
中国外生菌根真菌研究进展
2013-12-23 05:12:28温祝桂陈亚华
生物技术通报 2013年2期
温祝桂 陈亚华
(南京农业大学生命科学学院,南京 210095)
菌根是土壤中真菌与植物根系形成的互惠共生体,根据其形态和解剖学特征,可分为外生菌根、内生菌根和内外生菌根三大类。外生菌根真菌在生物学、生态系统稳定、生态安全、环境修复及社会经济发展等方面所表现出的巨大研究潜力和应用价值愈发引起人们的关注。
本文是通过万方数字期刊数据库,以“外生菌根”为主题词进行了检索(2011年11月),排除一些非相关文献,共检索出344篇中文目标文献;通过Web of Science数据库,以“China”和“Ectomycorrhizal fungi”为主题词进行了检索(2012年1月),排除部分以国外第1署名单位和以国外通讯作者发表的文献,共检索出116篇英文文献。对上述460篇文献进行了归纳汇总,较为全面地阐述了国内有关外生菌根的研究现状和研究成果。
从图1可以看出,我国外生菌根菌研究相关文献以中文形式发表的数量自2007年以后呈下降趋势,但以英文形式发表的文献自2009年以后有明显地增长。近5年,我国外生菌根菌研究相关文献每年发表的数量平均在50篇左右,呈相对平稳状态。
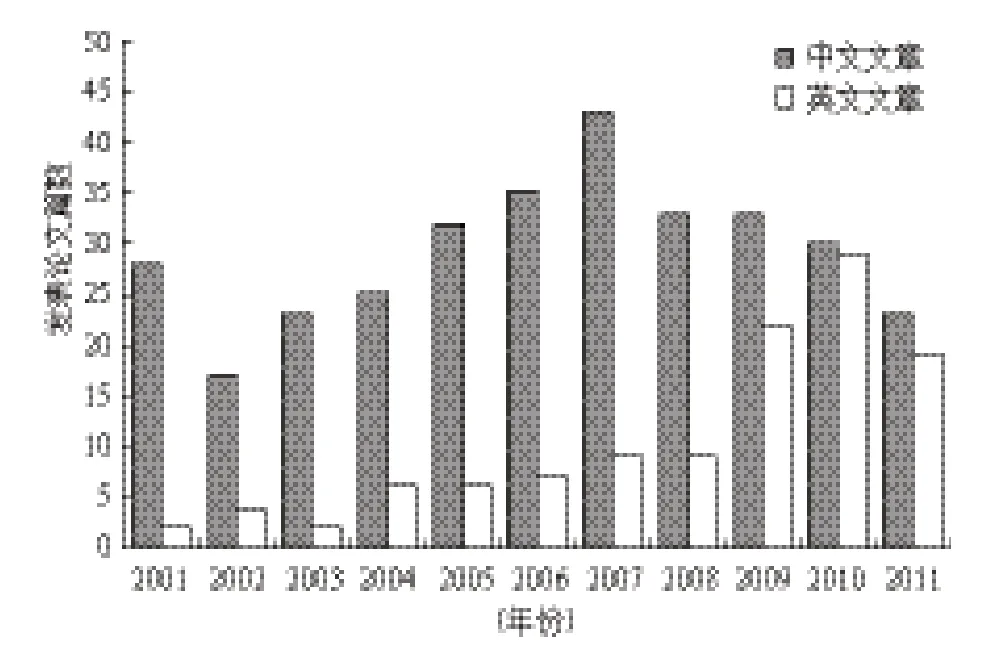
图1 我国外生菌根研究相关文献发表年份统计
对我国外生菌根真菌研究相关领域进行分析(图2):在针对外生菌根真菌资源调查、分离培养、分类鉴定、生理效应等方面的文章占大多数。在应用类相关文献中,林业造林育苗(菌根化幼苗、提高树木耐性)方面的文献占绝大多数;仅有极少数的文章涉及环境生物修复、生态系统效应等领域,而且此类文章很少涉及技术原理、分子及生理等机制。说明我国外生菌根真菌的研究面虽然广,但理论研究深度远远不足。
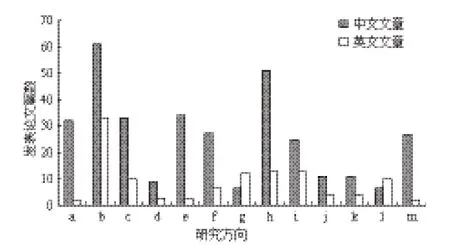
图2 我国外生菌根真菌研究相关内容分类统计
1 近10年我国外生菌根研究主要成果
1.1 外生菌根真菌资源调查、多样性、分类及分子鉴定
对于中国外生菌根真菌资源,有人进行了大量的调查[1-11],根据这些调查,我国现已确认与林木共生的外生菌根真菌约为28科,63属,近600种。而据不完全统计,全球约有5 000-6 000种外生菌根真菌[12]。说明我国还应继续重视并加强对外生菌根资源的调查力度,这对外生菌根真菌的资源开发利用和保护具有极其重要的意义。从调查发现的我国主要菌种来看,红菇科(Russulaceae),牛肝菌科(Boletaceae),口蘑科(Tricholomataceae)及马勃科(Lycoperdaceae)等分布较为广泛;从研究地理位置分析,东北和云南林区是主要的研究重点,而对于中、西部大面积林区的调查力度却显不足。
菌种分类鉴定以往常用形态学方法。在合适的外界条件下,外生菌根真菌可以在地上部分形成颜色、形状、大小各异的子实体,这是对外生菌根真菌进行分类鉴定的一个基本依据[13,14]。此外,利用切片、光学显微镜、扫描电镜、荧光显微镜(荧光素二醋酸染色后观测)等技术[15],对外生菌根真菌可以通过微观形态结构进行分类。
由于大部分外生菌根真菌并不在地面上形成子实体,而且不同的菌种可能会形成形态特征相似的子实体。因此,仅仅从形态学上对菌根真菌进行分类鉴定存在一定的局限性。随着分子生物学技术的发展,分子标记方法越来越被广泛采用。
国际上常利用PCR-RFLP区分外生菌根真菌不同种类,同时,还利用ITS序列测定分析的方法确定种名和遗传信息。RFLP技术与ITS序列分析技术相结合的方法,被广泛的应用于菌根真菌的鉴定和群落结构多样性的分析。我国在菌根真菌分子鉴定中也取得了一定的进展:rDNA在真菌的系统发育分析和分类鉴定中是很重要的靶序列,利用内转录间隔区(ITS)可以对外生菌根菌进行有效的鉴定[16-20],同时,RAPD分析技术[21]和单核苷酸多态性(SNPs)标记技术[22]也是外生菌根分离物真伪鉴定的一种快速可靠的方法。ITS和RAPD这两种分子标记鉴定方法的结合运用[23]是对外生菌根子实体组织分离的菌株进行准确鉴定的有效手段。分子生物学的鉴定方法具有可信度高、分类精确等优点,弥补了形态学研究方法的不足。利用分子鉴定手段并结合常规的形态学分类方法仍然是目前外生菌根真菌分类常用的分类方法。
1.2 外生菌根真菌分离、纯培养,以及外生菌根合成
菌根技术在人工造林方面的应用具有十分重要的意义,其中优良外生菌根真菌的分离筛选、纯培养以及菌根化培养条件的建立是该技术研究中的关键环节。
在菌种纯培养技术中,PDA和MMN是目前最常用的培养基。但是,单一的培养基毕竟存在一定的局限性,特别是对不同的菌种,其生长条件(如pH、温度、水势、营养物质种类和浓度等)有所不同[24]。因此,对于不同菌种实验室的纯培养有必要对其最适培养基、培养条件进行摸索研究[25]。姚庆智和闫伟[26]在实验室纯培养条件下,对油松菌根分离获得的11种外生菌根进行了营养生理代谢的研究,确定了最佳培养条件:在麦芽汁和Pachlewski培养基上生长,适宜温度为22.5-25℃,pH 以6.0-7.0为宜,C源以麦芽汁和葡萄糖为好,N源则以氨态N和有机态N中的蛋白胨为佳。与菌种单独纯培养相比,黄光文和张平[27]研究发现,利用愈伤组织和外生菌根真菌共同培养,能有效促进真菌菌丝的萌发和生长。
菌根苗造林能显著促进树苗的生长、提高造林成活率和保存率。菌根合成技术在外来植物引种、珍稀树种移栽等方面具有极其重要的意义,也一直是理论研究与应用研究所必须解决的问题。实验室常用的菌根合成培养基质常采用灭菌的蛭石、泥炭或两者的混合物。接种方式可以是菌液浇灌、或直接将根系与固体培养的活力高的菌丝接触(一般取菌斑外沿菌丝)、或者直接在已经菌根化的“母苗”周围播种或移栽小苗的方式接种。我们在近期菌根化条件优化试验中发现,使用贫瘠、疏松、透水性好的培养基质,采取对40 d龄松树幼苗剪去主根然后接种的方式,可获得近100%侵染率的菌根化苗(方法待发表)。孙玥等[28]研究发现,外源营养元素N的施用往往会导致外生菌根真菌对落叶松根系侵染率的下降,因此要严格控制N浓度。在野外田间菌根合成过程中,生态、土壤条件及宿主等因素[29]都能影响外生菌根的合成。非宿主植物与菌根真菌也存在着相互作用[1],如虎榛子与油松一起接种外生菌根真菌时,虎榛子的存在增加了油松的菌根侵染率[30]。另外,丛枝菌根真菌与外生菌根真菌之间也存在相互作用及混合接种效应[31]。在生产应用中,利用外生菌根真菌菌剂,如工业液体发酵法生产的厚环乳牛肝菌(Suillus grevillei)固体菌剂与彩色豆马勃(Pisolithus tinctorlus)孢子粉剂混合菌剂可以提高林木的成活率,并能多年发挥菌根的效应[32]。
1.3 外生菌根真菌的生态效应
选择优良菌种和菌剂培养工艺,对绿化荒山、提高森林覆盖率具有重要意义。通过N-15稳定同位素示踪技术,发现菌根菌丝体形成的菌丝网络结构(菌丝桥)为植物间营养元素的传递提供了媒介[30,33]。如果将虎榛子与油松一起接种外生菌根,菌根真菌不仅促进了两者的生长、改善了营养状况,还改变了植物根际的pH以及磷酸酶活性[30]。张海涵等[34]发现随着菌根侵染率的提高,根际土壤微生物功能多样性增加,有利于改善黄土高原退化土壤的微生物群落结构。外生菌根菌在阻止或延缓科尔沁沙地南缘樟子松人工林的衰退中表现出积极的作用[35]。外生菌根真菌还可以刺激宿主植物根系酸性磷酸酶和硝酸还原酶活性,特别是在严重酸雨(pH<3.0)发生时[36]。有研究发现,某些菌根真菌(Paxillus involutu和Laccaria laccata)可以通过改变植株根际化学效应来控制高等物种间的相互作用[37]。这些研究结果在森林植被恢复,以及维护生态系统稳定中具有重要的应用价值。
其次,《胭脂扣》本身具有浓郁的传统色彩。作为爱情信物的“胭脂扣”既是爱情的见证者又是历史的缩影。情感的变迁、香港的流变近半个世纪的历史哪一点不是凝聚在这枚胭脂扣上?“摸摸口袋,有件硬物,赫然是那枚胭脂匣子,她不要了!我一想也把它扔在夜路上。车子绝尘而去,永不回头。”三十年代珍贵而流行的定情物,到了八十年代竟成了私人小店内的幻影,甚至最终落得被扔掉的命运。历史亦是如此,所有历史不过都是当下史,过了“当下”似乎早已没有存在的意义与价值,它能呈现给他人的除了时间的变迁再无其他。
1.3.1 外生菌根真菌对树木生长、生理的影响 接种外生菌根能提高幼苗成活率、促进苗木的生长,对此人们已有广泛的共识[38-41]。与丛枝菌根真菌混合接种,也能表现出对幼苗的促生作用[31]。
菌根体系所具有的菌丝体系,促进了宿主植物根系对水分、养分的吸收和传输[42],特别是对外界氮、磷、钾[39,40]的代谢利用。菌根化能提高松树幼苗针叶中的叶绿素含量及叶绿素荧光参数(量子产量、电子传递速率)[43],提高幼苗净光合速率,降低蒸腾速率[38,44]。外生菌根真菌还可以刺激宿主植物合成生长素、细胞分裂素等内源激素,或产生一些次生代谢产物(如酶类、有机酸)等来调节植物体内的激素和代谢平衡,促进植物生长[45]。
1.3.2 提高植物耐性方面 众多研究表明,外生菌根菌的存在增强了树木抵抗重金属[46]、干旱[24]、盐[47]、病害[48,49]等胁迫的能力。
离体条件培养下,外生菌根菌Boletus edulis有很强的对铜、镉吸收积累能力,显示出较好的重金属耐受性。菌丝体对重金属元素铜、镉具有不同的积累模式,根据重金属种类,有效地调节生长微环境[50]以减少植物根际区重金属的生物有效性,增强植物对重金属铜、镉的耐性[51,52]。廖继佩等[46]认为,外生菌根菌丝体能与土壤中的重金属结合,限制重金属向宿主植物的迁移,这也是其增加宿主植物对重金属耐性的一个重要原因。李婷等[53]研究发现,樟子松在不同浓度Cu和Zn条件下,接种牛肝菌(Suillus bovinus)的植株不仅生物量有所增加,而且Cu和Zn的含量也增加了,但主要集中在根部,被根部菌丝体系吸收固持而减少向植株地上部转运。根据近期本实验室土培试验结果:在接种Pisolithus tinctorius,Cenococcum geophilum的松树幼苗时,当外界施加高浓度Cu、Cd时,在处理的12 h内,接菌苗地上部可以检测到大量积累的重金属;而非菌根化苗在处理的12 h内地上部重金属并没有明显积累,只有在处理24 h后地上部分才检测到明显增加的重金属,说明外生菌根真菌具有加速重金属向植物体内输送的特性。在另一个长期土培试验中,采用多次向土壤浇灌铜溶液的处理方法,与非菌根化植物相比,菌根化幼苗最初出现毒害症状,更易受到铜毒害。因此推断,由于菌丝增加了根系的吸收面积,菌丝束的毛细作用可能会加速重金属向植物根系富集和向植物体内的转运。当外界有毒重金属量有限时,植物往往通过改善植物本身的生长从而稀释重金属对植物的毒害;但当外界有毒重金属过量且持续胁迫时,菌丝促进重金属在植物体内的富集反而会加剧植物的毒害(数据待发表)。因此,菌丝可能并不会起到一个屏障作用来阻滞重金属向植物体内的积累和迁移。
盐胁迫对植物的伤害机理主要是离子毒害,渗透胁迫和营养亏缺,接种外生菌根真菌可以增加宿主可溶性糖类含量,提高渗透压,缓解渗透胁迫[54]。菌根真菌还可以通过增加植物对P等矿质营养元素的吸收,改善盐胁迫带来的营养亏缺、改变植物体内的离子平衡,同时增加水分的吸收和利用效率,从而缓解植物生理干旱[55]。
在外生菌根真菌提高林木的抗病害机理研究方面,张茹琴等[56]就灰鹅膏菌、血红铆钉菇、灰鹅膏菌和绒黏盖牛肝菌对油松幼苗的抗猝倒病机制进行研究发现,抗病性不仅与侵染率有关,还与形成的典型菌套和哈氏网结构的物理屏障有关,菌套能阻碍病原菌穿透植物根部,这可能是菌根化油松抗病机制之一。除了特殊结构的物理屏障外,外生菌根真菌还能产生化学抑制剂(挥发性化学抑制剂、抗病有关的酶类)与病原菌进行营养竞争[57]。通过离体培养试验,Tang等[58]发现外生菌根真菌(Suillus laricinus,Suillus tomentosus)与水稻纹枯病共培养,可以有效抑制其病原体的生长。
1.3.3 环境污染修复 近年来随着土壤污染问题的日益严重,菌根真菌对促进有机污染物的降解、转化中凸显出积极作用的一面:在柴油[59]、矿物油[60]污染土壤的修复;氯酚类[52,61]污染土壤的修复;有机农药和塑料薄膜增塑剂-酞酸酯类(PAEs)[48,62]污染土壤的降解修复等研究中均尝试了菌根生物技术的应用,并且均取得了良好的成效。Huang 等[63]发现Gomphidius viscidus 菌种,不论活的还是灭活的菌丝体,对多环芳烃都具有很高的吸附能力。外生菌根真菌还表现出很强的适应能力,通过调节自身的生理过程来适应外界生长微环境的变化。在露天煤矿脆弱生态地段的生态修复中,外生菌根真菌也具有出一定的应用价值[65]。
1.3.4 次生代谢物效应 有研究表明,纯培养条件下的Suillus grevillei、Suillus bovines、Rhizopogon luteolus和Pisolithus tinctorius均能产生除了脱落酸以外的其他4种植物激素[68]。外生菌根真菌产生的植物生长物质(乙烯、玉米素和赤霉素等)[69]在菌根的形成以及促进宿主植物的生长方面起到重要作用,同时产生的拮抗活性物质对立枯丝核菌(R. solani)具有一定的抗性[70],外生菌根真菌还能分泌多肽类毒素[71]。彩色豆马勃(Pisolithus tinctorius)子实体的丙酮抽提物对狼尾草和油菜幼苗生长均有抑制作用,而其纯化得到的豆马勃内酯和麦角甾醇也能抑制稗草幼苗的生长[72]。外生菌根真菌分泌物在生物防治方面的应用潜力还有很大的开发空间。
1.4 环境因素对外生菌根真菌生长、代谢和分布的影响
如上所述,外生菌根真菌在生态系统中承担重要角色,同时,外界众多的环境因子(如pH、温度、重金属和盐浓度,以及营养元素的供给等)也影响菌根真菌的生长和代谢。
外生菌根真菌往往喜欢偏酸性的环境,最适pH值一般在4-6范围。外生菌根真菌在不适合的土壤pH、不适的温度(<5℃和>37℃)以及干旱等逆境条件下不能生长,甚至死亡,这也是导致沙地樟子松人工林衰退的原因之一[73]。张茹琴等[74]探讨了重金属以及不同pH对黏盖牛肝菌(Suillus bovinus)、褐环黏盖牛肝菌(Suillus luteus)、褐黄牛肝菌(Suillus luridus)等3种菌根真菌生长的影响。黏盖牛肝菌对锌的耐性较强,而铜(10-1 000 mg/L)对3种外生菌根真菌的生长均有抑制作用,低浓度的铅对褐环黏盖牛肝菌(铅浓度 < 600 mg/L)、褐黄牛肝菌和黏盖牛肝菌(铅浓度 < 200 mg/L)的生长具有促进作用。据本实验室进行的MMN培养基液体培养耐铜性试验结果(铜处理浓度:0.013、1、10、50、125和250 μmol/L),低浓度铜(1、10 μmol/L)处理具有促进菌丝生长的作用,不同菌种对铜的耐性差异很大,如Suillus granulatus-1对铜的耐性最强(EC50>>250 μmol/L),Tricholoma imbricatum、Laccaria laccata-1等也有很高的耐性(EC50>250 μmol/L),而Rhizopogon sp.-4则最为敏感(EC50=50 μmol/L)(数据待发表)。Huang和Tao[75]研究发现,牛乳牛肝菌在含有较高浓度的Zn或Cu的营养液中培养时,其己糖激酶和磷酸果糖激酶的活性受到抑制,进而抑制菌丝体的生长和呼吸。
Liang等[76]通过蛋白质组学方法,利用双向电泳和质谱技术分析了菌种Boletus edulis 对盐胁迫的响应发现:有14个蛋白点上调,8个蛋白点下调,并对其中与蛋氨酸和S-腺苷甲硫氨酸合成、糖酵解、DNA修复、细胞周期调控相关的16个蛋白点与菌种抗逆性和适应性进行了探讨。Tang等[77]在离体条件下对3种外生菌根真菌的耐盐(NaCl)性进行了比较,发现Boletus luridus 具有很高的耐盐性,而Suillus bovinus则最为敏感。本实验室近期对18个外生菌根真菌进行耐盐碱性研究结果显示,Cenococcum geophilum表现出最强的耐盐碱性(数据待发表)。
很多菌株均以NH4+-N为最适宜生长的氮源。对于钾元素,其浓度将不同程度地影响真菌菌丝对其他营养元素的吸收,在钾浓度中等(0.65 g/L)条件下生长最好;低浓度钾将促进菌根真菌氢离子的分泌,活化土壤中的无效钾[78]。施用低浓度的磷也会促进外生菌根真菌松乳菇和双色蜡蘑的草酸、氢离子和磷酸酶的分泌[79]。
气候与季节等环境因素影响外生菌根的分布。树木的外生菌根对特定的季节有一定的生态适应性[6]:3-4月很少产生,夏初的5-6月开始产生,夏季7-8月达到最多,而后逐渐减少;树种、植被、林龄及林分密度对外生菌根真菌分布也会产生影响。
1.5 菌根食用菌开发
部分外生菌根真菌可供人类食用,其数量约占整个食用菌资源的一半[80],我国野生可食用的外生菌根真菌资源极其丰富,约占中国食用菌总数的70%[81],主要分布在云南、四川、贵州等地。如松口蘑(Tricholoma matsutake)[82]、鸡油菌(Cantharellus cibarius)、松乳菇(Lactarius deliciosus)、美味牛肝菌(Boletus edulis)、灰肉红菇(Russula griseocarnosa)[14]、中 国 块 菌(Chinese truffle Tuber sinense)[83]等。其中可供人类食用的子实体大多为珍贵食用菌,具有较高的营养价值,而且部分还有药用价值[84],价格昂贵,市场潜力很大。但仅有少部分菌种可以进行人工培育。
2 展望
综上所述,我国外生菌根研究面相对广泛。与国际相比,我国外生菌根真菌研究起步虽较晚,但发展迅速。研究重点主要集中在资源调查、菌根合成以及造林应用等方面。
在资源调查方面,需要进一步加强对各个自然区域菌根资源的调查力度,利用国际上公认的分子鉴定技术对菌种进行分类鉴定。完善菌种的分离、纯培养的技术,建立完善的菌种资源库、资料库和DNA文库,并加强科研单位菌种研究的横向交流。在分子水平上研究菌根宿主与外生菌根真菌多样性的关系,确定菌种与生长环境、树种之间的关系。针对我国主要造林树种和地理条件,充分利用外生菌根资源,为我国林业发展提供有力的技术支持。
食用菌国内的应用还基本局限在野生资源直接利用上,引种驯化是需要解决的难题。首先要了解菌种的生态习性以及其生活史,进行各种培养条件的筛选,利用共生林木进行林地栽培,还可利用现代生物技术,进行杂交育种,获得优良可用于生产的菌种。
针对外生真菌代谢物质的研究,需要加强对有机酸类和多肽类,以及一些胞外分泌酶等物质的分离、鉴定和效应的研究,探讨这些物质在食品、医药等行业的应用潜力。
在环境污染修复方面,加强利用菌根化苗对环境污染土壤进行生物修复的研究力度:如加强菌根真菌降解有机污染物作用机制的研究、利用菌根真菌辅助植物修复环境有机污染物,以及菌根真菌辅助植物钝化修复等方面。外生菌根真菌的耐重金属、耐盐碱胁迫机理的研究也需要深入,以期为利用外生菌根真菌辅助植物修复环境污染、提高植物耐盐性提供理论依据。我国有关外生菌根真菌生态效用方面的研究相对薄弱,菌种对濒危树种保护、对维护森林生态系统的稳定以及对全球气候变化的影响、对生物圈碳固定等方面的研究也亟待加强。
[1] Liang Y, Guo LD, Ma KP. Spatial pattern of the most common latestage ectomycorrhizal fungi in a subtropical forest in Dujiangyan, Southwest of China[J]. Acta Botanica Sinica, 2004, 46(1):29-34.
[2] 李海波, 吴学谦, 魏海龙, 等. 浙江丽水地区外生菌根菌资源调查初报[J]. 中国食用菌, 2005, 24(5):10-14.
[3] 柯丽霞, 刘必融. 黄山地区松树林外生菌根菌资源及生态分布[J]. 应用生态学报, 2005, 16(3):455-458.
[4] 白淑兰, 刘勇, 周晶, 等. 大青山外生菌根真菌资源与生态研究[J]. 生态学报, 2006, 26(3):837-841.
[5] 罗国涛, 王健祥, 袁玮. 凯里市马尾松外生菌根菌的调查[J]. 中国食用菌, 2007, 26(2):12-14.
[6] 于富强, 肖月芹, 刘培贵. 云南松(Pinus yunnanensis)林外生菌根真菌的时空分布[J]. 生态学报, 2007(6):2325-2333.
[7] 刘朝贵, 马根艳, 聂和平, 等. 武陵山脉七曜山区大型真菌资源调查[J]. 食用菌学报, 2009, 16(2):77-83.
[8] 图力古尔, 陈今朝, 王耀, 等. 长白山阔叶红松林大型真菌多样性[J].生态学报, 2010, 30(17):4549-4558.
[9] 宋福强, 刘远开, 杜春梅, 等. 小兴安岭红松外生菌根真菌资源及响应面法优化Suillus grevillei 培养基[J]. 自然资源学报, 2011, 26(3):440-449.
[10] 樊永军, 闫伟, 王黎元. 贺兰山地区青海云杉外生菌根的形态[J]. 林业科学, 2011, 47(6):108-113.
[11] Zhang Y, Xu YZ, Zheng ZX, et al. Analyses of diversity and distribution characteristics of macro-fungi in Huafo Mountain Nature Reserve of Yunnan Province[J]. Journal of Plant Resources and Environment, 2012, 21(1):111-117.
[12] Molina R, Massicotte HB, Trappe JM. Ecological role of specificity phenomena in ectomycorrhizal plant-communities potentials for interplant linkages and guild development[J]. Mycorrhizas in Ecosystems, 1992:106-112.
[13] 刘润进, 陈应龙. 菌根学[M]. 北京:科学出版社, 2007.
[14] Wang XH, Yang ZL, Li YC, et al. Russula griseocarnosa sp nov(Russulaceae, Russulales), a commercially important edible mushroom in tropical China:mycorrhiza, phylogenetic position and taxonomy[J]. Nova Hedwigia, 2009, 88(1-2):269-282.
[15] Qian XM, Kottke I, Oberwinkler F. Activity of different ectomycorrhizal types studied by vital fluorescence[J]. Plant and Soil, 1998, 199(1):91-98.
[16] Li YC, Yang ZL, Tolgor B. Phylogenetic and biogeographic relationships of Chroogomphus species as inferred from molecular and morphological data[J]. Fungal Diversity, 2009(38):85-104.
[17] Vaario LM, Suzuk K. Ectomycorrhizal synthesis between Abies firma roots/callus and Laccaria bicolor strain[J]. Acta Botanica Sinica, 2004, 46(1):63-68.
[18] Xie XD, Liu PG. Species diversity of the genus tomentella mycorrhizae-forming fungi on Pinus yunnanensis seedlings and the mycorrhizal morphology[J]. Journal of Fungal Research, 2011, 9(4):224-231, 243.
[19] Li H, Guo L, Zhou GY, et al. Analysis of genetic diversity of Lactarius hatsudake in South China[J]. CanadianIournal Canadian Journal of Microbiology, 2011, 57(8):661-666.
[20] Wang Q, Gao C, Guo LD. Ectomycorrhizae associated with Castanopsis fargesii(Fagaceae)in a subtropical forest, China[J]. Mycological Progress, 2011, 10(3):323-332.
[21] 曾东方, 罗信昌. 应用RAPD分析快速鉴定外生菌根蘑菇分离物的真伪[J]. 林业科学, 2001, 37(6):59-65.
[22] Xu JP, Guo H, Yang ZL. Single nucleotide polymorphisms in the ectomycorrhizal mushroom Tricholoma matsutake[J]. Microbilogy-SGM, 2007(153):2002-2012.
[23] 李海波, 吴学谦, 魏海龙, 等. 几种森林大型真菌纯培养菌种的RAPD及ITS分子标记鉴定[J]. 林业科技, 2007, 43(12):94-100.
[24] Zhu JJ, Li FQ, Xu ML, et al. The role of ectomycorrhizal fungi in alleviating pine decline in semiarid sandy soil of northern China:an experimental approach[J]. Annals of Forest Science, 2008, 65(3):304.
[25] 于富强, 纪大干, 刘培贵. 云南松外生菌根真菌分离培养研究[J]. 植物研究, 2003, 23(1):66-71.
[26] 姚庆智, 闫伟. 11株外生菌根真菌菌株纯培养营养生理特性的研究[J]. 内蒙古大学学报:自然科学版, 2005, 36(2):186-191.
[27] 黄光文, 张平. 共生法培养外生菌根菌菌丝体[J]. 中国食用菌, 2005, 24(3):16-17.
[28] 孙玥, 全先奎, 贾淑霞, 等. 施用氮肥对落叶松人工林一级根外生菌根侵染及形态的影响[J]. 应用生态学报, 2007, 18(8):1727-1732.
[29] 卢丽君, 白淑兰, 王静, 等. 外生菌根合成的条件及形成机制[J]. 微生物杂志, 2005, 3(2):84-87.
[30] Bai SL, Li GL, Liu Y, et al. Ostryopsis davidiana seedlings inoculated with ectomycorrhizal fungi facilitate formation of mycorrhizae on Pinus tabulaeformis seedlings[J]. Mycorrhiza, 2009, 19(6):425-434.
[31] Chen YL, Brundrett MC, Dell B. Effects of ectomycorrhizas and vesicular-arbuscular mycorrhizas, alone or in competition, on root colonization and growth of Eucalyptus globulus and E. urophylla[J]. New Phytologist, 2000, 146(3):545-556.
[32] 赵志鹏, 郭秀珍, 张良谱. 外生菌根菌剂在油松育苗造林中的应用[J]. 林业科学, 1993, 29(5):401-407.
[33] He XH, Xu MG, Qiu GY, et al. Use of15N stable isotope to quan-tify nitrogen transfer between mycorrhizal plants[J]. Journal of Plant Ecology, 2009, 2(3):107-118.
[34] 张海涵, 唐明, 陈辉, 等. 黄土高原5种造林树种菌根根际土壤微生物群落多样性研究[J]. 北京林业大学学报, 2008, 30(3):85-90.
[35] 朱教君, 康宏樟, 许美玲, 等. 外生菌根真菌对科尔沁沙地樟子松人工林衰退的影响[J]. 应用生态学报, 2007, 18(12):2693-2698.
[36] Kong FX, Zhou CL, Liu Y. Biochemical and cytological responses of ectomycorrhizae in Pinus massoniana to artificial acid rain[J]. Chemosphere, 1998, 37(1):179-187.
[37] Ren ZS, Mallik AU. Selected ectomycorrhizal fungi of black spruce(Picea mariana)can detoxify phenolic compounds of Kalmia angustifolia[J]. J Chem Ecol, 2006, 32(7):1473-1489.
[38] 阎秀峰, 王琴. 接种外生菌根对辽东栎幼苗生长的影响[J]. 植物生态学报, 2002, 26(6):701-701.
[39] Chen YL, Dell B, Malajczuk N. Effect of Scleroderma spore density and age on mycorrhiza formation and growth of containerized Eucalyptus globulus and E. urophylla seedlings[J]. New Forests, 2006, 31(3):453-467.
[40] Song RQ, Ju HB, Qi JY, et al. Effect of ectomycorrhizal fungi on seedliing growth of Mongol Scotch Pine[J]. Journal of Fungal Research, 2007, 5(3):142-145.
[41] 宋瑞清, 王锋,冀瑞卿, 祁金玉. 红皮云杉外生菌根菌单接种及混合种对苗木生长的影响[J]. 微生物学报, 2007, 47(6):1091-1094.
[42] 郑玲, 吴小芹. 黑松菌根共生体中真菌液泡形态构架及其活力[J]. 植物生态学报, 2008, 32(4):932-937.
[43] 高悦, 吴小芹. 6种外生菌根菌对3种松苗叶绿素含量及叶绿素荧光参数的影响[J]. 南京林业大学学报:自然科学版, 2010, 34(6):9-12.
[44] 刘现刚, 郭素娟, 赵菡, 等. 不同菌根菌对栓皮栎苗木光合及蒸腾特性的影响[J]. 浙江林业科技, 2010, 30(6):1-5.
[45] Yu FQ, Liu PG. Reviews and prospects of the ectomycorrhizal research and application[J]. Acta Ecol Sin, 2002, 22(12):2217-2226.
[46] 廖继佩, 林先贵, 曹志洪. 内外生菌根真菌对重金属的耐受性及机理[J]. 土壤, 2003, 35(5):370-377.
[47] 黄艺, 姜学艳, 梁振春, 等. 外生菌根真菌接种和施磷对油松苗抗盐性的影响[J]. 生态环境, 2004, 13(4):622-625, 640.
[48] 梁振春, 黄艺. 4种外生菌根真菌对滴滴涕的耐受性及生理响应[J]. 环境科学研究, 2006, 19(4):36-41.
[49] 刘润进, 李俊喜. 菌根真菌菌剂防治作物土传病害潜力分析[J]. 植物病理学报, 2007, 37(1):1-8.
[50] 李婷, 黄艺, 黄志基. 重金属胁迫下外生菌根真菌Boletus edulis重金属积累分配与生长微环境变化[J]. 生态环境, 2005, 14(6):870-875.
[51] 黄艺, 李婷, 费颖恒. 外生菌根真菌对油松幼苗根际土壤重金属赋存的影响[J]. 生态与农村环境学报, 2007, 23(3):70-76.
[52] 黄艺, 杨青, 敖晓兰. 外生菌根真菌对五氯芬的耐受性及生理响应[J]. 环境科学学报, 2008, 28(10):2078-2083.
[53] 黄艺, 陶澍, 陈有鑑, 张学青. 外生菌根对欧洲赤松苗(Pinus sylvestris)Cu、Zn积累和分配的影响[J]. 环境科学, 2000, 21(2):1-6.
[54] Luo ZB, Li K, Jiang XN, et al. Ectomycorrhizal fungus(Paxillus involutus)and hydrogels affect performance of Populus euphratica exposed to drought stress[J]. Annals of Forest Science, 2009, 66(1):106.
[55] Xu D, Dell B, Malajczuk N, et al. Effects of P fertilization and ectomycorrhizal fungal inoculation on early growth of eucalypt plantations in southern China[J]. Plant and Soil, 2001(233):47-57.
[56] 张茹琴, 唐明, 张海涵. 四种外生菌根真菌对油松幼苗的抗猝倒病和促生作用[J]. 菌物学报, 2011, 30(5):812-816.
[57] Sen R. Multitrophic interaction between a Rhizoctonic sp. and mycorrhizal fungi affect Scots pine seeding performance in nursery soil[J]. New Phytologist, 2001, 152(3):543-553.
[58] Tang M, Zhang RQ, Chen H, et al. Induced hydrolytic enzymes of ectomycorrhizal fungi against pathogen Rhizoctonia solani[J]. Biotechnology Letters, 2008, 30(10):1777-1782.
[59] 礼晓, 黄艺, 魏志成. 两种纯培养外生茵根真菌对柴油的降解效率[J]. 应用生态学报, 2008, 19(7):1579-1584.
[60] 程国玲, 李培军, 王风友, 等. 几种纯培养外生菌根真菌对矿物油的降解效果[J]. 中国环境科学, 2003, 23(1):74-76.
[61] 黄艺, 敖晓兰, 赵曦. 五氯酚生物降解机理与外生菌根真菌对五氯酚可降解性[J].生态环境, 2006, 15(5):1080-1085.
[62] Huang Y, Zhao X, Luan SJ. Uptake and biodegradation of DDT by 4 ectomycorrhizal fungi[J]. Science of the Total Environment, 2007, 385(1-3):235-241.
[63] Huang Y, Zhang SY, Lv MJ, et al. Biosorption characteristics of ectomycorrhizal fungal mycelium for Anthracene[J]. Biomedical and Environmental Sciences, 2010, 23(5):378-383.
[64] 韩桂云, 孙铁珩. 外生菌根真菌在大型露天煤矿生态修复中的应用研究[J]. 应用生态学报, 2002, 13(9):1150-1152.
[65] 黄艺, 黄志基. 外生菌根与植物抗重金属胁迫机理[J]. 生态学杂志, 2005, 24(4):422-427.
[66] 黄艺, 彭博, 李婷, 等. 外生菌根真菌对重金属铜镉污染土壤中油松生长和元素积累分布的影响[J]. 植物生态学报, 2007, 31(5):923-92.
[67] 黄艺, 黄志基. 外生菌根与植物抗重金属胁迫机理[J]. 生态学杂志, 2005, 24(4):422-427.
[68] 王有智, 黄亦存. 四种外生菌根真菌产生植物激素的研究[J]. 微生物学通报, 1997, 24(2):72-74.
[69] 简利茹, 李哲斐, 韩青梅, 等. 固相萃取-HPLC测定外生菌根真菌产生植物激素IAA 和GA3[J]. 西北农业学报, 2011, 20(9):165-168.
[70] 赵志鹏, 郭秀珍. 外生菌根真菌纯培养中生理活性物质代谢的研究[J]. 林业科技, 1990, 26(5):465-469.
[71] 郭学武, 汪国轮, 龚建华, 等. 三种鹅膏菌培养条件及所产肽类毒素的比较研究[J]. 菌物学报, 2008, 27(5):745-756.
[72] 曾任森, 骆世明, 石木标, 等. 彩色豆马勃子实体的化感作用及其化感物质的分离鉴定[J]. 应用生态学报, 1999, 10(2):206-208.
[73] 朱教君, 许美玲, 康宏樟, 等. 温度、pH及干旱胁迫对沙地樟子松外生菌根菌生长影响[J]. 生态学杂志, 2005, 24(12):1375-1379.
[74] 张茹琴, 唐明, 张峰峰, 等. 酸碱度和重金属对3种外生菌根真菌生长的影响[J]. 北京林业大学学报, 2008, 30(2):113-118.
[75] Huang Y, Tao S. Effects of excessive Cu or Zn on biomass respiration and activities of glycolytic enzymes in mycelia of mycorrhizal fungus Suillus bovines[J]. Acta Photophysiologica Sinica, 2001, 27(4):303-308.
[76] Liang Y, Chen H, Tang M, Shen S. Proteome analysis of an ectomycorrhizal fungus Boletus edulis under salt shock[J]. Mycological Research, 2007(111):939-946.
[77] Tang M, Sheng M, Chen H, et al. In vitro salinity resistance of three ectomycorrhizal fungi[J]. Soil Biology and Biochemistry, 2009, 41(5):948-953.
[78] 袁玲, 方德华, 汪智慧, 等. 钾对外生菌根真菌的分泌作用及氮、磷、钾含量的影响[J]. 生态学报, 2001, 21(2):254-258.
[79] 薛小平, 张深, 李海涛, 等. 磷对外生菌根真菌松乳菇和双色蜡蘑草酸、氢离子和磷酸酶分泌的影响[J]. 菌物学报, 2008, 27(2):193-200.
[80] Liu RJ, Liu ZJ. Retrospects of 40 years of studies on mycorrhizasand prospects in China[J]. Acta Phytopathol Sin, 1998, 28(3):201-208.
[81] 戴玉成, 周丽伟, 杨祝良, 等. 中国食用菌名录[J]. 菌物学报, 2010, 29(1):1-21.
[82] Xu JP, Sha T, Li YC, et al. Recombination and genetic differentiation among natural populations of the ectomycorrhizal mushroom Tricholoma matsutake from southwestern China[J]. Molecular Ecology, 2008, 17(5):1238-1247.
[83] Liu RS, Li DS, Li HM, et al. Response surface modeling the significance of nitrogen source on the cell growth and Tuber polysaccharides production by submerged cultivation of Chinese truffle Tuber sinense[J]. Process Biochemistry, 2008, 43(8):868-876.
[84] 戴玉成, 杨祝良. 中国药用真菌名录及部分名称的修订[J]. 菌物学报, 2008(27):801-824.
猜你喜欢
军事文摘(2021年18期)2021-12-02 01:28:04
科学(2020年2期)2020-08-24 07:56:56
园林科技(2020年2期)2020-01-18 03:28:26
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
微生物与感染(2015年5期)2015-12-08 07:03:20
实用手外科杂志(2015年3期)2015-08-27 01:53:08
山西农经(2015年7期)2015-07-10 15:00:57
食品工业科技(2014年15期)2014-03-11 18:17:52
食品工业科技(2014年9期)2014-03-11 18:15:28
环境与可持续发展(2013年6期)2013-03-11 16:21:51
