华哲水蚤和强壮箭虫为研究对象,通过研究了解 5%甲醛、-20°C冰冻和液氮3种不同的保存条件对中华哲水蚤和强壮箭虫生物量的影响,为更为准确地估算其生物量的真实值提供了一定的矫正因子,也为在不同条件下选取合适的保存方法提供了依据。中国近海浮游动物种类丰富,不同的浮游动物种类之间存在生理生化结构上的差异,因此不同种类的浮游动物在相同的保存条件下,生物量变化存在很大的差异。除本文所研究的这3种常见的浮游动物保存方法外,不同浓度下的甲醛和乙醇,以及添加不同缓冲剂的甲醛溶液也是常见的浮游动物保存方法。在以后的研究中,会进一步增加不同的保存条件对中国近海海域更多浮游动物优势种类生物量的影响的研究,为全面、准确地估算中国近海优势浮游动物的生物量提供理论支持。
王 倩,孙 松,霍元子等,2010.胶州湾毛颚类生态学研究.海洋与湖沼,41(4):639—644
左 涛,王 克,李超伦,2003.南黄海中华哲水蚤体长-干重的关系.水产学报,27(增刊):103—105
孙 松,周 克,杨 波等,2008.胶州湾浮游动物生态学研究I.种类组成.海洋与湖沼.39(1):1—7
孙晓霞,孙 松,王世伟等,2011.图像自动识别技术在胶州湾浮游动物生态学研究中的应用.海洋与湖沼,42(5):647—653
杨 波,徐汉光,1988.黄海主要桡足类的生物量.大连水产学院学报,3—4:35—42
陈亚瞿,朱启琴,陈清潮,1980.东海浮游动物量的分布特征.海洋学报.2(4):115—121
徐兆礼,洪 波,朱明远等,2003.东海赤潮高发区春季浮游动物生态特征的研究.应用生态学报,14(7):1081—1085
Ajah P O,Nunoo F K E,2003.The Effect of Four Preservation Methods on Length,Weight and Condition Factor of the ClupeidSardinella auviritaVal.1847.J.Appl.Ichthyol.19(2003):391—393
Beers J R,1976.Determation of zooplankton biomass.Paris:Unesco Press,35—84
Champalbert G,Kerambrun P,1979.Influence du mode de conservation sur la composition chimique élémentaire de Pontella mediterranea(Copepoda:pontellidae).Marine Biology,51(4):357—360
Donald G L,Paterson C G,1977.Effect of Preservation on Wet Weight Biomass of Chironomid Larvae.Hydrobiologia,53(1):75—80
Dumont H J,Van de Velde I,Dumont S,1975.The Dry Weight Estimate of Biomass in a Selection of Cladocera,Copepoda and Rotifera from the Plankton,Periphyton and Benthos of Continental Waters.Oecologia(Berl),19:75—97
Fudge H,1968.Biochemical Analysis of Preserved Zooplankton.Nature,219:380—381
Gaston G R,Bartlett J H W,Mcallister A P,1996.Biomass Variations of Estuarine Macrobenthos Preserved in Ethanol and Formalin.Estuaries,19(3):674—678
Gigugre L A,St-Pierre J F,Bernier Bet al,1989.Can We Estimate the True Weight of Zooplankton Samples after Chemical Preservation? Can J Fish Aquat Sci,46:522—527
Kuhlmann D,Fukuhara O,Rosenthal H,1982.Shrinkage and Weight Loss of Marine Fish Food Organisms Preserved in Formalin.Bull Nansei Reg Fish Res Lab,14:13—18
Leuven R S E W,Brock T C M,van Druten H A M,1985.Effects of Preservation on Dry-and Ash-Free Dry Weight Biomass of Some Common Aquatic Macro-invertebrates.Hydrobiologia,127:151—159
Mills E L,Pittman K,Munroe B,1982.Effect of Preservation on the Weight of Marine Benthic Invertebrates.Can J Fish Aquat Sci,39:221—224
Morris R J,1972.The preservation of Some Oceanic Animals for lipid Analysis.Journal of the Fisheries Research Board of Canada,29(9):1303—1307
Omori M,1978.Some Factors Affecting on Dry Weight,Organic Weight and Concentrations of Carbon and Nitrogen in Freshly Prepared and in Preserved Zooplankton.Hydrobiol,63(2):261—269
Salonen K,Sarvala J,1980.The Effect of Different Preservation Methods on the Carbon Content of Megacyclops Gigas.Hydrobiologia,72:281—285
Wiliams R,Robins D B,1982.Effects of Preservation on Wet Weight,Dry Weight,Nitrogen and Carbon Contents of Calanus helgolandicus(Crustacea:Copepoda).Marine Biology,71:271—281
猜你喜欢
皮肤病与性病(2021年3期)2021-07-30
生物学通报(2021年9期)2021-07-01
辽宁化工(2021年5期)2021-06-03
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30
黑龙江动物繁殖(2020年2期)2020-12-17
青年歌声(2019年7期)2019-07-26
趣味(语文)(2019年3期)2019-06-12
童话王国·文学大师班(2017年3期)2017-07-06
新高考·英语基础(高一)(2016年11期)2017-07-06
中国医疗美容(2015年2期)2015-07-19
