
音乐和语言神经基础的重合与分离
—— 基于脑成像研究元分析的比较*
2014-02-06 03:20宋宜颖
心理学报 2014年3期
赖 寒 徐 苗 宋宜颖 刘 嘉,2
(1北京师范大学认知神经科学与学习国家重点实验室;2北京师范大学心理学院,北京 100875)
1 引言
音乐和语言都是人类最基本的能力之一,他们都是由声音构成,有特定的组织规则,可读写,并且能够传达特定含义。在不同的人类群体中,音乐和语言都是其文化的重要组成部分。日常生活中,我们利用语言来交流思想,抒发见解,同时我们也借助于音乐来表达情感,引发共鸣。音乐同语言在组织结构和功能上的相似,使得不同领域的研究者对二者关系的探讨从未停止过。本研究将试图从认知神经科学的角度,探讨音乐和语言在加工区域和机制上可能存在的联系和区别。
由于音乐和语言的加工过程都较为复杂,无论是初级的声音特征(声源、音高、音色)的分析,还是高级的句法(Syntax)和结构(Structure)加工,都影响着人们对音乐和语言的理解。为系统全面地描述和分析音乐和语言的关系,本文依照音乐和语言的层级结构,从各个加工阶段出发,探讨音乐和语言的关系。
以往的研究表明,无论是音乐还是语言,都是按照一定的规则组织起来的,二者均采用层级分析的形式进行加工(Bharucha&Krumhansl,1983;Patel,2003)。音高(Pitch)按照不同的组合形成了不同的和弦,和弦经过排列变换又形成了有调的旋律;同样的,词语通过组织形成了短语,而短语经过排列变换也形成了具有语法规则的句子。音乐和语言的这种较为类似的组织方式(Jackendoff&Lerdahl,2006),暗示二者可能存在联系。根据Koelsch等在2011年(Koelsch,2011)提出的音乐知觉加工模型,当物理的声音流进入人耳后,第一个加工阶段是对声音特征的提取阶段(Auditory feature extraction)。基底神经核团(如上/下丘、膝状体、丘脑)和初级听觉皮层通过对声音信息的解码和分析来提取并分析频率、音色、强度和声源等声音特征。第二个阶段是格式塔片段形成阶段(Gestalt formation)。此阶段根据格式塔规则,如相似性、接近性等原则,对频率类似、节奏类似或音色类似的声音特征进行重构,将声音特征组织为具有整体特性的格式塔片段。第三阶段为音程分析阶段(Interval analysis),即在格式塔片段的基础上分析和弦内部或旋律内部音(Pitch)与音的音程关系,以对旋律的音程进行表征。最后是对音乐结构的分析(Music structure analysis)。在这个阶段,大脑依据音调系统和旋律和谐等原则,通过对和弦(Chord)功能及其关系的分析,从而对音乐的旋律信息进行进一步的整合(Koelsch,2011;Koelsch&Siebel,2005)。对比Friederici等(Friederici,2002,2005)提出的语言知觉加工模型,语言知觉的加工也可大致分为四个阶段。第一个阶段与音乐知觉类似,即为声音特征的提取阶段。第二个阶段是语音分析阶段(Phonological analysis),主要负责词语语音边界的分割与组织,与音乐知觉的格式塔片段形成阶段类似。第三个阶段为词语分析阶段(Lexical analysis),即对词语的词形(Lexical form)和词义(Lexicosemantic)进行分析,并进行词语形音义的转换。最后是句子分析阶段(Sentence analysis),即在词语分析的基础上,完成句子的句法及其句义信息的加工和整合(Friederici,2002,2005)。对比音乐和语言的知觉加工模型发现,音乐和语言都是从声音特征的提取开始,通过层级加工的模式完成对旋律或语言的加工。以往对音乐和语言不同层级加工过程的研究进一步暗示了音乐和语言可能存在加工过程的关联。
已有研究表明,在低层级的语音加工阶段及较为低层级的音程分析和词义分析阶段,音乐和语言可能存在加工机制的重合。对音程分析的研究发现,让被试进行与音程分析有关的任务时,在颞上回(superior temporal gyrus,STG)(Brown&Martinez,2007;Foster&Zatorre,2010;Wehrum et al.,2011)、中央前回(precentral gyrus,Prec)(Brown&Martinez,2007;Foster&Zatorre,2010;Wehrum et al.,2011)、额下回的三角部(pars triangularis of the inferior frontalgyrus,IFGt)(Brown & Martinez,2007;Doeller et al.,2003;Schwenzer&Mathiak,2011)和脑岛(insular,Ins)(Foster&Zatorre,2010;Schwenzer&Mathiak,2011;Wehrum et al.,2011)等脑区都有明显激活。同时,大量研究表明,这些脑区也参与语言加工(Bitan,Lifshitz,Breznitz,&Booth,2010;Burman,Bitan,&Booth,2008;Kurth,Zilles,Fox,Laird,&Eickhoff,2010;Mutschler et al.,2009;Raettig,Frisch,Friederici,&Kotz,2010;Shuster&Lemieux,2005;Vigneau et al.,2006,2011)。一些探讨音乐与语言关系的研究进一步支持了音乐与语言可能存在共有加工机制这一观点。研究发现,早期的声调语言环境能够促进音程分析的发展(Stevens,Keller,&Tyler,2004),音程分析过程受损的乐盲,其对语言声调的加工也受损(Jiang,Hamm,Lim,Kirk,&Yang,2010);并且,当给被试同时呈现旋律与歌词时,在颞上回、颞中回(middle temporal gyrus,MTG)、脑岛和额下回等脑区,音乐的音程分析与语言的词义分析相互干扰(Schön et al.,2010)。也就是说,在相对于句法加工较为低层级的加工阶段,音乐和语言就已表现出了共同的加工机制。
并且,研究发现,在高层级的加工阶段(结构分析和句子分析),音乐与语言也可能存在共有的加工机制。有关旋律结构分析的研究表明,旋律结构分析所激活的颞上回(Foster&Zatorre,2010;Fujisawa&Cook,2011;Koelsch,Fritz,V Cramon,Müller,&Friederici,2005)、额中回(middle frontal gyrus,MFG)(Foster&Zatorre,2010;Fujisawa&Cook,2011;Schmithorst,2005)、额下回的岛盖部(pars opercularis of the inferior frontal gyrus,IFGo)(Foss,Altschuler,&James,2007;Fujisawa&Cook,2011;Koelsch,Fritz,Schulze,Alsop,&Schlaug,2005;Schmithorst,2005;Tillmann et al.,2006;Wehrum et al.,2011)、中央前回(Foss et al.,2007;Foster&Zatorre,2010;Fujisawa&Cook,2011;Koelsch,2005;Schmithorst,2005;Wehrum et al.,2011)、额极(frontal pole,Fpole)(Foss et al.,2007;Fujisawa&Cook,2011;Janata,Birk,et al.,2002;Koelsch,2005;Schmithorst,2005;Schulze,Mueller,&Koelsch,2011;Tillmann et al.,2006;Wehrum et al.,2011)和脑岛(Fujisawa&Cook,2011;Tillmann et al.,2006;Wehrum et al.,2011)等脑区也参与语言的加工(Binder,Desai,Graves,&Conant,2009;Bitan et al.,2010;Burman et al.,2008;Kurth et al.,2010;Mutschler et al.,2009;Pinel&Dehaene,2010;Raettig et al.,2010;Shuster& Lemieux,2005;Vigneau et al.,2006,2011)。同时,旋律的加工激活了与语言紧密联系的布洛卡区(额下回区域)和威尔尼克区(颞上回区域),这两个区域均参与了语言的句法加工,表明在句法层面上,音乐和语言也可能存在共有的加工机制(Schmithorst,2005)。行为和ERP的研究进一步支持了这一观点。研究发现,同时让被试进行音乐和语言的任务时,语言的句法加工会受到音乐句法加工的干扰(Fedorenko,Patel,Casasanto,Winawer,& Gibson,2009;Koelsch,2005)。语言句法加工能力受损的儿童其音乐句法加工的能力也受到损坏(Jentschke,Koelsch,Sallat,&Friederici,2008)。研究者通过颅内ERP定位发现,在早期句法错误侦测阶段,在颞下回区域,音乐和语言存在加工的重合(Sammler et al.,2009)。此外,还有研究发现,当给被试同时呈现旋律与歌词时,和谐的旋律会促进歌词句义的加工(Poulin-Charronnat,Bigand,Madurell,&Peereman,2005)。
然而,也有证据表明音乐和语言存在分离的加工机制。有关脑损伤的研究发现,失语症患者的言语能力受损时其旋律表达能力完好(Polk&Kertesz,1993),颞枕区中风患者的言语理解能力受损时其音乐能力也基本未受影响(Mendez,2001),而乐盲在其旋律加工能力受损时其语言能力也基本表现正常(Liu et al.,2012;Piccirilli,Sciarma,&Luzzi,2000;Polk&Kertesz,1993)。并且,对正常被试的脑成像研究也发现,虽然音乐和语言存在加工重合的脑区,但是其不重合的脑区也有很多(Rogalsky,Rong,Saberi,&Hickok,2011),甚至音乐的结构分析几乎不能激活相应句子分析的区域(Fedorenko,Behr,&Kanwisher,2011)。研究者据此认为,音乐和语言分属不同的模块,各有一套独特的加工机制。
音乐和语言是否同属同一个模块争论已久,之所以出现研究结果的不一致,可能是由于研究者所采用的研究范式不一致导致,也可能受其被试数量和分析方法的影响。鉴于此,为系统探讨音乐和语言加工的关系,本研究希望通过对比音乐和语言的元分析结果,从各个加工层级出发,探查音乐和语言加工的关系,并具体探讨音乐和语言在神经基础上的异同。
2 方法
2.1 数据提取和描述
由于已有研究者对特定于音乐知觉的两个层级,即较为低层级的音程分析和高层级的结构分析(赖寒,徐苗,宋宜颖,刘嘉,2013),以及语言知觉的三个层级,即低层级的语音分析、较为低层级的词义分析和高层级的句子分析(Vigneau et al.,2006,2011),做了较为完善的元分析,因此,本研究直接采用其结果,以进行后续的分析。研究中各层级元分析的文献搜索策略、以及所纳入文献的标准、文献总数、实验对比条件(Contrast)总数、峰值(Peaks)总数和最终聚类得到的簇(Cluster)总数具体如表1所示。
2.2 元分析结果的对比
由于元分析所得到的簇的中心坐标所依据的标准大脑模板均采用的是MNI(Montreal Neurological Institute)坐标系,因此,为了清楚的将音乐和语言各层级的加工机制进行对比,我们首先使用Caret软件(van Essen et al.,2001)将音乐和语言各层级加工所激活的簇的中心坐标投射到Caret自带的MNI模板上,即得到音乐和语言各层级加工的簇中心坐标对比图(图1)。
2.3 音乐和语言加工区域的重合
为了探查音乐和语言知觉的神经基础之间的关系,我们进一步分析了分属音乐和语言各层级加工的簇在大脑皮层上是否有重合。具体而言,我们将首先探查音程分析和语言各层级的簇在大脑皮层上是否有重合。具体而言,重合分析由以下四个步骤构成。首先,我们将音程分析与语言各层级的簇两两随机进行配对,并计算出每对簇的中心坐标的欧式距离,筛选出簇间间距小于7 mm(约3个体素以内)的配对。随后,我们将筛选出的配对中的两个簇的峰值混合,采用K-means聚类分析方法,指定聚类数目为K=2,迭代次数T=10。若重新聚类后,两簇的峰值混合在一起,则从描述上表明这两个簇所在的脑区上可能有重合。再次,为了从统计上进一步验证配对中的两个簇是否存在重合,我们对由K-means聚类筛选出的配对做了多元方差分析。具体而言,我们对配对中的两个簇的峰值在X、Y、Z方向上分别进行了多元方差分析。若配对的簇在X、Y、Z方向上的组间差异均不显著,则可认为这两个簇所在的脑区上有重合(Reich,Szwed,Cohen,&Amedi,2011)。最后,为了进一步量化参与音程分析和语言加工的簇在多大程度上有重合,我们分别计算了其左右脑的簇重合率,计算方法为音程分析左(右)脑的重合簇总数除以音程分析左(右)脑的簇总数。同样的流程被采用来寻找结构分析与语言各层级加工有重合的簇。

表1 音乐和语言各层级元分析的详细信息
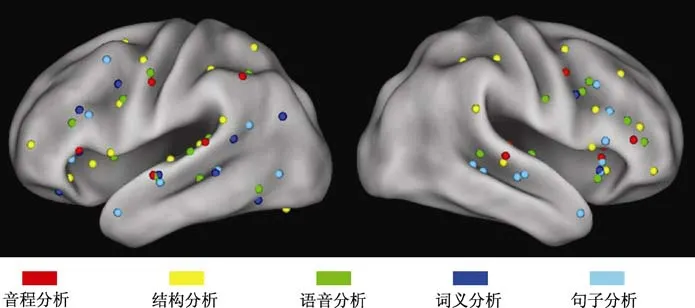
图1 音乐和语言各层级加工的簇中心坐标对比图
3 结果
首先,为描述性地考察音乐和语言各层级加工所激活的区域在大脑皮层上是否可能有重合,我们将音乐和语言的元分析研究所得到的簇中心坐标,标记在Caret自带的MNI模板上。结果如图1所示,音乐的两个层级,音程分析(红色的点)和结构分析(黄色的点)所激活的簇,与语言的三个层级,语音分析(绿色的点)、词义分析(蓝色的点)和句子分析(青色的点)所激活的簇,在左脑的额叶、颞叶和顶叶以及右脑的额叶和脑岛等区域,均比较接近,暗示音乐和语言可能存在共享的神经基础。
为了进一步探查音乐和语言知觉的神经基础之间的关系,我们分别对分属音乐和语言各层级加工的簇进行了重合分析。
具体而言,对于音程分析,我们筛选出了5个簇间间距小于7 mm的配对。进一步分别对筛选出来的5个配对做K-means聚类分析,我们发现,这5个配对的簇的峰值均混合在一起,表明这5个配对可能有重合。再次,为了从统计上进一步验证配对中的两个簇是否有重合,我们对由K-means聚类筛选出的5个配对做了多元方差分析。结果发现,这5个配对的簇中心坐标在X、Y、Z方向上的组间差异均不显著,表明了这5个配对的两个簇所在的脑区的重合。
各配对的簇的分布及坐标位置具体如图2所示,各配对的簇的中心坐标以及其在X、Y、Z方向上的F值与p
值具体如表2所示。其中,参与音程分析的簇,与参与语言三个层级加工的簇,两两间均存在重合。其中,它与语音分析发生重合的区域位于左侧颞上回和左侧中央前回;与词义加工发生重合的区域位于左侧颞上回和左侧额下回的三角部;与句子分析发生重合的区域位于右侧脑岛。通过计算左右脑的簇重合率发现,音程分析与语言加工重合较多,主要表现在大脑左侧(50%)有较多重合。对于结构分析,我们筛选出了6个簇间间距小于7 mm的配对。进一步分别对筛选出来的6个配对做K-means聚类分析,我们发现,这6个配对的簇的峰值均混合在一起,表明这6个配对可能有重合。再次,为了从统计上进一步验证配对中的两个簇是否有重合,我们对由K-means聚类筛选出的6个配对做了多元方差分析。结果发现,除了一对配对,即结构分析的簇,中央前回,与语音分析的簇,中央前回,在Y方向上的簇峰值坐标存在显著差异外(p
=0.04),其余5个配对的簇中心坐标在X、Y、Z方向上的组间差异均不显著,表明了这5个配对的两个簇所在的脑区的重合。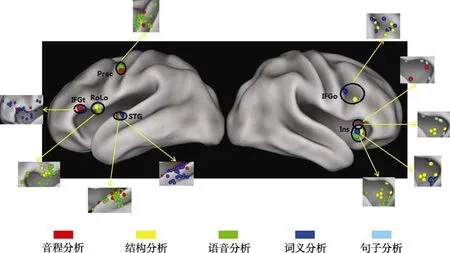
图2 音乐和语言各层级加工的重合
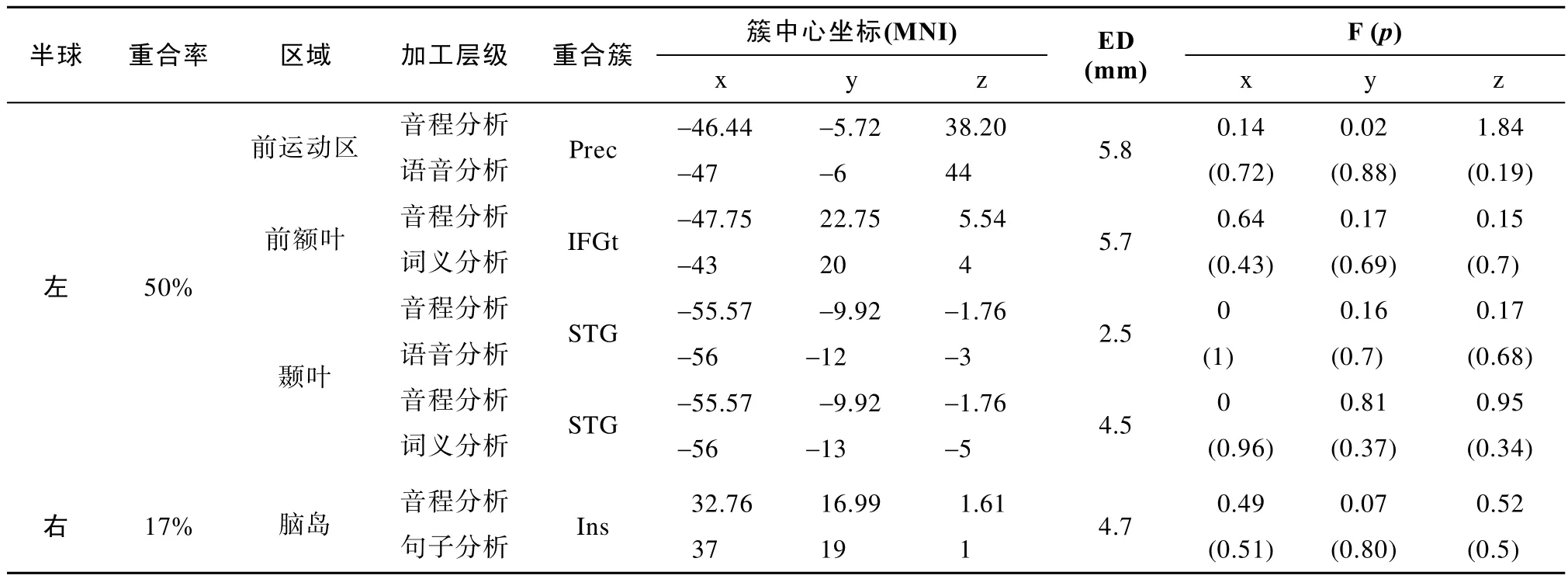
表2 音程分析和语言各层级加工的重合率、重合脑区坐标、簇间距离及F检验统计值
各配对的簇的分布及坐标位置具体如图2所示,各配对的簇的中心坐标以及其在X、Y、Z方向上的F值与p
值具体如表3所示。其中,参与结构分析的簇,与参与语言三个层级加工的簇,两两间均存在重合。其中,它与语音分析发生重合的区域位于左侧额盖区(Rolandic operculum,RoLo)和右侧脑岛;与词义分析发生重合的区域位于右侧额下回的岛盖部和右侧脑岛;与句子分析发生重合的区域位于右侧脑岛。通过计算左右脑的簇重合率发现,结构分析与语言加工重合较少,在大脑左侧(7%)和右侧(14%)均只存在少量重合。4 讨论
本研究根据音乐和语言的层级结构,从各个加工阶段出发,探讨音乐和语言的关系。具体而言,我们对二者已有的元分析结果进行对比,进而探讨了音乐和语言在神经基础上的异同。结果表明,音乐的两个层级,音程分析和结构分析,与语言的三个层级,语音分析、词义分析和句子分析,两两间均存在加工重合的区域。音程分析和语言加工重合更多,在大脑左侧有较多重合(50%),而结构分析与语言加工重合较少,在大脑左侧(7%)和右侧(14%)均只有少量重合,暗示了在较为低层级的加工阶段,音乐和语言更可能存在共享的神经基础,而在更为高层级的加工阶段,音乐和语言的加工更多地表现出各自独特的特点。
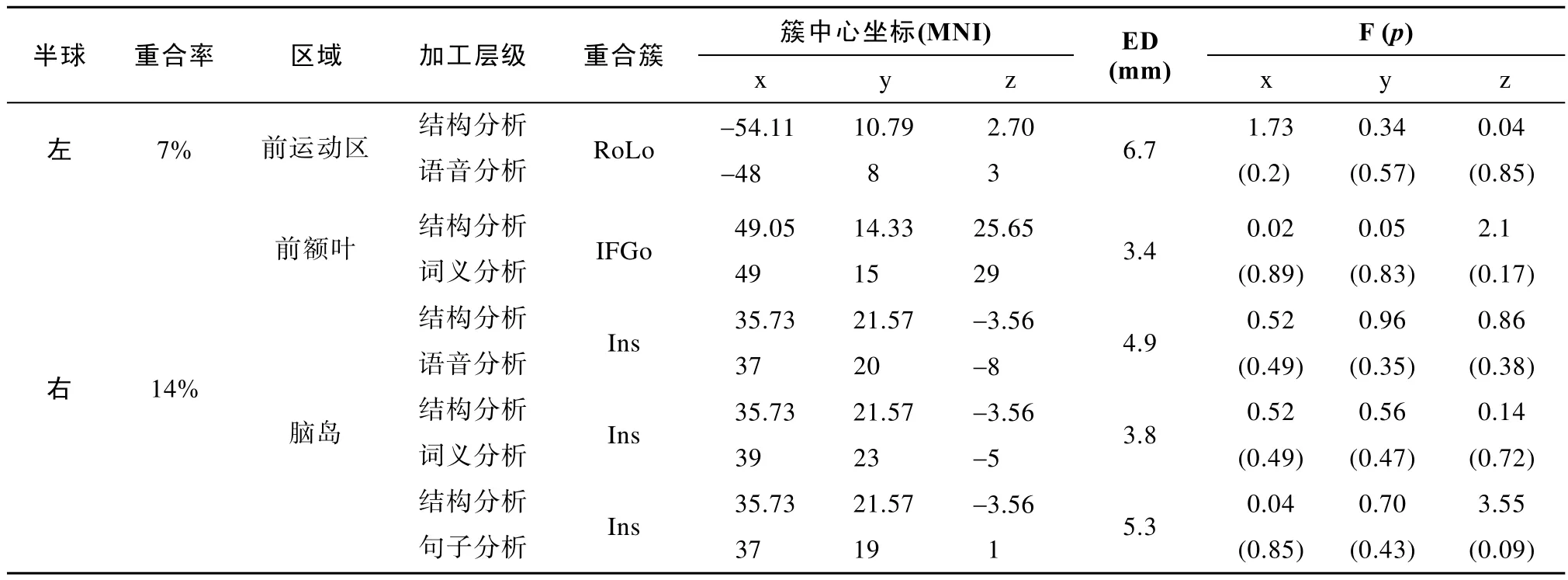
表3 结构分析和语言各层级加工的重合率、重合脑区坐标、簇间距离及F检验统计值
具体而言,音程分析与语言三个层级的重合区域主要分布在左侧颞叶(颞上回)、左侧前运动区(中央前回)、左侧前额叶(额下回的三角部)和右侧脑岛;结构分析与语言三个层级的重合区域主要分布在左侧前运动区(额盖区)、右侧前额叶(额下回的岛盖部)和右侧脑岛(图3)。这些区域可能为音乐知觉与语言知觉各层级加工的神经环路的共有节点,暗示了二者在听觉—运动环路、核心环路和认知—情绪环路上可能存在的联系。下面,我们将在音乐知觉和语言知觉的神经环路的基础上,详细探讨音乐和语言的关系。
4.1 音乐和语言知觉在神经环路上的重合
4.1.1 音乐与语音分析在“听觉—运动环路”存在重合节点
已有研究表明,音程分析和结构分析均需要听觉—运动环路的参与,而此在语音分析中也有涉及(Vigneau et al.,2006,2011;赖寒等,2013)。本结果表明,音程分析和结构分析均可能与语音分析在听觉—运动环路存在重合节点。不同的是,音程分析与语音加工的重合节点主要分布在左侧中央前回,而结构分析与语音加工的重合节点则主要分布在左侧额盖区(图4)。
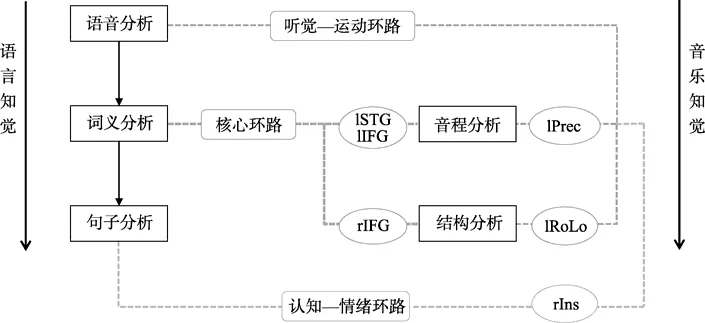
图3 音乐和语言知觉示意图

图4 音程分析、结构分析与语音分析的的各神经环路关系图
具体而言,对于音程分析,其与语音分析在左侧中央前回存在重合,表明二者可能存在听觉—运动整合上的联系。已有研究表明,位于前运动皮层的中央前回与音乐和语言的听觉—运动整合有关(Bangert et al.,2006;Hyde et al.,2009;Pulvermüller&Fadiga,2010;Vigneau et al.,2006),主要负责口舌和鄂的运动(Mohammadietal.,2009;Pulvermülleretal.,2006;Schubotz,Anwander,Knösche,Von Cramon,&Tittgemeyer,2010;Wong,Dzemidzic,Talavage,Romito,&Byrd,2011),参与音程分析和语音分析听觉—运动环路的加工(Vigneau et al.,2006;赖寒等,2013)。因此,我们认为,左侧中央前回可能为音程分析和语音分析在听觉—运动环路的共有节点。在音程分析阶段,左侧中央前回主要担任音程—运动联合区的角色,整合音程—运动的加工,以帮助重复和记忆旋律信息,反映了旋律加工中所存在的默唱的加工过程(赖寒等,2013);而在语音分析阶段,左侧中央前回则主要担任语音—运动联合区的角色,整合语音—运动的加工,帮助重复和记忆语音信息,反映了语音加工中所存在的默读的加工过程(Szenkovits,Peelle,Norris,&Davis,2012;Vigneau et al.,2006)。
同样,结构分析与语音分析在左侧额盖区存在重合,也表面二者可能存在听觉—运动整合上的联系。位于前运动区的额盖区也与音乐和语言的听觉—运动整合有关(Kell et al.,2009;Koelsch et al.,2009;Szenkovits et al.,2012;Vigneau et al.,2006),主要负责喉结的运动控制(Brown,Ngan,&Liotti,2008;Chang,Erickson,Ambrose,Hasegawa-Johnson,&Ludlow,2008;Jäncke,Hänggi,&Steinmetz,2004),参与结构分析和语音分析听觉—运动环路的加工(Vigneau et al.,2006;赖寒等,2013)。因此,我们认为,左侧额盖区可能为结构分析和语音分析在听觉—运动环路的共有结点。在结构分析阶段,左侧额盖区主要担任旋律—运动联合区的角色,整合旋律—运动的加工;而在语音分析阶段,左侧额盖区则和左侧中央前回一起,担任语音—运动联合区的角色,整合语音—运动的加工。
4.1.2 音乐与词义分析在“核心环路”存在重合节点
已有研究表明,左侧颞上回与左侧额下回均参与音程分析核心环路的加工,右侧额下回参与结构分析核心环路的加工(赖寒等,2013);这些区域在词义加工的核心环路之一,背侧语义环路中均有涉及(Friederici,2012;Hickok & Poeppel,2004;Vigneau et al.,2006,2011;赖寒等,2013)。本研究结果表明,音程分析和结构分析均可能与词义分析在其核心环路存在重合节点。不同的是,音程分析与词义分析的重合节点主要分布在左侧颞上回与左侧额下回,而结构分析与词义分析的重合节点主要分布在右侧额下回(图5)。
具体而言,对于音程分析,其与词义分析在左侧颞上回存在重合,表明二者可能存在听觉加工上的联系。左侧颞上回可能为连接音程分析核心环路和词义分析背侧语义环路的共有节点。在音程分析阶段,左侧颞上回主要负责旋律轮廓、和弦与旋律音程变化的知觉以及相对音高关系的精细分析(赖寒等,2013);而在词义分析阶段,左侧颞上回主要担任词语语音、词形和词义的中转站的角色,负责整合词语的语音、词形和词义信息,将所知觉的字母和字形转换成音节形式,并将语音信息转换成相应的语义表征,进而促进词语的概念加工和后续的句子加工(Friederici,2012;Houdé,Rossi,Lubin,&Joliot,2010;Pulvermüller&Fadiga,2010;Vigneau et al.,2006)。此外,音程分析还与词义分析在左侧额下回存在重合,表明二者还存在工作记忆加工上的联系。左侧额下回可能与左侧颞上回一起,共同作为连接音程分析核心环路和词义分析背侧语义环路的节点。在音程分析阶段,左侧额下回负责对和弦及旋律信息进行保持与存储,为颞上回完成音程分析奠定基础(赖寒等,2013);而在词义分析阶段,左侧额下回则主要负责对词语的抽象词义进行判断、选择与提取,为后续的词义整合及句子分析奠定基础(Moss et al.,2005;Vigneau et al.,2006;Whitney,Kirk,O'Sullivan,Ralph,& Jefferies,2011)。
同样,结构分析与词义分析在右侧额下回存在重合,表明二者也可能存在工作记忆加工上的联系。已有研究表明,右侧额下回的岛盖部与执行功能有关(Boehler,Appelbaum,Krebs,Hopf,&Woldorff,2010;Brown et al.,2012;Hampshire,Chamberlain,Monti,Duncan,&Owen,2010;Vigneau et al.,2011),在旋律结构和词语加工等工作记忆有关的任务中均有激活,既参与结构工作记忆环路的加工(Janata,Tillmann,et al.,2002;赖寒等,2013),又辅助语言各层级工作记忆环路的加工(Vigneau et al.,2011)。因此,我们认为,右侧额下回的岛盖部可能为连接结构分析核心环路和词义分析背侧语义环路的共有节点。在结构分析阶段,右侧额下回的岛盖部与前额叶其他区域一起,负责对颞上回整合而来的声音序列信息进行注意、保持与存储,以助于旋律句法结构的建构及实验任务的完成(赖寒等,2013);而在词义分析阶段,右侧额下回的岛盖部则主要负责对词义信息的注意、保持及存储过程,以助于词义提取及实验任务的完成(Vigneau et al.,2011)。
4.1.3 音乐与句子分析在“认知—情绪环路”存在重合节点
以往研究表明,音程分析和结构分析均需要认知—情绪环路的参与,而此在句子分析中也有涉及(Vigneau et al.,2006,2011;赖寒等,2013)。本研究结构表明,音程分析和结构分析均可能与句子分析在认知—情绪环路存在重合节点,且重合节点均分布于右侧脑岛(图6)。
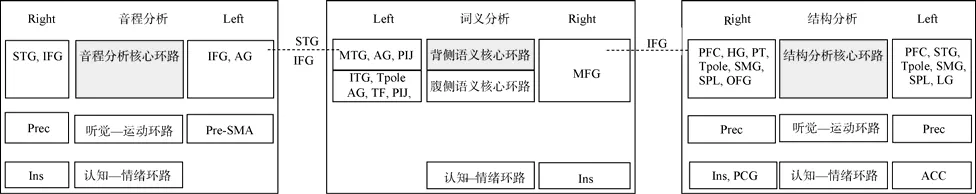
图5 音程分析、结构分析与词义分析的的各神经环路关系图
已有研究表明,无论是音乐知觉(Han,Rho,Jun,&Hwang,2010;Huq,Bello,&Rowe,2010;Kim et al.,2010;Omar et al.,2011;Yang,Lin,Su,&Chen,2008),还是语言知觉(Jones,Ward,&Critchley,2010;Mutschler et al.,2009),均会引发个体情绪体验的产生,并且这种情绪体验的加工大多需要脑岛的参与(Boso,Politi,Barale,&Emanuele,2006;Jones et al.,2010;Koelsch,2010;Koelsch&Siebel,2005)。对音乐和语言知觉的元分析发现,右侧脑岛不仅参与音程分析和结构分析的认知—情绪环路的加工(赖寒等,2013),也参与句子分析认知—情绪环路的加工(Kurth et al.,2010)。
因此,我们认为,右侧脑岛可能为音程分析和结构分析分别与句子分析在认知—情绪环路的共有节点。在音程分析和结构分析阶段,脑岛参与旋律音程分析和旋律结构分析的情绪唤起过程,而在句子分析阶段,脑岛参与句子分析的情绪唤起过程。
4.2 音乐和语言知觉在神经环路上的分离
然而,虽然音乐和语言知觉在神经环路上存在共有的节点,但是二者不重合的节点则更多,二者的加工表现出各自独特的特点,在神经环路上存在以下三个方面的区别。
第一、音乐和语言知觉的神经环路不尽相同(图4-6)。例如,音乐与语言知觉均包含各自独特的核心环路。虽然音程分析和结构分析的核心环路与词义分析的背侧语义环路分别存在着共有节点,但是环路内二者不重合的节点更多,可能反映了各自独特的加工机制。
第二、虽然音乐和语言知觉均包含了相同的神经环路,但是二者所涉及的脑区绝大多数并不相同(图4-6)。并且,即使有的脑区名字相同,但是他们却不是重合在一起的,可能反映了他们在功能上的侧重不一样。例如,结构分析与句子分析均包含认知—情绪环路,但结构分析的认知—情绪环路主要依靠脑岛、扣带前回、旁扣带回完成,而句子分析主要依靠脑岛完成。又如音程分析和语音分析的听觉—运动环路均包含右侧中央前回,然而二者在右侧中央前回却并未发生重合。
第三、神经环路的分离更多地发生在更为高层级的加工阶段。对于音乐知觉,与较为低层级的音程分析相比,结构分析在听觉—运动环路、核心环路和认知—情绪环路内,均与语言加工存在更多不重合节点。且对于语言知觉,与低层级的语音分析及较为低层级的词义分析相比,句子分析只在外围的认知—情绪环路与音乐知觉发生重合,在句子分析核心环路则并未与音乐知觉发生重合。这可能反映出音乐和语言所特有的加工过程更可能发生在更为高层级的加工阶段。
5 展望
本研究通过对比音乐和语言的元分析结果,从各个加工层级出发,首次对音乐和语言的神经基础的关系进行了全面地分析,从而探讨了音乐和语言在神经基础上的异同。
本研究认为,音乐和语言的各层级间均存在共有的神经环路节点,分别位于左侧颞叶(颞上回)、左侧前运动区(中央前回和额盖区)、双侧前额叶(左侧额下回的三角部和双侧额下回的岛盖部)和右侧脑岛。并且,音乐和语言共有的神经基础更可能存在于较为低层级的加工阶段,而在更为高层级的加工阶段,音乐和语言的加工更多地表现出各自独特的特点。然而,本研究的结果只是基于已有音乐和语言元分析结果的对比,还需要更为科学和严格的方法加以验证。未来研究有必要在同一批被试中直接比较音乐和语言各层级加工的关系,看看哪些脑区特异于音乐和语言知觉,哪些脑区是音乐和语言知觉所共享,严格探查其共享及分离的神经基础具体发生于音乐和语言知觉的哪一层级。同时,由于音程分析和结构分析是一个相对粗略的加工过程,音程分析包含了旋律轮廓和音程两类信息的加工,结构分析包含了和旋和谐、旋律和旋和音调结构等信息的加工。因此,随着相关fMRI研究量的增加,在未来的元分析中,有必要对二者的加工层级进行进一步细分,并在其与语言知觉所对应的层级上,更为精细的比较音乐知觉和语言知觉的关系。
此外,对比音乐和语言的元分析结果发现,不仅音乐和语言的各层级间均存在共有的神经环路节点,而且在不同层级,其重合的神经环路不尽相同。由于音乐训练相较于语言训练更方便也更简单,已有大量研究将音乐训练应用于提高个体的语言能力(Jäncke,2012;Moreno&Besson,2006;Moreno et al.,2009;Paul,Sharda,&Singh,2012;Schön&François,2011),因此,本研究不仅为其提供了理论支持,而且对如何将音乐训练更为有效的运用于语言发展提供了线索。例如,本研究发现,语音分析与音乐旋律分析在听觉—运动环路存在共有节点,那么对于在语音加工方面存在障碍的个体,则可以通过器乐和歌唱等训练加强听觉—运动整合能力,由此提高诵读等语音能力,实现个体语音能力的改善。又如,本研究发现,结构分析和词义分析在核心环路存在工作记忆加工过程的联系,那么对于在词义加工方面存在障碍的个体,则可以通过加强其结构分析的工作记忆能力提高其词义分析的工作记忆能力,从而改善个体的词义加工。
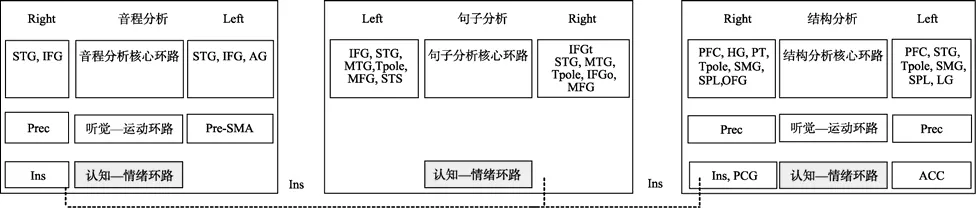
图6 音程分析、结构分析与句子分析的各神经环路关系图,虚线表示联系
Bangert,M.,Peschel,T.,Schlaug,G.,Rotte,M.,Drescher,D.,Hinrichs,H.,...Altenmüller,E.(2006).Shared networks for auditory and motor processing in professional pianists:Evidencefrom fMRIconjunction.Neuroimage,30
(3),917–926.Bharucha,J.,&Krumhansl,C.L.(1983).The representation of harmonic structure in music:Hierarchies of stability as a function of context.Cognition,13
(1),63–102.Binder,J.R.,Desai,R.H.,Graves,W.W.,&Conant,L.L.(2009).Where is the semantic system?A critical review and meta-analysis of 120 functional neuroimaging studies.Cerebral Cortex,19
(12),2767–2796.Bitan,T.,Lifshitz,A.,Breznitz,Z.,&Booth,J.R.(2010).Bidirectional connectivity between hemispheres occurs at multiple levels in language processing but depends on sex.The Journal of Neuroscience,30
(35),11576–11585.Boehler,C.N.,Appelbaum,L.G.,Krebs,R.M.,Hopf,J.M.,& Woldorff,M.G.(2010).Pinning down response inhibition in the brain—Conjunction analyses ofthe Stop-signal task.Neuroimage,52
(4),1621–1632.Boso,M.,Politi,P.,Barale,F.,&Emanuele,E.(2006).Neurophysiology and neurobiology of the musical experience.Functional Neurology,21
(4),187–191.Brown,M.R.G.,Lebel,R.M.,Dolcos,F.,Wilman,A.H.,Silverstone,P.H.,Pazderka,H.,...Dursun,S.M.(2012).Effects of emotional context on impulse control.Neuroimage,63
,434–446.Brown,S.,&Martinez,M.J.(2007).Activation of premotor vocal areas during musical discrimination.Brain Cogn,63
(1),59–69.Brown,S.,Ngan,E.,&Liotti,M.(2008).A larynx area in the human motor cortex.Cerebral Cortex,18
(4),837–845.Burman,D.D.,Bitan,T.,& Booth,J.R.(2008).Sex differences in neuralprocessing of language among children.Neuropsychologia,46
(5),1349–1362.Chang, S.-E., Erickson, K. I., Ambrose, N. G.,Hasegawa-Johnson,M.A.,&Ludlow,C.L.(2008).Brain anatomy differences in childhood stuttering.Neuroimage,39
(3),1333–1344.Doeller,C.F.,Opitz,B.,Mecklinger,A.,Krick,C.,Reith,W.,&Schröger,E.(2003).Prefrontal cortex involvement in preattentive auditory deviance detection:Neuroimaging and electrophysiologicalevidence.Neuroimage,20
(2),1270–1282.Fedorenko,E.,Behr,M.K.,& Kanwisher,N.(2011).Functional specificity for high-level linguistic processing in the human brain.Proceedings of the National Academy of Sciences of the United States of America,108
(39),16428–16433.Fedorenko,E.,Patel,A.,Casasanto,D.,Winawer,J.,&Gibson,E.(2009).Structural integration in language and music:Evidence for a shared system.Memory&Cognition,37
(1),1–9.Foss,A.H.,Altschuler,E.L.,&James,K.H.(2007).Neural correlates of the Pythagorean ratio rules.Neuroreport,18
(15),1521–1525.Foster,N.E.V.,&Zatorre,R.J.(2010).A role for the intraparietal sulcus in transforming musical pitch information.Cerebral Cortex,20
(6),1350–1359.Friederici,A.D.(2002).Towards a neural basis of auditory sentence processing.Trends in Cognitive Sciences,6
(2),78–84.Friederici,A.D.(2005).Neurophysiological markers of early language acquisition:From syllables to sentences.Trends in Cognitive Sciences,9
(10),481–488.Friederici,A.D.(2012).The cortical language circuit:From auditory perception to sentence comprehension.Trends in Cognitive Sciences,16
,262–268.Fujisawa,T.X.,&Cook,N.D.(2011).The perception of harmonictriads:An fMRIstudy.Brain
Imaging
and Behavior,5
(2),109–125.Hampshire,A.,Chamberlain,S.R.,Monti,M.M.,Duncan,J.,&Owen,A.M.(2010).The role of the right inferior frontal gyrus:Inhibition and attentionalcontrol.Neuroimage,50
(3),1313–1319.Han,B.J.,Rho,S.,Jun,S.,&Hwang,E.(2010).Music emotion classification and context-based music recommendation.Multimedia Tools and Applications,47
(3),433–460.Hickok,G.,&Poeppel,D.(2004).Dorsal and ventral streams:A framework for understanding aspects of the functional anatomy of language.Cognition,92
(1-2),67–99.Houdé,O.,Rossi,S.,Lubin,A.,&Joliot,M.(2010).Mapping numerical processing,reading,and executive functions in the developing brain:An fMRI meta-analysis of 52 studies including 842 children.DevelopmentalScience,13
(6),876–885.Huq,A.,Bello,J.P.,&Rowe,R.(2010).Automated music emotion recognition:A systematic evaluation.Journal of New Music Research,39
(3),227–244.Hyde,K.L.,Lerch,J.,Norton,A.,Forgeard,M.,Winner,E.,Evans,A.C.,&Schlaug,G.(2009).Musical training shapes structuralbrain development.The
Journalof Neuroscience,29
(10),3019–3025.Jackendoff,R.,&Lerdahl,F.(2006).The capacity for music:What is it,and what's special about it?Cognition,100
(1),33–72.Janata,P.,Birk,J.L.,van Horn,J.D.,Leman,M.,Tillmann,B.,&Bharucha,J.J.(2002).The cortical topography of tonal structures underlying Western music.Science,298
(5601),2167–2170.Janata,P.,Tillmann,B.,&Bharucha,J.J.(2002).Listening to polyphonic music recruits domain-general attention and working memory circuits.Cognitive,
Affective,
&Behavioral Neuroscience,2
(2),121–140.Jäncke,L.(2012).Therelationship between musicand language.Frontiers in Psychology,3
,123.Jäncke,L.,Hänggi,J.,&Steinmetz,H.(2004).Morphological brain differences between adult stutterers and non-stutterers.BMC Neurology,4
(1),23.Jentschke,S.,Koelsch,S.,Sallat,S.,&Friederici,A.D.(2008).Children with specific language impairment also show impairment of music-syntactic processing.Journal of Cognitive Neuroscience,20
(11),1940–1951.Jiang,C.M.,Hamm,J.P.,Lim,V.K.,Kirk,I.J.,&Yang,Y.F.(2010).Processing melodic contour and speech intonation in congenital amusics with Mandarin Chinese.Neuropsychologia,48
(9),2630–2639.Jones,C.L.,Ward,J.,&Critchley,H.D.(2010).The neuropsychological impact of insular cortex lesions.Journal of Neurology,Neurosurgery&Psychiatry,81
(6),611–618.Kell,C.A.,Neumann,K.,von Kriegstein,K.,Posenenske,C.,von Gudenberg,A.W.,Euler,H.,&Giraud,A.-L.(2009).How the brain repairs stuttering.Brain,
132
(10),2747–2760.Kim,Y.E.,Schmidt,E.M.,Migneco,R.,Morton,B.G.,Richardson,P.,Scott,J.,...Turnbull,D.(2010).Music emotion recognition:A state of the art review.ISMIR,Utrecht,Netherlands.
Koelsch,S.(2005).Neural substrates of processing syntax and semantics in music.Current Opinion in Neurobiology,15
(2),207–212.Koelsch,S.(2010).Towards a neural basis of music-evoked emotions.Trends in Cognitive Sciences,14
(3),131–137.Koelsch,S.(2011).Toward a neural basis of music perception-a review and updated model.Frontiers in Psychology,2
,110.Koelsch,S.,Fritz,T.,Schulze,K.,Alsop,D.,&Schlaug,G.(2005).Adults and children processing music:An fMRI study.Neuroimage,25
(4),1068–1076.Koelsch,S.,Fritz,T.,V Cramon,D.Y.,Müller,K.,&Friederici,A.D.(2005).Investigating emotion with music:An fMRI study.Human Brain Mapping,27
(3),239–250.Koelsch,S.,Schulze,K.,Sammler,D.,Fritz,T.,Müller,K.,&Gruber,O.(2009).Functional architecture of verbal and tonal working memory:An fMRI study.Human Brain Mapping,30
(3),859–873.Koelsch,S.,&Siebel,W.A.(2005).Towards a neural basis of music perception.Trends in Cognitive Sciences,9
(12),578–584.Kurth,F.,Zilles,K.,Fox,P.T.,Laird,A.R.,&Eickhoff,S.B.(2010). A link between the systems: Functional differentiation and integration within the human insula revealed by meta-analysis.Brain Structure and Function,214
(5-6),519–534.Lai,H.,Xu,M.,Song,Y.Y.,&Liu,J.(2013).The neural mechanism underlying music perception:A meta-analysis of fMRI studies.Acta Psychologica Sinica,45
(5),491–507.[赖寒,徐苗,宋宜颖,刘嘉.(2013).音乐知觉的神经基础:脑成像研究的元分析.心理学报,45
(5),491–507.]Liu,F.,Jiang,C.M.,Thompson,W.F.,Xu,Y.,Yang,Y.F.,&Stewart,L.(2012).The mechanism of speech processing in congenitalamusia:Evidencefrom mandarin speakers.PLoS One,7
(2),e30374.Mendez,M.F.(2001).Generalized auditory agnosia with spared music recognition in a left-hander.Analysis of a case with a right temporal stroke.Cortex,37
(1),139–150.Mohammadi,B.,Kollewe,K.,Samii,A.,Krampfl,K.,Dengler,R.,&Münte,T.F.(2009).Decreased brain activation to tongue movements in amyotrophic lateral sclerosis with bulbar involvement but not Kennedy syndrome.Journal of Neurology,256
(8),1263–1269.Moreno,S.,&Besson,M.(2006).Musical training and language-related brain electrical activity in children.Psychophysiology,43
(3),287–291.Moreno,S.,Marques,C.,Santos,A.,Santos,M.,Castro,S.L.,&Besson,M.(2009).Musical training influences linguistic abilities in 8-year-old children:More evidence for brain plasticity.Cerebral Cortex,19
(3),712–723.Moss,H.E.,Abdallah,S.,Fletcher,P.,Bright,P.,Pilgrim,L.,Acres,K.,& Tyler,L.K.(2005).Selecting among competing alternatives:Selection and retrieval in the left inferior frontal gyrus.Cerebral Cortex,15
(11),1723–1735.Mutschler,I.,Wieckhorst,B.,Kowalevski,S.,Derix,J.,Wentlandt,J.,Schulze-Bonhage,A.,&Ball,T.(2009).Functional organization ofthehuman anterior insular cortex.Neuroscience Letters,457
(2),66–70.Omar,R.,Henley,S.M.D.,Bartlett,J.W.,Hailstone,J.C.,Gordon,E.,Sauter,D.A.,...Warren,J.D.(2011).The structural neuroanatomy of music emotion recognition:Evidence from frontotemporal lobar degeneration.Neuroimage,56
(3),1814–1821.Patel,A.D.(2003).Language,music,syntax and the brain.Nature Neuroscience,6
(7),674–681.Paul,A.,Sharda,M.,&Singh,N.C.(2012).Effect of music instruction on cognitive development:A review.Journal of the Indian Institute of Science,92
(4),441–446.Piccirilli,M.,Sciarma,T.,&Luzzi,S.(2000).Modularity of music:Evidence from a case of pure amusia.Journal of Neurology,Neurosurgery&Psychiatry,69
(4),541–545.Pinel,P.,& Dehaene,S.(2010).Beyond hemispheric dominance:Brain regions underlying the joint lateralization of language and arithmetic to the left hemisphere.Journal of Cognitive Neuroscience,22
(1),48–66.Polk,M.,&Kertesz,A.(1993).Music and language in degenerative disease of the brain.Brain and Cognition,22
(1),98–117.Poulin-Charronnat,B.,Bigand,E.,Madurell,F.,&Peereman,R.(2005).Musical structure modulates semantic priming in vocal music.Cognition,94
(3),B67–B78.Pulvermüller,F.,&Fadiga,L.(2010).Active perception:Sensorimotor circuits as a cortical basis for language.Nature Reviews Neuroscience,11
(5),351–360.Pulvermüller,F.,Huss,M.,Kherif,F.,del Prado Martin,F.M.,Hauk,O.,& Shtyrov,Y.(2006).Motor cortex maps articulatory features of speech sounds.Proceedings of the National Academy of Sciences of the United States of America,103
(20),7865–7870.Raettig,T.,Frisch,S.,Friederici,A.D.,&Kotz,S.A.(2010).Neural correlates of morphosyntactic and verb-argument structureprocessing:An EfMRIstudy.Cortex,46
(5),613–620.Reich,L.,Szwed,M.,Cohen,L.,&Amedi,A.(2011).A ventral visual stream reading center independent of visual experience.Current Biology,21
(5),363–368.Rogalsky,C.,Rong,F.,Saberi,K.,&Hickok,G.(2011).Functional anatomy of language and music perception:Temporal and structural factors investigated using functional magnetic resonance imaging.Journal
of Neuroscience,31
(10),3843–3852.Sammler,D.,Koelsch,S.,Ball,T.,Brandt,A.,Elger,C.E.,Friederici,A.D.,...Schulze-Bonhage,A.(2009).Overlap of musical and linguistic syntax processing:Intracranial ERP evidence.Annals of the New York Academy of Sciences,1169
(1),494–498.Schmithorst,V.J.(2005).Separate cortical networks involved in music perception:Preliminary functional MRI evidence for modularity of music processing.Neuroimage,25
(2),444–451.Schön,D.,& François,C.(2011).Musical expertise and statistical learning of musical and linguistic structures.Frontiers in Psychology,2
,167.Schön,D.,Gordon,R.,Campagne,A.,Magne,C.,Astésano,C.,Anton,J.L.,&Besson,M.(2010).Similar cerebral networks in language, music and song perception.Neuroimage,51
(1),450–461.Schubotz,R.I.,Anwander,A.,Knösche,T.R.,Von Cramon,D.Y.,&Tittgemeyer,M.(2010).Anatomical and functional parcellation of the human lateral premotor cortex.Neuroimage,50
(2),396–408.Schulze,K.,Mueller,K.,& Koelsch,S.(2011).Neural correlates of strategy use during auditory working memory in musicians and non-musicians.European Journal of Neuroscience,33
(1),189–196.Schwenzer,M.,&Mathiak,K.(2011).Numeric aspects in pitch identification:An fMRI study.BMC Neuroscience,12
,26.Shuster,L.I.,&Lemieux,S.K.(2005).An fMRI investigation of covertly and overtly produced mono-and multisyllabic words.Brain and Language,93
(1),20–31.Stevens,C.,Keller,P.,&Tyler,M.(2004).Language Tonality and its effects on the perception of contour in short spoken and musical items.In S.D.Lipscomb,R.Ashley,R.O.Gjerdingen,&P.Webster(Eds.),Proceedings of the 8th international conference on music perception&cognition
(pp.713–716).Evanston,IL.Szenkovits,G.,Peelle,J.E.,Norris,D.,&Davis,M.H.(2012).Individualdifferencesin premotorand motor recruitment during speech perception.Neuropsychologia,50
,1380–1392.Tillmann,B.,Koelsch,S.,Escoffier,N.,Bigand,E.,Lalitte,P.,Friederici,A.D.,&von Cramon,D.Y.(2006).Cognitive priming in sung and instrumental music:Activation of inferior frontal cortex.Neuroimage,31
(4),1771–1782.van Essen,D.C.,Drury,H.A.,Dickson,J.,Harwell,J.,Hanlon,D.,&Anderson,C.H.(2001).An integrated software suite for surface-based analyses of cerebral cortex.Journal of the American Medical Informatics Association,8
(5),443–459.Vigneau,M.,Beaucousin,V.,Hervé,P.Y.,Duffau,H.,Crivello,F., Houdé, O.,... Tzourio-Mazoyer, N. (2006).Meta-analyzing left hemisphere language areas:Phonology,semantics,and sentence processing.Neuroimage,30
(4),1414–1432.Vigneau,M.,Beaucousin,V.,Hervé,P.Y.,Jobard,G.,Petit,L.,Crivello,F.,...Tzourio-Mazoyer,N.(2011).Whatis right-hemisphere contribution to phonological,lexico-semantic,and sentence processing?Insights from a meta-analysis.Neuroimage,54
(1),577–593.Wehrum,S.,Degé,F.,Ott,U.,Walter,B.,Stippekohl,B.,Kagerer,S.,...Stark,R.(2011).Can you hear a difference?Neuronal correlates of melodic deviance processing in children.Brain Research,1402
,80–92.Whitney,C.,Kirk,M.,O'Sullivan,J.,Ralph,M.A.L.,&Jefferies,E.(2011).The neural organization of semantic control:TMS evidence for a distributed network in left inferiorfrontaland posteriormiddle temporalgyrus.Cerebral Cortex,21
(5),1066–1075.Wong,D.,Dzemidzic,M.,Talavage,T.M.,Romito,L.M.,&Byrd,K.E.(2011).Motor control of jaw movements:An fMRI study of parafunctional clench and grind behavior.Brain Research,1383
,206–217.Yang,Y.H.,Lin,Y.C.,Su,Y.F.,&Chen,H.H.(2008).A regression approach to music emotion recognition.IEEE Transactions on Audio,Speech,and Language Processing,16
(2),448–457.猜你喜欢
河北工业大学学报(2021年4期)2021-09-23
乐器(2021年6期)2021-09-10
乐器(2021年1期)2021-09-10
中国电气工程学报(2020年17期)2020-11-30
音乐生活(2020年6期)2020-06-21
电脑知识与技术(2017年30期)2017-11-17
琴童(2017年3期)2017-04-05
中国新通信(2016年12期)2016-08-09
现代电子技术(2009年14期)2009-09-05
电气世界(2009年7期)2009-08-28
