
接种菌根菌后板栗幼苗光合作用的光响应曲线
2013-12-27 04:57柴迪迪郭素娟秦天天孙小兵
中南林业科技大学学报 2013年8期
柴迪迪,郭素娟,秦天天,邹 峰,孙小兵
( 北京林业大学 省部共建教育部森林培育与保护重点开放实验室,北京 100083)
接种菌根菌后板栗幼苗光合作用的光响应曲线
柴迪迪,郭素娟,秦天天,邹 峰,孙小兵
( 北京林业大学 省部共建教育部森林培育与保护重点开放实验室,北京 100083)
以接种外生菌根菌绵毛丝膜菌Cortinarius sublanatus的板栗幼苗和未接菌的板栗幼苗为材料,分别测定它们叶片的光合数据,用非直角双曲线方法对他们的光响应曲线进行拟合。拟合结果显示,两种材料非直角双曲线模型的拟合结果与实际测量值非常接近,接种菌根菌后的苗木α表光量子效率增长了13.33%,Pmax最大净光合速率增长了49.72%,Rd暗呼吸速率降低了26.91%,LCP光补偿点降低了4.34%,LSP光饱和点增长了25.66%。上述结果表明非直角双曲线模型适合板栗幼苗光合作用的光响应曲线拟合,接种菌根菌后板栗幼苗的光能利用效率增高,利用弱光能力增强,对强光的适应能力和利用能力增强,而消耗光合产物的速率降低。
板栗;外生菌根菌;光响应曲线;非直角双曲线
光合作用是植物获得能源的基础,研究了解植物的光合作用是非常有意义的。研究的方法主要是通过光响应曲线拟合,获得相应的生理参数,这些生理参数是研究各种植物生理生态学的基础,可以反映出不同植物对环境适应能力的差异[1-2]。目前,国内外有多种模型用于光响应曲线拟合,主要分为双曲线和指数模型,双曲线模型有包含直角双曲线和非直角双曲线两种模型,非直角双曲线模型与其他模型相比较,反映植物光合作用特征更准确,可以参考的参数较多,但是拟合过程复杂[3-6]。
板栗Castanea mollissima Bl.属于壳斗科Fagaceae栗属Castanea Miller植物,在我国北方地区主要的栽培品种为中国栗和日本栗,产地分布在山东、北京、天津、河北和河南北部等地,在太行山及燕山山脉的片麻岩地区分布较为集中[7-8]。板栗的各种优良抗性都与它的外生菌根真菌密不可分,接种外生菌根真菌可以提高板栗苗木的抗旱性、加快苗木的生长速度和增强根系从土壤中吸收营养元素的能力等,针对上述内容开展的相关研究较多[9-11],而菌根化对板栗光合作用的影响研究相对较少,虽然有部分学者进行过初步的研究[10,12],但是利用非直角双曲线模型分析的研究尚未见报道。
本研究以接种菌根菌和未接菌的板栗幼苗为研究对象,采用非直角双曲线方法对其光响应曲线进行研究,讨论接种菌根菌以后对板栗幼苗的影响,为进一步研究板栗菌根菌共生机理和菌根菌的应用开发奠定基础。
1 材料与方法
1.1 菌剂制备和苗木培育
1.1.1 菌剂制备
本实验室保存菌种,绵毛丝膜菌Cortinarius sublanatus,制作菌剂后侵染板栗幼苗效果良好[11]。利用PDA 培养基平板培养,然后从平板培养基内打孔得到菌饼,接种到PD(不含琼脂)液体培养基内,摇床培养1周,之后将含有大量菌丝的菌液加入到灭过菌的的基质中,在培养箱内进行菌根菌种扩大培养,制作菌剂。菌剂基质按照草炭∶蛭石∶珍珠岩以3∶1∶1比例混和,每kg基质中加入25 g红糖。
1.1.2 苗木材料
河北省迁西县板栗主栽品种燕山早丰种子,2011年3月5日,于温室内低温层积催芽,4月18日播种,容器为黑色塑料营养杯,规格高20 cm,直径18 cm,温室土壤和沙子比例为3∶1,用1% 福尔马林浇灌消毒基质。每盆播种1个板栗种子,每5盆一个重复,每种处理4个重复,并在周围设置保护行。板栗苗在第一对真叶展开时进行接种,此时的接菌效果最好[11]。每个容器苗接种10 g,在距苗6 cm 处对衬打孔,深度为5 cm,每孔加入搓碎的菌剂5 g,覆盖基质,对照苗用同样方法接入等量菌剂基质。接种1个月后,用番红-淡绿染色法测定菌根侵染率[13]。
侵染率计算公式:
侵染率=侵染根数/全部根数×100%。
1.2 测定方法和拟合方法
1.2.1 光合测定方法
从接种菌根菌和未接菌的板栗幼苗中分别选出具有代表性的植株和叶片,使用LI-6400(美国 LICOR 公司生产)便携式光合测量系统,红蓝光源叶室测定不同处理板栗幼苗叶片的IAPR(瞬时光合速率)。密闭系统测定条件:使用“ light-curve”曲线测定功能,设定光源PPFD(光合光量子通量密度)梯度值分别为:0、20、50、100、150、300、500、800、1 200、1 400、1 600 μmol·m-2s-1,仪器自动测定板栗叶片Pn(净光合速率)。叶室内温度(30±1)℃,气体流速(400±1) mmol·s-1,CO2浓度(400±1) μmol·mol-1。
1.2.2 拟合方法
非直角双曲线模型来源于:
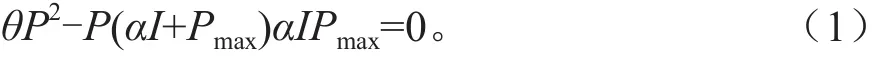
式(1)中α为表观量子效率,θ(0≤θ≤1)为光响应曲线曲角,I为光合有效辐射(μmol·m-2s-1),P为总光合速率(μmol·m-2s-1),Pmax为最大光合速率(μmol·m-2s-1)。

式 (2)中 Pn为净光合速率(μmol·m-2s-1),Rd为暗呼吸速率(μmol·m-2s-1)。当θ≠0时即为非直角双曲线,将式(2)代入式(1)变形得到:

运用SPSS17.0软件,通过迭代计算的方法对接菌和未接菌的幼苗光响应曲线拟合,式(3)中的Rd、Pmax和θ的预估值和预差值,拟合后的预差值用于标准曲线的绘图,在低光强下,0~150 μmol·m-2s-1范围内,随着光强的增大光合速率线性增大,对Pn-I线性回归直线,计算α、LSP、LCP三个值[6,14-17]。
2 结果与分析
2.1 光合仪测定数据结果
在培养苗木的温室内,用光合仪分别测定,得到接种菌根菌的板栗幼苗叶片和未接菌的叶片光合数据(见表1)。

表1 光合仪测定数据Table 1 Data measured by portable photosynthesis system
2.2 侵染率结果
接种绵毛丝膜菌Cortinarius sublanatus制作的菌剂1个月后,调查接菌后的板栗幼苗侵染率,结果为56.38%。
2.3 光响应曲线拟合结果
由图1可以看出,用非直角双曲线模型拟合未接菌板栗幼苗的光响应曲线结果接近实际测量值,拟合效果好,可以解释至少96. 2%的变异。有效光辐射值在 0 ~ 150 μmol·m-2s-1区间时,I与Pn两者呈线性关系;有效光辐射值在大于150 μmol·m-2s-1区间时,随着I的梯度升高,Pn在此阶段升高速度逐渐变缓慢,直到最后下降。

图1 非直角双曲线模型拟合未接菌板栗苗叶片的光响应曲线Fig. 1 Light response curve of un-inoculated chestnut seedling leaves ftted by non-hyperbola
由图2可以看出,用非直角双曲线模型拟合接菌板栗幼苗的光响应曲线结果接近实际测量值,拟合效果好,可以解释至少99. 2%的变异。随着I在不同区间梯度改变,Pn的改变趋势与未接菌幼苗相同,有效光辐射值在0~150 μmol·m-2s-1区间,两者呈线性关系,大于150 μmol·m-2s-1区间,随着I的梯度升高,Pn值在此阶段升高速度逐渐变缓慢,直到最后下降。但是接种菌根菌幼苗增速减慢的过程更加平缓,而且在大于150 μmol·m-2s-1区间,相同的I,接菌幼苗的Pn值明显高于未接菌幼苗。
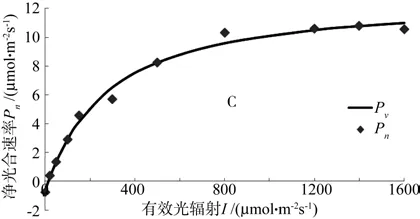
图2 非直角双曲线模型拟合接菌板栗苗叶片的光响应曲线Fig. 2 Light response curve of inoculated chestnut seeding leaves ftted by non-hyperbola
在光照强度较低的时候,光合速率随光强的增大呈线性增高 (见图3和图4)。此时可以对LCP、Rd、α进行线性回归计算。当光照强度在0~150 μmol·m-2s-1范围时,对 Pn-I 进行直线回归,得到直线方程,未接菌苗拟合直线方程为y=0.03x-0.807,拟合直线与实际测量值接近,可以解释至少96.2%的变异(见图3)。该直线与x轴的交点值为未接菌苗的 LCP= 26.900 μmol·m-2s-1,直线与y=Pmax线的交点值为未接菌苗的LSP=324.700 μmol·m-2s-1,直线的斜率为未接菌苗的α=0.03。
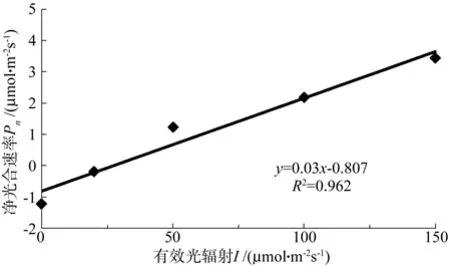
图 3 光照强度 0~150 μmol·m-2s-1 条件下未接菌苗的 Pn– I直线回归Fig. 3 Pn– I linear regression of un-inoculated chestnut seedling under conditions of illumination intensity 0~150 μmol·m-2s-1

图 4 光照强度 0~150 μmol·m-2s-1 条件下接菌的 Pn-I直线回归Fig. 4 Pn– I linear regression of inoculated chestnut seedling under conditions of illumination intensity 0~150μmol·m-2s-1
接菌苗的拟合直线方程为y=0.034x-0.497,拟合直线与实际测量值十分接近,可以解释至少99.2%的变异(见图4),该直线与x轴的交点值为接种菌根菌苗的 LCP= 14.618 μmol·m-2s-1,直线与y=Pmax线的交点值为接种菌根菌苗的LSP=408.029 μmol·m-2s-1,直线的斜率为接种菌根菌苗的α=0.034。
2.4 不同处理的光响应曲线参数比较结果
接菌板栗苗的叶片表光量子效率值为0.034,未接菌板栗苗的叶片表光量子效率值为0.03,接种菌根菌后表光量子效率增加了13.33%。接菌苗的 Pmax值为 13.376 μmol·m-2s-1,未接菌苗的 Pmax值为 8.934 μmol·m-2s-1,接种菌根菌后 Pmax增长了49.72%。接菌苗的 Rd为 0.611 μmol·m-2s-1,未接菌苗的 Rd为 0.836 μmol·m-2s-1,接种菌根菌后 Rd降低了 26.91%。接菌苗的 LCP为 14.618 μmol·m-2s-1,未接菌苗的 LCP为 26.900 μmol·m-2s-1,接种菌根菌后LCP降低了54.34%。接菌苗的LSP为408.29 μmol·m-2s-1,未接菌苗的 LSP为 324.700 μmol·m-2s-1,接种菌根菌后LSP增加了25.66%(见表2)。
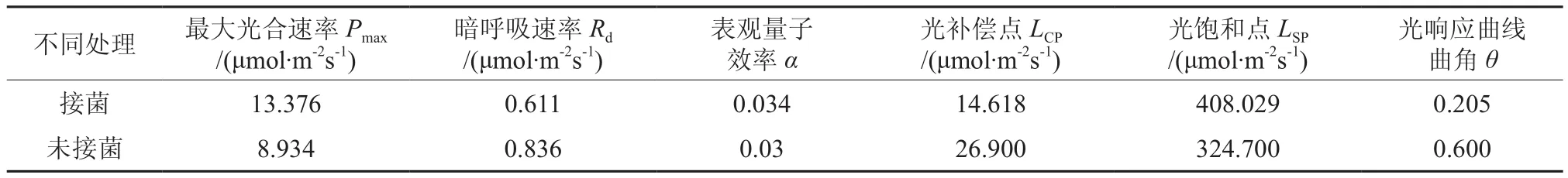
表2 不同处理的光响应曲线参数及光饱和点、光补偿点Table 2 Parameters of light response curves, VLSP and VLCP with different treatments
3 讨论与结论
植物光合作用拟合的方法有二项式拟合、直角双曲线拟合等方法,但是其中以非直角双曲线拟合得出的光合作用生理参数值最为符合生理意义[4-6]。本研究中用非直角双曲线模型拟合结果接近实际测量值,完整的反映了板栗叶片在稳定的外界环境条件下,随着I的梯度改变,Pn的改变趋势,在光照强度较低时,突然增加光强,使叶片的Pn值快速上升达到正常值,此过程称为光合作用诱导期。诱导初期,CO2浓度相对饱和,气孔限制因素是非主要因素,在这个阶段板栗叶片Pn主要受中间产物水平和酶活化水平的限制。诱导后期,主要限制因素为气孔限制,即使此时Pn也随着气孔导度的增加而增加,但是增速却有较大差异,使得光吸收与CO2同化不协调,Rubisco 羧化活性降低,因此Pn值在此阶段升高缓慢[18-20],而接种菌根菌后板栗苗木在此阶段增速减慢的过程更加平缓,并且Pn值明显高于对照,是因为菌根菌促进了板栗根系对水分和养分的吸收,间接地增加叶片内水分和叶绿素含量,从而使植物减缓对外界胁迫的感受且Pn值较高。
本实验中利用该方法拟合出的数据,与许多学者的结论相近,例如植物最大初始量子效率理论上小于0.125,且在强光下光响应曲线趋于平缓[21-22],阳生植物光补偿点在 9 ~ 18 μmol·m-2s-1[23]。因此本研究的拟合结果真实的反映了板栗幼苗的光合特点,同时通过对比可以了解,接种菌根菌后,板栗苗木光合特性的变化。
对比分析不同数据的意义,表观量子效率代表的是光能利用效率,接菌与未接菌苗的表观量子效率均在0.03~0.07 之间,属于生长良好的范围内[24]。接种菌根菌后表光量子效率增加了13.33%,表明菌根菌促进了苗木对光能的利用效率。
最大净光合速率,这一指标反应不同品种对强光的利用能力[15]。接菌后Pmax增长了49.72%,由此可以看出接种菌根菌促进了叶片对强光的利用能力。造成净光合速率增大的原因是菌根菌能促进植物生长,增加多种营养元素的吸收,导致叶肉质化程度增大,叶绿素含量增高,进而净光合速率增大。
暗呼吸速率,能够反映植物消耗光合产物的速率[15],接种菌根菌后Rd降低了26.91%,由此可见,接种菌根菌后减少了板栗叶片的消耗产物速度。由于接种菌根菌后,板栗菌根周围菌丝网络可以增大吸收面积,降低土壤与植物之间的液流阻力,减少了根系吸收土壤养分和水分所需要的能量[25],因此接种菌根菌后板栗苗的暗呼吸速度减弱。
光补偿点LCP是植物利用弱光能力的重要指标,其值越小表明植物利用光能的能力越强[26]。接种菌根菌后LCP降低了54.34%,由此可见,接种菌根菌增强了板栗幼苗对弱光能的利用能力。
LSP光饱和点能够反应不同品种对强光的适应能力[27],接种菌根菌后LSP降低了25.66%,由此可见,接种菌根菌提高了板栗幼苗对强光的适应能力。由于强光会引起蒸腾作用增强,导致植物易受干旱因素影响,吕全等人在研究板栗幼苗抗旱机理时发现,菌根菌(绵毛丝膜菌)处理后的苗木,光合作用能力增强,尤其是在逆境条件下,促进作用更加明显[10]。分析其产生的主要原因为菌根菌增大根系吸收面积,提高吸水能力,而且菌套还能防止根系内水分的丧失,增加对抗旱胁迫的抵御能力,进而增强了对强光的适应能力[28]。
通过对接菌和未接菌的板栗幼苗光合数据进行非直角双曲线拟合,可以全面的了解接种菌根菌后板栗植物光合特性的变化,经过生理生化的分析,解释产生变化的原因,为进一步研究菌根菌侵染机理和菌根化苗木在生产实际应用奠定基础。本研究只选用了其中一种效果较好的菌根真菌作为材料,但可以侵染板栗的菌根菌种类较多,在今后的研究中将选用不同种类的菌根真菌回接板栗幼苗,并进一步对光合数据拟合对比分析。
[1] Kull O, Kruijt B. Leaf photosynthetic light response:a mechanistic model for scaling photosynthesis to leaves and canopies[J]. Functional Ecology, 1998, 12(5):767-777.
[2] Niinemets U, Oja V, Kull O. Shape of leaf photosynthetic electron transport versus temperature response curve is not constant along canopy light gradients in temperate deciduous trees[J]. Plant, Cell & Environment, 1999, 22 (12): 1497-1513.
[3] Walcroft A S, Whitehead D, SilvesterW B, et al. The response of photosynthetic model parameters to temperature and nitrogen concentration in Pinus radiata D. Don[J]. Plant, Cell &Environment, 1997, 20 (11): 1338-1348.
[4] Olsson T, Leverenz J W. Non2uniform stomatal closure and the apparent convexity of the photosynthetic photon flux density response curve[J]. Plant, Cell & Environment, 1994, 17(6): 701-710.
[5] 吴家兵,关德新,张 弥,等. 长白山地区蒙古栎光合特性[J]. 中国科学院研究生院学报,2006,23(4): 548-554.
[6] 张 弥,吴家兵,关德新,等. 长白山阔叶红松林主要树种光合作用的光响应曲线[J]. 应用生态学报, 2006, 17(9): 1575-1578.
[7] 郗荣廷.果树栽培学分论[M].北京:中国农业出版社, 1997.
[8] 熊 欢,郭素娟,李广会,等.不同叶苞比对板栗光合特性及果实品质的影响[J]. 中南林业科技大学学报,2012,32(10):52-57.
[9] 冯 固, 徐 冰, 秦 岭, 等. 外生菌根真菌对板栗生长及养分吸收的影响[J]. 园艺学报, 2003, 30 (3): 311-313.
[10] 吕 全,雷增普.外生菌根提高板栗苗木抗旱性能及其机理的研究[J].林业科学研究,2000,13(3): 249-256.
[11] 张 波.板栗菌根化容器嫁接苗培养技术研究[D].北京:北京林业大学,2007.
[12] 池彦涛.缺铁及接种外生菌根真菌对板栗苗生理的影响[D].武汉:华中农业大学, 2012.
[13] 牛晓丹.板栗菌根菌剂制作技术与芽苗嫁接成活机理研究[D].北京:北京林业大学,2009.
[14] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].农业基础科学,2005,21(8):76-79.
[15] 靳甜甜,刘国华,胡婵娟,等.黄土高原常见造林树种光合蒸腾特征[J]. 生态学报, 2008, 28(11): 758-5765.
[16] 张保玉.葡萄光合作用光响应曲线的研究[D] .陕西省杨凌:西北农林科技大学, 2009.
[17] 佘诚棋,程 鹏,季琳琳,等.油茶光合作用光响应曲线的拟合[J]. 经济林研究, 2012, 30(1):118-120.
[18] 许大全,徐宝基.气孔限制在植物叶片光合诱导中的作用[J].植物生理学报,1989,15(3):275-280.
[19] Zhang Hai-Bo, Xu Da-Quan. Different mechanisms for photosystem reversible down-regulation[J]. Photosynthetica,1992, 26: 261.
[20] 杨晓玲,杨 晴,郭守华,等.燕龙板栗光合作用及其相关因素的日变化[J]. 经济林研究, 2008,26(1) : 67-70.
[21] Long SP, Humphries S, Falkowski PG. Photo inhibition of photosynthesis in nature[J]. Annu Rev Plant Physiol Mol Biol,1994, 45: 633-662.
[22] 陆佩玲,于 强,罗 毅,等.冬小麦光合作用的光响应曲线的拟合[J].中国农业气象, 2001, 22(2): 12-14.
[23] 潘瑞炽. 植物生理学[M] (第四版). 北京: 高等教育出版社,2001: 55-57, 91-95.
[24] 丘国维.植物光合作用的效率.植物生理学和分子生物学[M].北京:科学出版社, 1992. 236-243.
[25] Dixon R K. Comparative water relation of container-grow and bare-root ectomycorrhizal and nonmycorrhizal Quercus velutine seedlings [J]. Can J Bot, 1983, 61: 1559-1565.
[26] 潘瑞炽.植物生理学[M] (第五版).北京:高等教育出版社,2004. 88.
[27] 陈晓林,陈建华,侯伯鑫,等.不同品种红花檵木光合特性的比较分析[J]. 中南林业科技大学学报, 2010, 30(5):117-122.
[28] Reid C P P. Root Physiology and Symbiosis[M]. Nancy, 1978.392-408.
Light response curve of inoculated Castanea mollissima seedling photosynthesis
CHAI Di-di, GUO Su-juan, QIN Tian-tian, ZOU Feng, SUN Xiao-bing
(Key Lab. for Siliviculture and Conservation Jointly Constructed by Provinces and State Ministry of Education, Beijing Forestry University, Beijing 100083, China)
By taking Castanea mollissima seedlings inoculated with Cortinarius sublanatus and no ctomycorrhizal fungi inoculated chestnut seedlings as the tested materials, their photosynthetic data were respectively measured. Based on the photosynthetic data, the light response curves of the two tested materials were ftted by using the non-rectangular hyperbolic model. The results show that the ftting value of non-rectangular hyperbolic model was close to the measured value; the apparent quantum yield of inoculated chestnut seedling increased by 13.33%,the maximum net photosynthetic rate of inoculated chestnut seeding increased by 49.72%, the dark respiration rate of inoculated chestnut seeding decreased by 26.91%, the light compensation point of inoculated chestnut seeding decreased by 4.34%,the light saturation point of inoculated chestnut seeding increased by25.66%. The conclusion was that the rectangular hyperbolic model was propitious to fit the photosynthetic data of chestnut seedling; after the seedling inoculated with C. sublanatus, their’s effciency of light utilization was increased,the capacity of using low light was increased and the capacity of ftting and using high light was increased,however the speed of consuming of photosynthetic products was reduced.
Castanea mollissima; ectomycorrhizal fungi; light response curve; non-rectangular hyperbolic
S723.1;S664.2
A
1673-923X(2013)08-0038-05
2013-01-21
国家林业公益性行业科研专项重大项目“板栗产业链环境友好丰产关键技术研究与示范”(项目编号:201204401),十二五”科技支撑专题“北方板栗高效生产关键技术研究与示范”(项目编号:2013BAD14B0402)
柴迪迪(1985-),男,黑龙江安达人,博士研究生,主要从事苗木培育方面的研究;E-mail:040108311@163.com
郭素娟(1965-),女,内蒙古呼和浩特人,博士生导师,主要从事经济林栽培和利用理论与技术研究;
E-mail:gwangzs@263.net
[本文编校:文凤鸣]
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
特种经济动植物(2022年2期)2022-02-15
食药用菌(2021年4期)2021-01-08
新农业(2020年15期)2020-12-19
新农业(2020年8期)2020-08-26
中国林副特产(2020年2期)2020-01-15
小学生学习指导(低年级)(2019年9期)2019-09-25
中学生数理化·中考版(2019年8期)2019-07-13
中学生数理化·七年级数学人教版(2019年4期)2019-05-20
食品安全导刊(2018年18期)2018-01-18
