
鸡枞菌ITS区克隆、测序及其系统发育关系
2014-01-21 02:32郭莉娟何雪梅刘小艳邹立扣
食品科学 2014年17期
龙 梅,郭 放,郭莉娟,何雪梅,刘小艳,李 蓓,罗 燕,邹立扣,*
鸡枞菌ITS区克隆、测序及其系统发育关系
龙 梅1,2,郭 放1,郭莉娟1,2,何雪梅1,2,刘小艳1,李 蓓1,罗 燕1,邹立扣1,2,*
(1.四川农业大学资源与环境学院,四川 成都 611130;2.四川农业大学都江堰校区微生物学实验室,森林资源保护与利用 实验室,四川 都江堰 611830)
采集鸡枞菌(Termitomyces)子实体,对其进行rDNA-ITS 区序列聚合酶链式反应扩增测序,利用MEGA5对rDNA-ITS不同区域作序列分析,并构建鸡枞菌转录间隔区(internal transcribed spacer,ITS)系统发育树。结果表明:在10种鸡枞菌中,测序结果表明鸡枞菌rDNA-ITS区长度在527~661 bp。系统发育树表明,10种鸡枞菌中,有8种鸡枞菌各为一支,尖盾鸡枞菌与球盖白蚁伞聚为一支;待定种A1、E1、H1独为一支,可能为新种;B1、G1、D1可能为谷堆鸡枞菌;I1可能为根白蚁伞;ZZ1可能为粗柄鸡枞菌;待定种C1不能明确。结果证明,ITS1及ITS1-5.8S rDNA-ITS2可用于鸡枞菌进行种间系统发育树的建立,但ITS1区构建的鸡枞菌种间系统发育树支持率最高,ITS2区可辅助进行某些待定种的鉴定。
鸡枞菌;系统发育;ITS序列
鸡枞菌(Termitomyces spp.)又称伞把菇、蚁巢伞、鸡丝菇等,属伞菌目,口蘑科,是一种营养丰富,味道鲜美的珍贵野生食用菌。鸡枞菌蛋白质含量高,含有多种微量元素,且具有抗氧化、镇痛抗炎的作用[1-3]。多年来多次在西昌、巴中、洪雅、资中、大邑等地采集大量新鲜鸡枞菌子实体,通过形态学鉴定对四川省鸡枞菌种类进行记述[4]。在形态学及分子生物学的基础上多次对鸡枞菌及其内生真菌有过报道[5-7]。鸡枞菌多糖、氨基酸含量均较高,本研究曾对10 种鸡枞菌进行氨基酸含量的分析,发现其总氨基酸含量高达28.86%,必需氨基酸含量在7.08%~10.20%[8-9]。
鸡枞菌因其与白蚁共生关系的复杂性,目前尚未能成功驯化栽培[10]。因此,从其他途径分析鸡枞菌的生长有重要意义。近年来,分子生物学的方法越来越多地应用于真菌的系统学研究及分类鉴定中。但是,对于鸡枞菌的系统学研究,国外仅少数报道,国内仅个别报道。Rouland-Lefevre等[11]首次采用分子生物学手段对鸡枞菌及与其共生的白蚁种群转录间隔区(internal transcribed spacer,ITS)结合分析,建立鸡枞菌属与其共生系统(白蚁)之间的进化关系,发现鸡枞菌种群是单源进化的,属于口蘑科(Tricholomataceae)。经对蚁巢伞的系统发育树和大白蚁亚科的生物分类树进行比较发现,在一定条件下,鸡枞菌和白蚁是协同进化的。Frøslev等[12]对Nlsu-rDNA和mtSSU-rDNA序列经Bayesian、Maximum Likelihood及最大简约性分析,讨论了鸡枞菌属与相关类群之间及鸡枞菌属种间的系统发育关系,发现鸡枞菌是一个自然单系群。付子艳等[13]经分子系统学对云南产鸡枞进行ITS区分析,云南鸡枞菌也是自然单系群,发现华鸡枞菌属真菌与鸡枞菌属真菌有较密切的亲缘关系。四川省是国内鸡枞菌的主要产地之一,对四川鸡枞菌的分类及分子发育等研究具有重要意义。本实验共收集了四川省内10多种鸡枞菌,确定种类有10 种,据报道目前全世界已知鸡枞菌有25 种[7],占世界种类的40%。本实验对鸡枞种内进行系统发育关系的研究,构建系统发育树,弄清鸡枞菌进化关系,旨在为鸡枞菌的进一步驯化栽培提供新的途径。
1 材料与方法
1.1 材料与试剂
1.1.1 鸡枞菌的采集与鉴定
鸡枞菌子实体采集时间为每年的6—9月,采集地为西昌、洪雅、资中、大邑、巴中。采集时拍摄照片,记录鸡枞菌子实体颜色,描述鸡枞菌形态学特征,包括其菌柄长度,菌柄直径,菌盖周长等。测量其根部距蚁巢的距离、蚁巢处土壤pH值、周围植被环境。样品采集后在室外接种,并用冰袋进行保鲜,送回实验室。实验室内对鸡枞菌子实体和产生的孢子进行观察,结合野外记录和参考资料[14]确定鸡枞菌种类。
1.1.2 试剂
真菌通用引物ITS4/ITS5 生工生物工程(上海)股份有限公司;DNA分子质量标准DL2000、Taq DNA聚合酶 宝生物工程(大连)有限公司;小量胶回收试剂盒天根生化科技(北京)有限公司。
1.1.3 仪器与设备
JSM-5900LV型扫描电子显微镜 日本电子株式会社;Eppendorf 5804R高速冷冻离心机、Eppendorf Bio-Photometer核酸蛋白仪 德国Eppendrof公司;凝胶成像系统、电泳仪 美国 Bio-Rad公司;Leica显微镜 德国Leica公司;水浴锅 上海佑柯仪器设备有限公司;小型涡旋振荡器 德国IKA公司。
1.2 鸡枞菌孢子形态特征
获得鸡枞菌孢子后,用扫描电子显微镜观察其孢子形态,取孢子置于盖玻片上,加4%戊二醛2滴固定5~10 min,用滤纸吸掉多余的水分,自然干燥[15]。在真空10 Pa,电流15 mA,时间80 s的条件下,用Hitachi离子溅射仪喷金提高样品导电性。用JSM-5900LV型扫描电镜,加速电压20 kV。
1.3 鸡枞菌子实体总DNA的提取
采用改良二次沉淀法[16]提取鸡枞菌子实体总DNA。将采集自蚁巢上方的新鲜子实体用灭菌蒸馏水冲洗2~3 次后,用无菌手术剪剪取菌柄,液氮研磨2~3 次,提取DNA。将自然风干的DNA用灭菌超纯水溶解,用核酸蛋白仪测定其总DNA浓度,经1%琼脂糖凝胶电泳检查其DNA质量,置于-20 ℃冰箱保存备用。
1.4 ITS区聚合酶链式反应(polymerase chain reaction,PCR)扩增、克隆、测序
取出备用总D NA,采用真菌通用引物I T S 4(5’-TCCTCCGCTTATTGATATGC-3’)和ITS5(5’- GGAAGTAAAAGTCGTAACAAG-3’)进行菌丝体DNA ITS区扩增。扩增体系为50 μL:40.5 μL无菌超纯水,5 μL 10×Taq Reaction Buffer(含Mg2+),1 μL 10 mmol/L dNTP,1 μL ITS4,1 μL ITS5,1 μL DNA模板,0.5 μL Taq DNA 聚合酶。反应参数:94 ℃预变性5 min;94 ℃变性1 min、52 ℃退火50 s、72 ℃延伸1 min,30 个循环;72 ℃延伸10 min。扩增产物经1%凝胶电泳检测后回收,连接pMD19-T载体,大肠杆菌JM109感受态细胞转化,进行蓝白斑筛选和菌落PCR鉴定后送生工生物工程(上海)股份有限公司测序,利用Clustal X 软件对测序结果进行比对分析。
1.5 ITS区序列及系统发育分析
将测序ITS区序列在GenBank中BLAST比对,选取其中相关序列,利用DNAstar软件分析ITS区序列特征,用Clustal X(ftp://ftp-igbmc.u-strasbg.fr/pub/Clustal X/)对ITS区序列不同基因片段进行比对对齐,ITS区序列结构如图1所示。以MEGA5(http://www.gegasoftware.net/)分析序列的碱基组成及计算遗传距离等,构建Neighbor-Joining系统发育树,Kimura双参数法计算核算距离,Bootstrap=1 000。
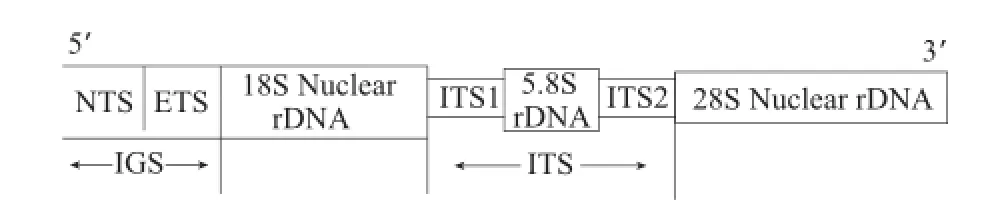
图1 ITS区示意图Fig.1 Schematic representation of ITS region
2 结果与分析
2.1 鸡枞菌孢子特征
光学显微镜及电镜镜检发现,各种鸡枞菌孢子形态、大小无差异。在光学显微镜下,观察到鸡枞菌孢子呈椭圆形、外缘光滑,萌发时在椭圆形的一头形成小尖(图2a)。在扫描电子显微镜下,观察到鸡枞菌孢子呈不规则椭球形,测定短直径在3~4 μm,长直径5~7 μm,表面不光滑,有颗粒状物质附着(图2b)。

图2 鸡枞菌孢子形态Fig.2 Morphology of Termitomyces spores
2.2 鸡枞菌种类

图3 部分鸡枞菌子实体形态Fig.3 Photographs of fruit bodies of some Termitomyces spp.
根据形态学特征[4]、孢子镜检及ITS区序列结果鉴定鸡枞菌种类主要有以下几种:普通鸡枞菌(T. albuminosus)、粗柄鸡枞菌(T. rodustus)、小鸡枞菌(T. microcarpus)、谷堆鸡枞菌(T. heimii)、端圆白蚁伞(T. tyleranus)、根白蚁伞(T. eurrhizus)、尖盾白蚁伞(T. clypentus)、裂纹白蚁伞(T. schimperi)、球盖白蚁伞(T. globules)、乌黑白蚁伞(T. badius)及9 个待定种鸡枞菌。其中测得鸡枞菌菌柄长度在5~52 cm,菌柄直径在0.5~15 cm,菌盖直径在2.1~38 cm,测得蚁巢pH值在5.9~6.5。部分鸡枞菌种类如图3所示。
2.3 测序结果及序列分析
使用引物ITS4/ITS5对鸡枞菌总DNA进行扩增,在600~700 bp处得到明亮清晰的条带,送生工生物工程(上海)股份有限公司测序后分析其序列特征。GenBank (http://www.ncbi.nlm.nih.gov/)中下载鸡枞菌ITS区序列,与本研究种类比较,使用ITS区序列构建系统发育树的菌株来源如表1所示。ITS不同区域长度、GC含量等详细情况如表2所示。结果表明不同种类鸡枞菌ITS区序列长度不等,其中ITS区最长的鸡枞菌(小鸡枞)与ITS区最短的鸡枞菌(待定种E1)相差134 bp,其余鸡枞菌ITS区序列在527~661 bp变化。
ITS1区差异较大,长度在155~248 bp,其中小鸡枞菌菌(T. microcarpus)和端圆白蚁伞(T. tyleranus)最长,为248 bp。小鸡枞GC含量最高,达到56.85%,其他鸡枞菌GC含量均在40.65%~56.85%变动。
ITS2区各种间差异较小,长度在202~251 bp之间,其中,小鸡枞(T. microcarpus)和待定鸡枞菌(A1)最长,为251 bp。待定鸡枞菌(G1)GC含量最高,达到52.00%,其他鸡枞菌在43.07%~52.00%变动。
5.8 S区相对保守,各种间长度在156~162 bp。但GC含量有较大差异,在42.00%~55.35%变动,其中待定种(A1)达到最高,为55.35%。ITS区各区GC含量的变化,可能与各区的碱基长度有关。
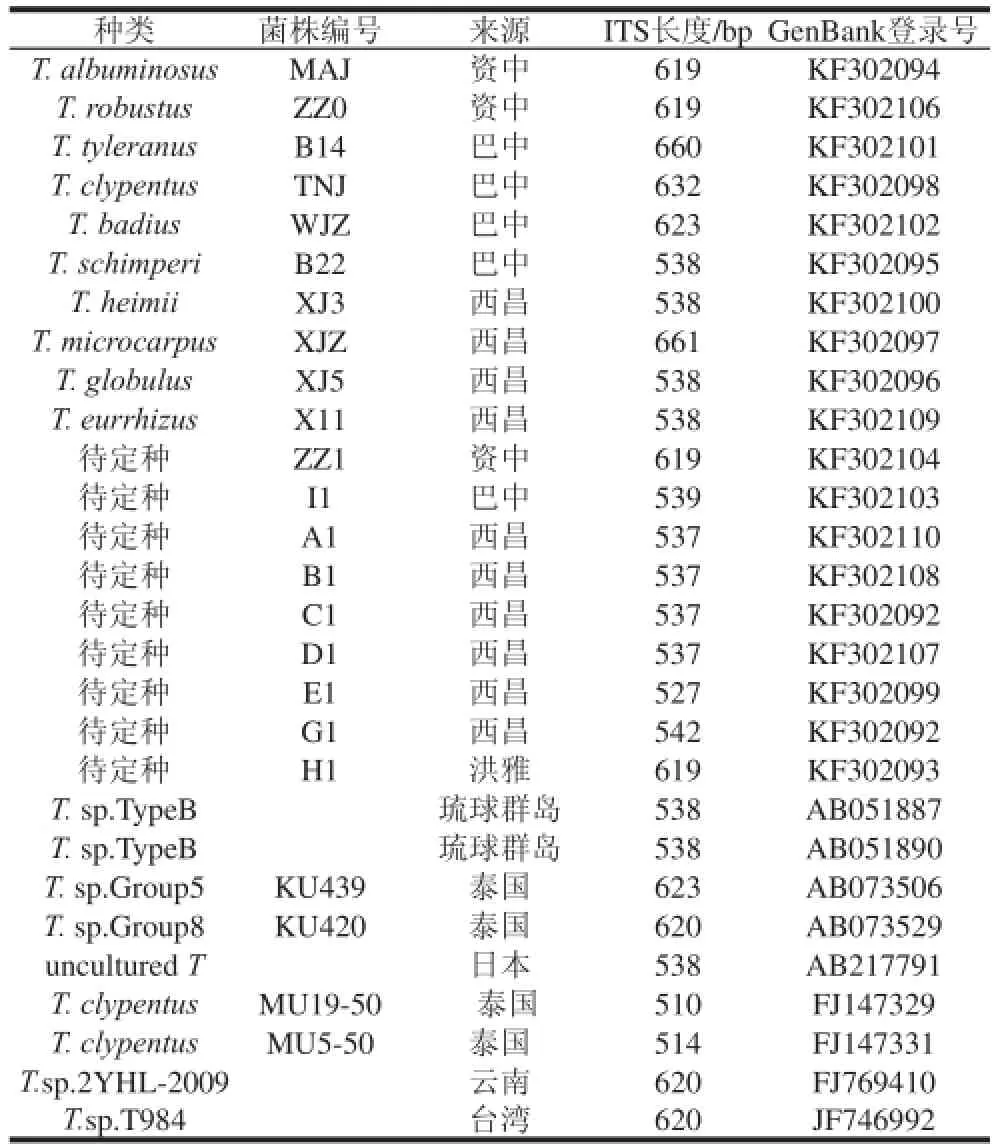
表1 用ITS区序列构建系统发育树的菌株来源Table 1 Origins of the ITS sequences used for the establishment of phylogenetic tree
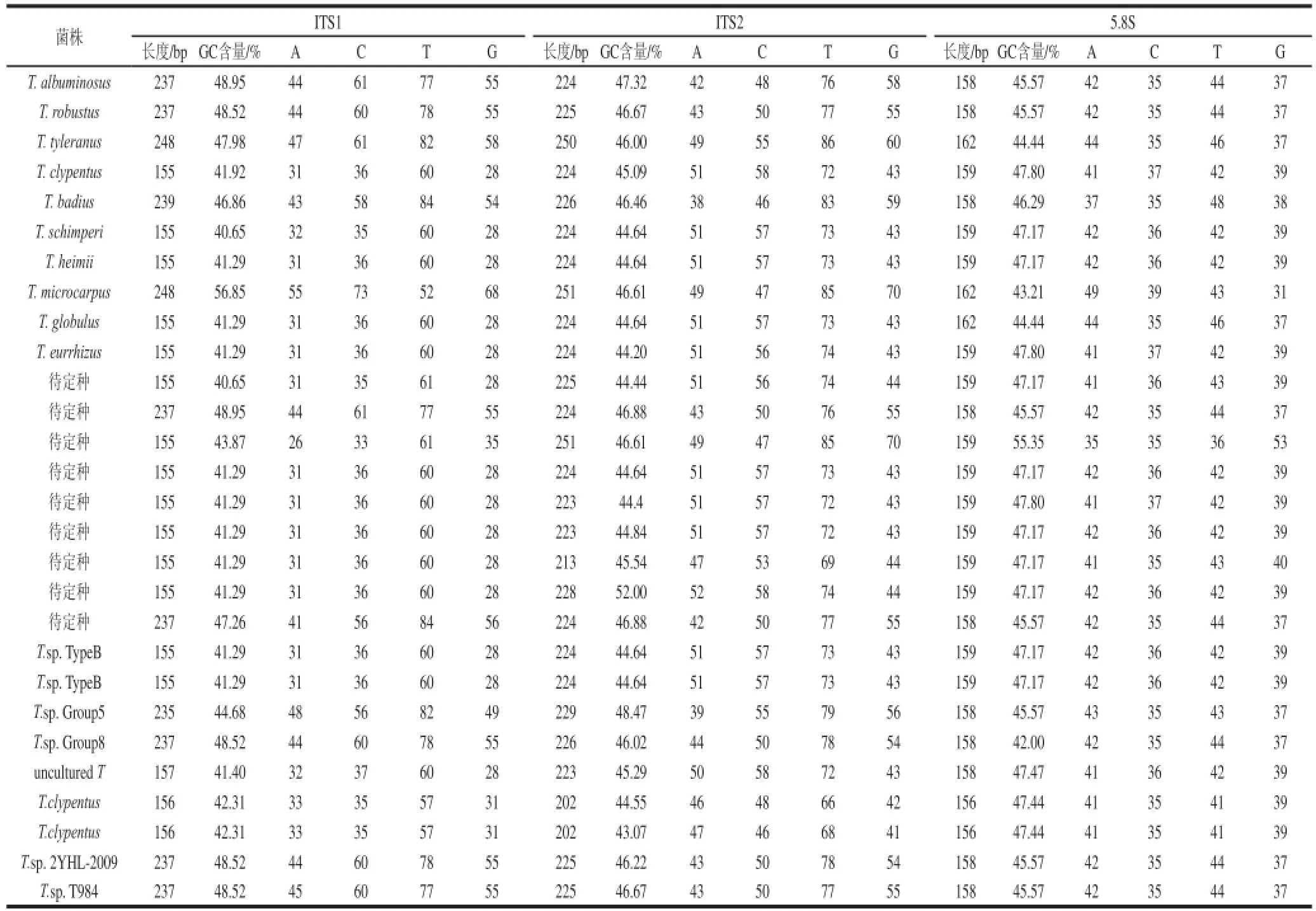
表2 ITS区碱基组成及GC含量分析Table 2 Nucleotide composition and GC contents of rDNA ITS1 and ITS2 sequences of Termitomyces
生物信息学软件分析表明,鸡枞菌种类多样性及差异主要体现在ITS1区和ITS2区,由图4可知,不同鸡枞菌在ITS1及ITS2区不同部位出现序列缺失情况不一,因此,可重新设计不同引物进行鸡枞菌种间的PCR鉴定。

图4 部分鸡枞菌rDNA-ITS缺失序列Fig.4 Partial sequences of the missing region of rDNA-ITS in Termitomyces
2.4 鸡枞菌系统发育分析
基于ITS1-5.8S rDNA-ITS2区全序列构建系统发育树表明,鸡枞菌形成两个进化组(图5a),小鸡枞菌和端圆白蚁伞聚为一组,Bootstrap(即支持率)值为64%;其余鸡枞菌及待定鸡枞菌聚为一大支,Bootstrap值为73%。在这一大支中,待定种H1、普通鸡枞菌、粗柄鸡枞菌、待定种ZZ1聚为一小支,其Bootstrap值均大于67%,最高为87%。剩下的鸡枞菌聚为另一支,最高Bootstrap值为99%,最低为40%。
基于ITS1区序列构建系统发育树表明,鸡枞菌形成两个进化组(图5b),小鸡枞菌和端圆白蚁伞聚为一支,Bootstrap值为99%。其余鸡枞分化为3 支,其中标准株AB073506独为一小支;乌黑白蚁伞、待定种H1、普通鸡枞菌、待定种I1、粗柄鸡枞菌聚为一小支,最高Bootstrap值为93%;剩余鸡枞菌聚为第三小支,最高Bootstrap值为10v0%,最低为 36%。
基于ITS2区序列构建系统发育树表明,由小鸡枞菌最先开始进化(图5c),后分为两级进化,乌黑白蚁伞独为一级;余下鸡枞菌再分为两大级进行进化,其中普通鸡枞菌、待定种I1、H1、粗柄鸡枞菌为第一级进化,剩余鸡枞菌逐级进化,除个别Bootstrap值较高外,普遍Bootstrap值较低。
综上所述,在种间关系中,小鸡枞菌与端圆白蚁伞亲缘关系较近,且由小鸡枞菌开始进化;乌黑蚁巢伞、普通鸡枞菌、粗柄鸡枞菌及待定种H1、ZZ1、I1 之间亲缘关系较近;待定种A1、B1、C1、D1、G1、E1及谷堆鸡枞菌、裂纹白蚁伞、尖盾鸡枞菌、球盖白蚁伞亲缘关系较近。在各区系统发育树中,ITS1及ITS1-5.8S rDNA-ITS2可用于鸡枞菌进行种间系统发育树的建立,但ITS1区系统发育树支持率最高,与全序列构建的系统发育树较为一致,可用于分析鸡枞菌种间差异;ITS2区支持率一般,可辅助进行某些待定种的鉴定。


图5 基于rDNA-ITS 不同区域的系统发育树Fig.5 Phylogenetic tree based on the sequences of rDNA-ITS region
3 结论与讨论
鸡枞菌为一种珍贵的野生实用菌,鸡枞菌营养价值高,主要分布在我国四川、云南、贵州等省的山区,每年6—9月采摘。近年来,由于其多糖的免疫功能及降血脂、抗肿瘤等作用的发现,吸引了大批学者对其多糖的注意[17-20]。但鸡枞菌因其与白蚁复杂的共生关系,不能人工栽培,资源不能广泛供应,故实现鸡枞菌的人工驯化栽培,具有重要意义。本实验用真菌鉴定的常规方法进行子实体形态特征观察和显微镜鉴定,根据卯晓岚[14]的分类描述,鉴定出10 个种,有2 个样品根据形态学描述及显微镜观察并不能将其准确归类,另有7个样品在采样时因形态描述资料缺乏,均归为待定种。本实验为鸡枞菌的系统发育关系作了阐明,旨在弄清鸡枞菌进化关系,获得新的途径找到解决问题的方法。
结果表明,鸡枞菌ITS区GC含量与其碱基长度存在一定关系(表2),小鸡枞菌、粗柄鸡枞菌及普通鸡枞菌在ITS1区GC含量均较其他鸡枞高,最高为56.85%,可见GC含量分析能够提供系统发育信息[15]。基于ITS1-5.8S rDNA-ITS、ITS1、ITS2区不同区段对10 种确定种鸡枞菌及9 种待定鸡枞菌进行种间发育关系进行研究,结果表明ITS1-5.8S rDNA-ITS2全序列系统发育树支持率较高,但ITS1支持率最高,结合其GC含量及碱基长度的情况能清晰的显示鸡枞种间的差异,能够应用于建立鸡枞菌种间系统发育树,ITS2区系统发育树支持率不高,可进行辅助分析。在ITS1-5.8S rDNA-ITS2区各区建立的系统发育树均可将鸡枞菌各种进行区分,可见能利用ITS1-5.8S rDNA-ITS2区及各区进行鸡枞菌种间鉴定及其进化发育关系的研究。
系统发育树结果表明,尖盾鸡枞菌(T. clypeatus)与GenBank中下载的标准株T. clypeatus(FJ147329、FJ147331)并不聚拢一支,其ITS区差异也较大,尖盾鸡枞菌ITS区长度为538 bp, FJ147329、FJ147331两标准株ITS区分别为510、514 bp。可能是不同国家在对鸡枞菌进行种间判定的标准不同,导致了不同的结果,使其在分子系统发育树中不能聚拢。而尖盾鸡枞菌在系统发育树中总是与球盖白蚁伞聚为一支,说明在分子水平上的鉴定并不能将这两种鸡枞进行区分,有待找出新的途径对两种鸡枞菌进行鉴定区分。9 个待定种中,A1、E1、H1 各为一支,可能为新种;B1、G1、D1为可能为谷堆鸡枞菌;C1与尖盾鸡枞菌及球盖白蚁伞聚为一支,暂不能明确;I1 为根白蚁伞;ZZ1为粗柄鸡枞菌的可能性较大。待定种的确定,表明由ITS1-5.8S rDNA-ITS2 区及各区进行鸡枞菌种间鉴定及其系统发育关系的研究是可行的。
根据前期研究,比较鸡枞菌的形态特征及生长环境[4],鸡枞菌的聚类可能与生长环境、生长方式、生长地域及共生白蚁种类有关。谷堆鸡枞菌、球盖白蚁伞、根白蚁伞、裂纹鸡枞菌、尖盾鸡枞菌及待定鸡枞菌聚为一大支,其采集地主要为西昌及巴中,均喜生长在混交林地或浅草地中,在形态特征上,如菌盖直径、菌柄长度等的差异较小。端圆白蚁伞与小白蚁伞聚为一大支,其采集地分别为巴中、西昌,喜夏季生长在落叶林地中,且均为群生、数量较多。粗柄鸡枞菌、尖盾鸡枞菌及普通鸡枞菌聚为一大支,采集地为资中、西昌,其生长环境差异较大,其中粗柄鸡枞菌喜生长在林缘草地或空旷处,尖盾鸡枞菌喜秋季生长在阔叶林下,普通鸡枞菌喜混交林地草坡,其聚类可能与共生白蚁种类有关[11]。
本实验利用10 种鸡枞菌及9 种待定鸡枞菌,构建四川省鸡枞菌系统发育树,因其种类占世界已知种类的40%,故深具代表性,有利于弄清四川省鸡枞菌进化关系,了解鸡枞菌生长发育过程。在四川各地采集鸡枞菌子实体,提取其总DNA进行ITS区的PCR扩增,结果发现不同种鸡枞菌ITS区长度不等,在527~661 bp,不同种长度不一,在分子水平上揭露了鸡枞菌的遗传多样性,为鸡枞菌种类序列多样性增添了新内容,提供了多种鸡枞菌ITS区序列信息,有利于鸡枞菌进行分子水平上的种间鉴定,构建了基于rDNA-ITS1不同区域的系统发育树,为四川鸡枞菌系统发育研究提供理论基础。
[1] 周继平,许鸿瑜.鸡枞菌粉不同组分的体外抗氧化活性研究[J].中国野生植物资源,2008, 27(5): 46-49.
[2] 陆奕宇, 敖宗华, 成成,等.鸡枞菌粉提取物镇痛抗炎作用的研究[J].中成药, 2007, 29(12): 1742-1745.
[3] 王一心, 狄勇, 杨桂芝. 鸡枞菌在大鼠高胆固醇血症中的抗氧化作用[J].中国预防医学杂志,2005, 6(1): 10-12.
[4] 岳爱玲,张悦,姚琼, 等.四川省鸡枞菌种类记述[J].中国食用菌,2010, 29(4): 5-7.
[5] 邹立扣, 潘欣, 韩军涛, 等. 鸡枞菌内生拟盘多毛孢菌的分离和培养及鉴定[J]. 中国食用菌, 2008, 27(5): 44-47.
[6] 邹立扣, 潘欣, 韩军涛, 等. 鸡枞菌形态学及其分子鉴定[J]. 食用菌, 2009(2): 17-18.
[7] 邹立扣, 潘欣. 粗柄鸡枞菌总DNA提起及ITS区克隆测序研究[J]. 北方园艺, 2009(6): 217-219.
[8] 邹立扣, 潘欣, 岳爱玲, 等. 四川省鸡枞菌氨基酸组成及硒元素含量分析[J]. 食品科学, 2011, 32(14): 245-248.
[9] 王化远, 何俊, 超呈裕, 等. 鸡枞菌菌丝体中多糖含量提取及测定[J].华西医大学报, 1996, 27(4): 436-437.
[10] 施渺筱, 包兴起, 李祝. 白蚁与鸡纵菌关系的研究进展[J]. 农技服务, 2011, 28(8): 1211-1212.
[11] ROULAND-LEFEVRE C, DIOUF M N, BRAUMAN A, et al. Phylogenetic relationships in Termitomyces (Family Agaricaceae) based on the nucleotide sequence of ITS: a first approach to elucidate the evolutionary history of the symbiosis between fungus-growing termites and their fungi[J]. Molecular Phylogenetics and Evolution, 2002, 22(3): 423-429.
[12] FRØSLEV T G, AANEN D K, LAESSØE T, et al. Phylogenetic relationships of Termitomyces and related taxa[J]. Mycological Research, 2003, 107: 1277-1286.
[13] 付子艳, 李荣春. 云南鸡枞菌属系统关系的初步研究[J]. 福建农林大学学报,2009, 38(3): 271-274.
[14] 卯晓岚. 中国大型真菌[M]. 郑州: 河南科学技术出版社, 2000.
[15] 孔祥林, 骆荣, 刘鲜林, 等. 贵阳腐霉(Pythium guiyanense Su)菌丝体及无性繁殖阶段的扫描电镜形态学观察[J]. 电子显微学报, 2008, 27(4): 332-335.
[16] 潘欣, 邹立扣, 彭培好, 等. 杜鹃褐斑病病原菌的分离与鉴定[J]. 北方园艺, 2008(6): 198-200.
[17] MONDAL S, CHAKRABORTY I, PRAMANIK M, et al. Structural studies of water-soluble polysaccharides of an edible mushroom, Termitomyce eurhizus: a reinvestigation[J]. Carbohydrate Research, 2004, 339: 1135-1140.
[18] MONDAL S, CHANDRA K, MAITI D, et al. Chemical analysis of a new fucoglucan isolated from an edible mushroom Termitomyces robustus[J]. Carbohydrate Research, 2008, 343: 1062-1070.
[19] MONDAL S, CHAKRABORTY I, ROUT D, et al. Isolation and structural elucidation of a water-soluble polysaccharide (PS-I) of a wild edible mushroom Termitomyces striatus[J]. Carbohydrate Research, 2006, 341(7): 878-886.
[20] CHAKRABORTY I, MONDAL S, ROUT D, et al. A water-insoluble (1→3)-β-D-glucan from the alkaline extract of an edible mushroom Termitomyces eurhizus[J]. Carbohydrate Research, 2006, 341(18): 2990-2993.
Cloning, Sequencing and Phylogenetic Relationships of ITS Region of rDNA from Termitomyces
LONG Mei1,2, GUO Fang1, GUO Li-juan1,2, HE Xue-mei1,2, LIU Xiao-yan1, LI Bei1, LUO Yan1, ZOU Li-kou1,2,*
(1. College of Resources and Environment, Sichuan Agricultural University, Chengdu 611130, China; 2. Laboratory of Microbiology, Laboratory of Forestry Resource Conservatio n and Utilization, Dujiangyan Campus of Sichuan Agricultural University, Dujiangyan 611830, China)
The fruit bodies of Termitomyces were collected, and total DNA was extracted as the tem plate to amplify internal transcribed spacer (ITS). Subsequently, the PCR products were sequenced, and the phylogenetic tree of Termitomyces was established based on different parts of ITS region by the MEGA5 software. The results showed that the rDNA-ITS region of 10 different species of Termitomyces was between 527 bp and 661 bp in length. The phylogenetic tree showed that there were eight branches of Termitomyces. T. clypeatus and T. globules were gathered together. The unknown species A1, E1 and H1 which were in a single branch may be new species. B1, G1, and D1 could be identified as T. heimii. In addition, I1 could be T. eurrhizus, and ZZ1 could be T. robustus. However, C1 could not be determined based on the present study. The results showed that the region of ITS1 and ITS1-5.8S rDNA-ITS2 could be used to build the phylogenetic tree for interspecific Termitomyces. The phylogenetic tree of ITS1 had a significantly higher supporting rate, and the ITS2 could be helpful for the confirmation of some unknown species. Therefore, this work has laid a foundation for the classification system of Termitomyces and provided ITS region sequence information for the identification at the molecular level.
Termitomyces; phylogenetic tree; ITS region
TS255.1
A
1002-6630(2014)17-0186-06
10.7506/spkx1002-6630-201417036
2013-04-19
龙梅(1991—),女,硕士研究生,研究方向为微生物分子生物学。E-mail:longzm1110@163.com
*通信作者:邹立扣(1979—),男,教授,博士,研究方向为微生物分子生物学、细菌耐药性。E-mail:zoulk124@163.com
猜你喜欢
今日农业(2021年4期)2021-11-27
食品安全导刊(2021年20期)2021-08-30
传染病信息(2021年6期)2021-02-12
作文周刊·小学三年级版(2019年16期)2019-06-20
科海故事博览·下旬刊(2019年6期)2019-04-16
中学生数理化·高一版(2018年6期)2018-07-09
中学生数理化·高一版(2017年10期)2017-12-19
故事作文·高年级(2017年11期)2017-11-15
科学启蒙(2015年8期)2015-08-07
生物医学工程学进展(2015年1期)2015-02-28
