
转GsSAMS基因苜蓿农艺性状调查
2014-01-14 02:37朱延明文益东王臻昱魏正巍
东北农业大学学报 2014年3期
朱延明,文益东,王臻昱,才 华,魏正巍
(1.东北农业大学农业生物功能基因重点实验室,哈尔滨 150030;2.塔里木大学生命科学学院,新疆 阿拉尔 843300)
转GsSAMS基因苜蓿农艺性状调查
朱延明1,文益东2,王臻昱1,才 华1,魏正巍1
(1.东北农业大学农业生物功能基因重点实验室,哈尔滨 150030;2.塔里木大学生命科学学院,新疆 阿拉尔 843300)
以前期试验获得的两个耐盐转GsSAMS基因苜蓿株系A-21和A-23为试验材料,进行产量性状和形态性状的调查与分析,为其产业化提供依据。结果表明,在产量性状上,两个转基因苜蓿株系春秋季的茎分枝数、产草量及干重与对照株系相比,不存在显著差异;在形态性状上,两个转基因苜蓿株系的生长速度、株高、节数及节间长度与对照株系相比,差异亦不显著。说明外源基因GsSAMS在苜蓿体内表达并未显著改变苜蓿的产量性状和形态性状。
紫花苜蓿;GsSAMS基因;耐盐碱;农艺性状
紫花苜蓿(Medicago sativa L.)是世界上栽培最早、分布最广泛的一种优质豆科牧草,因其蛋白含量丰富,并含有多种维生素[1],被誉为“牧草之王”。苜蓿还具有高产、抗旱、耐寒等特点,可以改良土壤结构[2],保持水土和保护环境。但苜蓿耐盐能力有限,盐碱等逆境严重影响苜蓿产量与质量。因此,如何培育耐盐碱苜蓿新品种,充分利用我国盐碱地资源[3],已成为亟待解决的重大问题。
采用传统杂交育种技术手段培育耐盐碱苜蓿新品种,存在周期长、效率低、工作量大、基因资源匮乏等弊端。近年来,随着分子生物学与转基因技术的日趋成熟,通过转基因手段获得新品种已成为一种有效途径[4]。通过转基因分子育种技术手段培育耐盐碱苜蓿新品种,能够在不改变综合优良性状基础上,来改变单一不良性状,周期短、效率高。
本研究前期将来源于野生大豆的S-腺苷甲硫氨酸合成酶基因GsSAMS转入肇东苜蓿,获得两个耐盐性明显提高的转基因苜蓿株系A-21和A-23[5],对其进行系统的产量性状及形态性状调查与分析,对实现转基因苜蓿的产业化具有重要意义。
1 材料与方法
1.1 材料
转GsSAMS基因苜蓿株系A-21、A-23和对照品种非转基因苜蓿(肇东苜蓿,由东北农业大学崔国文教授提供)。
1.2 方法
试验地点为黑龙江省哈尔滨市东北农业大学香坊实验实习基地。每个株系种植面积为30 m2,栽种时施足基肥,田间栽培及管理方法符合《农业转基因生物安全评价管理办法》附件I[6]要求。
1.2.1 产量性状调查
茎分枝数:分别调查春季与秋季单株的茎分枝个数,每个株系随机选择10株进行测定,计算平均值。
产草量:在初花期测定每平方米地上部分鲜草重量,3次重复,计算平均值(kg·m-2)。
干草量:株系混合后,取100 g鲜草,110℃杀青15 min后,80℃烘干至恒重,计算干草含量,用百分数表示。每个株系重复3次,计算平均值(%)。
1.2.2 形态性状调查
生长速度:分别在春季和秋季调查苜蓿的生长速度,每个株系随机选择10株,调查其春季和秋季20 d内的生长量,计算日生长量,取平均值(cm·d-1)。
株高:测定盛花期的株高,每个株系随机取10株进行调查,计算平均值(cm)。
节数:调查每个株系在盛花期的节数,每株取10个分枝测定节数,每个株系随机调查10株,计算平均值。
节间长度:调查每个株系在盛花期的节间长度,每个单株随机测10个节间长度,取平均值作为该株节间长度,每个株系随机取10株调查,计算平均值(cm)。
1.2.3 刈割后形态性状调查
在第一次盛花期时刈割,刈割后生长速度、株高、节数及节间长度按1.2.2调查。
2 结果与分析
2.1 转基因苜蓿产量性状的调查与分析
对转GsSAMS基因苜蓿及对照品种肇东苜蓿产量性状的调查结果如表1所示。
株系A-21和A-23与对照品种相比,在春季,两个转基因株系茎分枝数分别是46和45.3,非转基因株系为44.4,转基因株系与对照株系间不存在显著差异。在秋季,转基因株系和对照株系茎分枝数分别为71.8、72.4和69.4,无显著差异。
苜蓿作为饲草,产草量和干草含量是衡量其价值的重要指标。本研究中,转基因株系和非转基因株系的产草量分别为23.8、22.5和21.8 kg·m-2,产草量之间不存在显著差异。干草含量分别为28.4%、30.3%和29.3%,无显著差异。

表1 转GsSAMS基因苜蓿产量性状的调查结果与分析Table 1 Investigation results and analysis on yield traits of the transgenic alfalfa with GsSAMS gene
以上结果表明,两个耐盐转基因苜蓿株系与对照株系的产量性状无显著差异。
2.2 转基因苜蓿形态性状的调查与分析
2.2.1 未刈割转基因苜蓿形态性状的调查与分析
对未刈割的转GsSAMS基因苜蓿及对照品种的形态性状调查结果如表2所示。
分别对未刈割的转GsSAMS基因苜蓿的两个株系及对照品种的春季及秋季两个季节的生长速度进行测定,转基因株系与对照品种在春季生长速度分别为1.23、1.37和1.31 cm·d-1,而秋季生长速度分别为0.44、0.45和0.42 cm·d-1,春季和秋季的生长速度均无显著差异。
并对未刈割的两个转基因株系及对照品种的株高、节数及节间长度进行调查,其株高分别为141、140和148 cm,不存在显著差异,且两个转基因株系与对照品种间节数和节间长度均差异不显著。
以上结果说明未刈割时,转GsSAMS基因苜蓿在形态性状上与对照品种相比无显著差异。

表2 转GsSAMs基因苜蓿形态性状的调查结果与分析Table 2 Investigation results and analysis on morphological traits of transgenic alfalfa with GsSAMS gene
2.2.2 刈割后转基因苜蓿形态性状的调查与分析
对刈割后的转GsSAMS基因苜蓿及对照品种的形态性状调查结果如表3所示。
分别对刈割后的转GsSAMS基因苜蓿的两个株系及对照品种的生长速度进行测定,转基因株系与对照品种在第一次盛花期刈割后,其再生生长速度分别为3.09、3.02和2.98 cm·d-1,刈割后的生长速度无显著差异。
并对刈割后两个转基因株系及对照品种的株高、节数和节间长度进行调查,结果表明刈割后的盛花期两个转基因株系与对照品种之间节数、节间长度和株高差均无显著差异。
以上结果说明转GsSAMS基因苜蓿刈割后与对照品种的形态性状相比无显著差异。
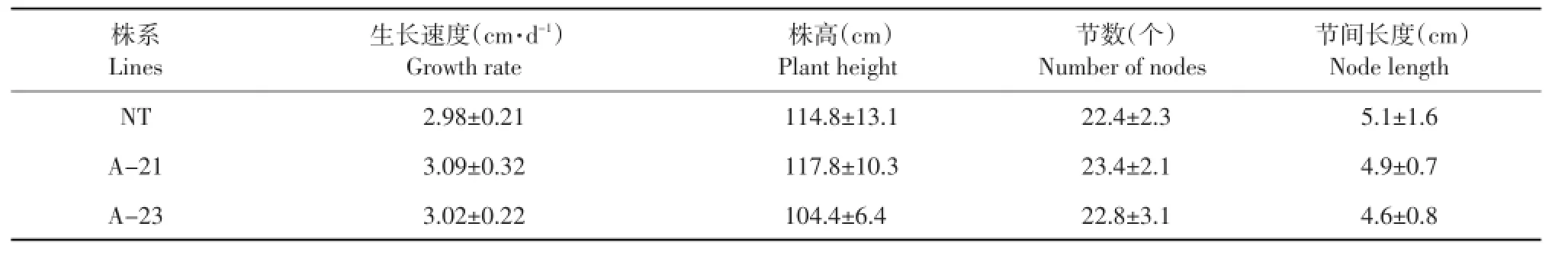
表3 刈割后转GsSAMS基因苜蓿形态性状的调查结果与分析Table 3 Investigation results and analysis on morphological traits of cut transgenic alfalfa with GsSAMS gene
3 讨论与结论
植物转基因主要目的不仅是将目的基因导入受体植物使其获得相应的优良性状,而且还不应改变其原有的优良性状。因此,转基因株系农艺性状表现的好坏,直接影响通过转基因技术获得的植物能否实现产业化。国内外有许多有关转基因植株出现变异或生长受抑制的报道[7-9]。引起这些变异的原因主要有以下几方面:一是外源基因插入植物基因组的位点是随机的,这可能导致植物外显基因被打断,从而无法表达,影响植物正常生长;而且外源基因也可能因为其插入的位置不同,表现型也不同(即转基因的位置效应)[10];二是外源基因插入的拷贝数不同,也可能会引起转基因植株表现型的变异;三是组织培养过程中,体细胞无性系的变异,转座子的活化,DNA发生点突变等,都可能引起转基因植株及其后代的表现型出现变异。因此必须要对转基因植株的农艺性状进行系统研究,只有保持农艺性状的稳定,才能使转基因植物具有良好的应用价值,并实现产业化。
本研究中,两个耐盐转基因株系的农艺性状与对照植株相比无显著差异,说明将GsSAMS导入苜蓿,使转基因苜蓿获得耐盐性,又能保持原来优良的农艺性状,具有良好的应用价值。但是要实现产业化,还需对转基因植物的安全性问题,如食品安全性以及环境安全性[11],进行深入评价分析。
[1]王鑫,马永祥,李娟.紫花苜蓿营养成分及主要生物学特性[J].草业科学,2003,20(10):39-40.
[2]张晓琴,胡明贵.紫花苜蓿对盐渍化土地理化性质的影响[J].草业科学,2004,21(11):31-34.
[3]化烨,才华,柏锡,等.植物耐盐基因工程研究进展[J].东北农业大学学报,2010,41(10):150-156.
[4]Trinh T H,Ratet P,Kondorosi E,et al.Rapid and efficient transformation of diploid Medicago truncatula and Medcago sativa ssp.falcata lines improved in somatic embryogenesis[J].Plant Cell Reports,1998,17(5):345-355.
[5]Hua Y,Zhang B X,Cai H,et al.Stress-inducible expression of GsSAMS2 enhances salt tolerance in transgenic Medicago sativa [J].African Journal of Biotechnology,2012,11(17):4030-4038.
[6]农业部第五次常务会议.农业转基因生物安全评价管理办法,中华人民共和国农业部令第8号[S].北京:农业部,2002.
[7]盛慧,朱延明,李杰,等.DREB2A基因对苜蓿遗传转化的研究[J].草业科学,2007,24(3):40-45.
[8]Jin T,Chang Q,Li W,et al.Stress-inducible expression of GmDREB1 conferred salt tolerance in transgenic alfalfa[J].Plant Cell Tissue and Organ Culture,2010,100(2):219-227.
[9]Morran S,Eini O,Pyvovasenko T,et al.Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors[J].Plant Biotechnology Journal,2011,9(2): 230-249.
[10]郭玉双,朱延明,李杰,等.转双价抗真菌基因大豆主要农艺性状的调查与分析[J].东北农业大学学报,2008,31(9):10-12.
[11]李瑞峰,王莹,王宇,等.转基因作物及其生物安全性[J].东北农业大学学报,2007,38(3):405-410.
Investigation and analysis of main agronomic traits of transgenic alfalfa withGsSAMSgene
ZHU Yanming1,WEN Yidong2,WANG Zhenyu1,CAI Hua1,WEI Zhengwei1(1.Key Laboratory of Agricultural Biological Functional Genes,Northeast Agricultural University,Harbin 150030,China;2.School of Life Sciences,Tarim University,Alar Xinjiang 843300, China)
The agronomic traits including the yield traits,morphological traits and quality traits of GsSAMStransgenic alfalfa(line A-21 and A-23)were investigated and analyzed statistically.Our results showed no significant differences between the transgenic and non-transgenic alfalfa plants in the number of branches,grass yield and dry weight.We also found no significant differences regarding to the growth rate in spring and autumn,plant height,the number of nodes and the node length.Taken together,our results indicated that over-expression ofGsSAMSin alfalfa did not alter the agronomic traits of transgenic alfalfa.
alfalfa;GsSAMSgene;tolerance to salt-stress;agronomic trait
S541.9
A
1005-9369(2014)03-0031-03
时间2014-3-20 17:47:00 [URL]http://www.cnki.net/kcms/detail/23.1391.S.20140320.1747.003.html
朱延明,文益东,王臻昱,等.转GsSAMS基因苜蓿农艺性状调查[J].东北农业大学学报,2014,45(3):31-33.
Zhu Yanming,Wen Yidong,Wang Zhenyu,et al.Investigation and analysis of main agronomic traits of transgenic alfalfa withGsSAMSgene[J].Journal of Northeast Agricultural University,2014,45(3):31-33.(in Chinese with English abstract)
2012-02-29
国家自然科学基金资助项目(31171578);黑龙江省高校科技创新团队建设项目(2011TD005);国家大豆转基因重大专项(2011 ZX08004-002);东北农业大学创新团队项目(CXT004-1)
朱延明(1955-),男,教授,博士,博士生导师,研究方向为植物基因工程与分子生物学。E-mail:ymzhu2001@neau.edu.cn
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
科学文化评论(2021年3期)2021-11-22
食品研究与开发(2020年18期)2020-09-10
河北科技师范学院学报(2020年1期)2020-07-02
四川农业科技(2019年5期)2019-07-01
中学生英语·外语教学与研究(2017年10期)2017-11-16
作文周刊(高考版)(2016年16期)2017-06-01
艺术研究(2016年3期)2016-10-27
党建文汇·下(2016年5期)2016-05-14
奥秘(2016年4期)2016-04-21
