
大白菜雄性不育系MS200712 的研究简报
2014-02-23 10:50
中国蔬菜 2014年8期
(北京市农林科学院蔬菜研究中心,北京 100097)
大白菜雄性不育源主要有细胞核雄性不育和细胞质雄性不育。细胞核雄性不育是张书芳等(1990)在大白菜品种万泉青帮采种田中发现的;细胞质雄性不育则由原生质体融合、杂交和多代回交等途径,转育萝卜Ogu CMS、甘蓝型油菜Pol CMS 和新型甘蓝型油菜CMS96 而获得的大白菜Ogu CMS(Williams,1981;Health et al.,1994)、Pol CMS(柯桂兰 等,1992)和CMS96(张德双 等,2002)细胞质雄性不育系。可见,大白菜细胞质雄性不育系的主要来源是转育已有的十字花科细胞质雄性不育系,目前还未见直接利用芸薹属种间杂交的方法创制新型大白菜雄性不育系的报道。
为了拓宽大白菜雄性不育系资源、类型以及研究雄性不育发生的机制,本试验以芸薹属种间杂交的方法创制和获得新型大白菜雄性不育系,为今后在大白菜育种中更广范地应用雄性不育系奠定 基础。
1 材料与方法
1.1 材料
供试芸薹属异源六倍体(AABBCC,n=27)拥有芸薹A、黑芥B、甘蓝C 的全部染色体,主要性状表现为:株型高大,心叶略带紫色,叶片绿色、有缺刻、覆蜡粉,茎部粗大、略带紫色、覆蜡粉,腋芽萌发能力强,花瓣黄色,花药和柱头正常,结实性好,种荚细长,种子较大、黑褐色(张德双 等,2009),该异源六倍体来源于北京市农林科学院蔬菜研究中心大白菜课题组;北京新3号的父本832172 也来源于本中心大白菜课题;大白菜品种金春购自辽宁东亚国际种苗有限公司,义和5号购自青岛胶州市胶白蔬菜育种中心。
1.2 方法
1.2.1 杂交方法 植株盛花期时,在父本花序上预先套硫酸纸袋,选取母本生长旺盛、花蕾多的主枝做杂交,彻底摘除用于杂交的全部花蕾的6个雄蕊,然后涂抹父本的花粉到母本柱头上,套上硫酸纸袋,并用曲别针扎口,以防止被昆虫携带的花粉污染;第2 天上午再涂抹父本的花粉到母本柱头上,然后扎好袋口。杂交完成后应注意浇水,及时修剪多余枝条,撤掉硫酸纸袋。在母本上收获的种子即为杂交一代。
1.2.2 试验步骤 试验在本中心日光温室内完成。首先,以芸薹属异源六倍体为母本,以大白菜品种金春为父本进行杂交,获得F1(AABBCC×AA);以大白菜品种义和5号为母本,以F1(AABBCC×AA)为父本再次进行杂交。在多亲本杂交后代中即可筛选到大白菜雄性不育系MS200712(图1)。
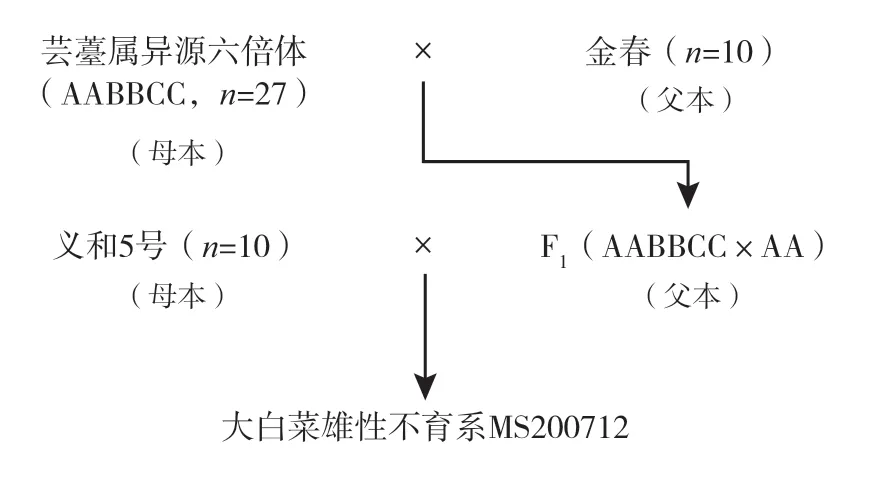
图1 获得大白菜雄性不育系MS200712 的技术路线
最后,用义和5号、北京新3号的父本832172 与大白菜雄性不育系MS200712 进行回交转育,并鉴定转育后代的花粉育性。
2 结果与分析
2.1 杂交一代F1(AABBCC×AA)的获得
在金春的花序上预先套上硫酸纸袋,彻底摘除芸薹属异源六倍体花序中用于杂交的全部花蕾的6个雄蕊,然后涂抹金春的花粉,套上硫酸纸袋,并扎好袋口;第2 天上午再重复授粉1次。在芸薹属异源六倍体上收获的种子即为F1(AABBCC× AA)。
在人工授粉获得F1(AABBCC×AA)的过程中,为了防止因雄蕊去除不彻底而混杂自交种子,在剥蕾时只注重质量,不追求数量,AABBCC 异源六倍体的花药被彻底去除。采用2种保障措施:①用镊子剥蕾时,舍弃掉那些不容易操作的小花蕾,每一个花序的授粉花蕾数不要过多,以5~10个为宜;②在第2 天重复授粉时,再次确认是否残留母本的花药。由图2可见,F1叶片具有叶翼、长倒卵形,抽薹、现蕾期较晚;而AABBCC 异源六倍体叶片没有叶翼、近圆形,抽薹、现蕾期较早(张德双 等,2009)。可见,F1与母本的性状已经存在较明显差异,特别是F1叶片具有叶翼,标志着F1基因组中存在父本金春AA基因组。SRAP 标记的电泳检测结果进一步证明F1为真正的杂交种(待 发表)。

图2 F1(AABBCC×AA)的植株
2.2 大白菜雄性不育系MS200712 的获得和鉴定
在F1(AABBCC×AA)花序上预先套上硫酸纸袋,彻底摘除义和5号花序中用于杂交的全部花蕾的6个雄蕊,然后涂抹F1(AABBCC×AA)的花粉,套上硫酸纸袋,并扎好袋口;第2 天上午再重复授粉1次。在义和5号上收获的种子即包含大白菜雄性不育系MS200712。
翌年春季继续播种MS200712。在盛花期观察MS200712 的花药,结果花药完全退化、败育彻底,表现为雄性不育,即获得了人工创造的大白菜雄性不育系MS200712。该雄性不育系叶片深绿色、长椭圆形,叶缘多锯齿,花瓣黄色,花药浅黄色、退化、败育彻底,花药长短正常,柱头正常,蜜腺发达,可正常吸引昆虫授粉(图3~5)。

图3 大白菜雄性不育系MS200712 的植株和花序

图4 大白菜雄性不育系MS200712 的花器官

图5 大白菜雄性不育系MS200712 的花药和蜜腺
2.3 大白菜雄性不育系MS20071208-840 和MS20071208-916 的获得
以MS200712 雄性不育系为母本,分别与义和5号、832172 进行杂交,在MS200712 不育系上 收获的种子即为MS20071208-840和MS20071208-916。分别播种MS20071208-840和MS20071208-916,在盛花期观察花药的育性,结果大白菜雄性不育系MS200712 不仅可以与大白菜品种正常杂交、结实,而且MS20071208-840和MS20071208-916的花药依然表现为稳定的雄性不育性。但MS200712所属的不育类型有待进一步确认。
3 结论与讨论
目前,在大白菜育种中3种主要的细胞质雄性不育系Ogu CMS、Pol CMS 和CMS96 应用较多。但改良的Ogu CMS 不育系存在回交转育多代后植株生长势严重退化、杂交优势差等缺点;Pol CMS 不育系由于温度敏感存在育性受环境条件影响容易出现有粉株等缺点(张德双,2006);CMS96 不育系虽然克服了上述缺点,但在实际应用中存在个别组合制种产量偏低的问题。因此,在大白菜育种中新型不育系的不断推陈出新显得尤为重要,不仅有利于提高杂交种的纯度,保护育种者的利益,而且提供了深入开展雄性不育系机理探究的平台。2004年10月笔者引入芸薹属异源六倍体材料,初衷是以芸薹属异源六倍体为桥梁,将紫色甘蓝的紫色性状转到大白菜品种中,以期获得类似于紫色甘蓝叶片的紫色大白菜资源。首先,采用芸薹属异源六倍体与紫色甘蓝进行正反交,试图导入甘蓝的紫色基因,实践中发现,虽然每年的授粉工作量很大,但很难获得真正的杂交种;然后,用芸薹属异源六倍体与大白菜杂交,结果发现二者可以正常杂交;于是,选用芸薹属异源六倍体与大白菜杂交,以期拓宽大白菜育种资源特别是雄性不育资源。
3.1 种间杂交拓宽了大白菜雄性不育系类型
在以往的研究中,有很多经由属间或种间杂交途径获得雄性不育尤其是新型细胞质雄性不育的成功范本可供借鉴。Shiga 和Baba(1971,1973)在甘蓝型油菜千荚油菜(Chisayanatane)×北陆23(Hokuriku 23)的F1中发现S-CMS 细胞质雄性不育(属于Nap CMS)。Thompson(1972)在冬春甘蓝型油菜RD58×Bronowski 的F2中发现T-CMS细胞质雄性不育(属于Nap CMS),反交则花粉正常。李殿荣(1984)在甘蓝型油菜S74-3(陕西)×〔丰收4号(四川)×7207(西藏)〕多亲本杂交后代中发现雄性不育株,并育成1763A和6223A 雄性不育系。本试验中先配制远缘杂种(AABBCC×AA),再用普通大白菜品种义和5号作母本、(AABBCC×AA)作父本进行杂交,结果在义和5号×(芸薹属异源六倍体×金春)多亲本杂交后代中首次发现雄性不育材料MS200712,这是本试验的最大收获。之所以选用春种大白菜金春和秋种大白菜义和5号作亲本,是因为在进行杂交授粉时金春、义和5号与芸薹属异源六倍体的花期正好相遇。
3.2 大白菜雄性不育系MS200712 的败育原因
以雄性不育系MS200712 为母本,分别与义和5号、北京新3号的父本832172 进行杂交,结果表明大白菜雄性不育系MS200712可以与大白菜品种正常杂交,并获得回交二代MS20071208-840和回交一代MS20071208-916,但结实较少。经花期鉴定,播种的3株MS20071208-840和5株MS20071208-916全部表现为花药败育。
雄性不育系MS200712 为多亲本杂交后代,细胞质来源于义和5号,导致雄性不育产生的可能原因:①芸薹属异源六倍体AABBCC 的细胞核基因与义和5号的细胞质基因互作结果;②金春的细胞核基因与义和5号的细胞质基因互作结果。但由于本试验仅用义和5号获得回交二代,鉴定的植株又少,目前很难判断雄性不育产生的原因。今后的工作重点:①继续进行回交转育,增加回交代数,并跟踪回交后代的育性;②着手配制(AABBCC×金春)×义和5号、金春×(AABBCC×义和5号)、(AABBCC× 义 和5号)× 金春、义和5号× 金春的多亲本杂交后代,通过调查各后代单株育性,进一步解释MS200712 雄性不育系产生的可能原因;③通过分子标记手段,开发鉴别雄性不育系的线粒体mtDNA(张扬勇 等,2011)、叶绿体cpDNA(张扬勇 等,2011;Wang et al.,2012)及核DNA 的SSR 等标记,进一步鉴定MS200712 雄性不育系类型。
本试验中,大白菜雄性不育系MS200712 的获取方法具有操作简单、重复性好等优点,细胞质雄性不育系的不育性稳定、败育彻底,且与其他大白菜品种杂交性良好、转育容易。因此,可以继续用于大白菜育种中的回交转育,以获得更多骨干雄性不育系材料。
今后,将开展有关大白菜雄性不育系MS200712 败育机理的研究,包括细胞学、分子生物学等方面研究,力争阐明大白菜雄性不育系MS200712 的所属类型及其败育机理,为该雄性不育系更好地在大白菜育种中应用及通过种间或属间杂交途径创造更多种类的雄性不育系奠定基础。
柯桂兰,赵稚雅,宋胭脂,张鲁刚,赵利民.1992.大白菜异源胞质雄性不育系CMS3411-7的选育与应用.园艺学报,19(4):333-340.
李殿荣.1984.甘蓝型油菜雄性不育系1763A 和6223A 的研究应用.陕西农业科学,(1):7-10.
张德双,徐家炳,曹鸣庆,张凤兰.2002.大白菜转育新型甘蓝型油菜细胞质雄性不育系的研究.华北农学报,17(1):60-63.
张德双.2006.大白菜CMS96 细胞质雄性不育的分子特性及育种应用研究〔博士论文〕.北京:中国农业科学院.
张德双,张凤兰,余阳俊,赵岫云,于拴仓,徐家炳.2009.芸薹属异源六倍体的叶片突变体的获得.中国蔬菜,(4):12-16.
张书芳,宋兆仲,赵雪云.1990.大白菜细胞核基因互作雄性不育系选育及应用模式.园艺学报,17(2):117-125.
张扬勇,方智远,王庆彪,刘玉梅,杨丽梅,庄木,孙培田.2011.两种甘蓝Ogura 细胞质雄性不育源的分子鉴别.中国农业科学,44(14):2959-2965.
Health D W,Earle E D,Dickson M H.1994.Introgressing coldtolerant Ogura cytoplasm from rapeseed into Pak Choi and Chinese cabbage.HortScience,29:202-203.
Shiga T,Baba S.1971.Cytoplasmic male sterility in rape plants (Brassica napusL.).Japan J Breed,21:15-17.
Shiga T,Baba S.1973.Cytoplasmic male sterility in oilseed rape (Brassica napusL.)and its utilization to breeding.Japan J Breed,23:187-197.
Thompson K F.1972.Cytoplasmic male sterility in oil-seed rape plants.Heredity,29:253-257.
Wang Q B,Zhang Y Y,Fang Z Y,Liu Y M,Yang L M,Zhuang M.2012.Chloroplast and mitochondrial SSR help to distinguish allocytoplasmic male sterile types in cabbage(BrassicaoleraceaL.var.capitata).Mol Breeding,30:709-716.
Williams P H.1981.The origin and development of cytoplasmic male sterile Chinese cabbage.Taiwan:Asian Vegetable Research and Development Center:293-300.
猜你喜欢
海洋通报(2022年4期)2022-10-10
麦类作物学报(2022年8期)2022-08-08
农业工程学报(2022年5期)2022-06-22
课外生活·趣知识(2022年2期)2022-02-08
中华戏曲(2020年2期)2020-02-12
高考·中(2019年6期)2019-09-10
蔬菜(2019年5期)2019-05-21
作物学报(2018年11期)2018-11-06
科学中国人(2016年30期)2016-07-14
中国蔬菜(2015年9期)2015-12-21
