
中国褐毛藻属Halothrix的分类学研究
2013-12-23 05:13:24黄冰心栾日孝丁兰平政晓英孙国栋谭华强
海洋科学 2013年4期
黄冰心 , 栾日孝, 丁兰平, 政晓英, 孙国栋 谭华强
(1. 汕头大学, 广东 汕头 515063; 2. 中国科学院 海洋研究所, 山东 青岛 266071; 3. 大连自然博物馆, 辽宁 大连 116023)
褐毛藻属Halothrix隶属于褐藻门Phaeopyta, 索藻目 Chordariales, 短毛藻科 Elachistaceae, 由Reinke(1888: 19)[1]建立。其主要特征为藻体丝状, 数毫米高, 束状, 基部由藻丝交织成固着器; 藻体向上生有单列细胞的直立丝体(同化丝), 直立丝体在基部分枝, 可分为长同化丝和短同化丝(侧丝); 长同化丝长而粗, 短同化丝短而细, 着生于长同化丝的基部侧面, 常多回分枝; 色素体盘状, 多数; 多室囊无柄, 在同化丝中上部形成; 模式种为H. 1umbricalis(Kuetzing) Reinke。本属在国际上已报道了9个种及变型[2-7], 其中东亚报道了6个种[3,6], 中国报道了其中的2个种, 即褐毛藻H.lumbricalis(Kuetzing) Reinke[8-11]和可疑褐毛藻H. ambiguaYamada[12]。
为了完成“中国海藻志 褐藻门第一册”的编研工作, 作者自1998~2003年间对中国沿海多个地区开展了野外调查, 收集了一些该属的样品。结合我国的过去报道, 经分类与鉴定研究, 共发现我国褐毛藻属物种4个, 分别为可疑褐毛藻H. ambiguaYamada、细弱褐毛藻H. gracilisLuan et Ding sp. nov.、褐毛藻H. lumbricalis(Kuetzing) Reinke和垫状褐毛藻H. pulvinataLuan et Ding sp. nov., 其中新种2个。本文对这4个种进行了较详细的描述, 其结果在一定程度上丰富了我国大型海洋褐藻的多样性。本研究标本存放于中国科学院海洋研究所海洋生物标本馆(AST)。
1 材料与方法
1.1 实验材料
1998~2003年间作者在我国沿海野外现场调查采集的标本。
1.2 实验方法
1.2.1 外形特征比较
主要包括藻体颜色、基部情况、同化丝、繁殖器官等特征。
1.2.2 显微制片
实验材料经徒手处理后制成显微镜下观察的薄片。
1.2.3 物种鉴定
通过分类特征的对比及相关数据的比较, 结合文献报道鉴定物种。
1.2.4 绘图
利用显微镜和解剖镜的自配描绘装置对物种特征绘图。
2 研究结果
2.1 可疑褐毛藻(图1)
Halothrix ambiguaYamada 1928[13]: 513; Okamura 1936[14]: 148; Noda 1971[12]: 1458; Yoshida et al. 1998[3]: 255。

图1 可疑褐毛藻Halothrix ambigua Yamada Fig.1 Halothrix ambigua Yamada
特征描述: 藻体黄褐色, 密集丛生, 高 5~10 mm。固着器由假根状丝体交织而成。直立丝体(同化丝)可分成长同化丝和短同化丝(侧丝)。长同化丝粗而长, 细胞长多小于宽; 下部细胞长12.5~20 µm, 宽15~40 µm, 长宽比为0.4~1; 中部细胞长20~60 µm, 宽50~75 µm, 长宽比为0.3~1; 向上渐细, 顶细胞长15~25 µm, 宽13~25 µm, 长宽比为0.6~1.4。短同化丝着生于长同化丝的侧基部, 基部分枝, 棒状, 多弯曲, 长120~200 µm, 由7~12个细胞组成, 细胞横壁处缢缩呈念珠状; 中、下部细胞长10~20 µm, 宽5~7 µm, 长宽比为2~4; 顶细胞亚球形, 长10~20 µm, 宽10~15 µm。多室囊在长同化丝上部, 间生或顶生, 长短不一, 长100~300 µm, 宽50~75 µm。单室囊着生于短同化丝的基部或顶端; 着生于基部的为长卵形, 长50~100 µm, 宽25~40 µm; 着生于枝端的为卵形, 长18~40 µm, 宽18~30 µm。腊叶标本紧密附着于标本纸上。
习性和产地: 在低潮带的大石沼中, 附着于海草的虾海藻属Phyllospadix植物体上。产于辽宁大连、长海县。
国外分布: 日本, 朝鲜岛。
模式标本产地: 日本青森县(Aomori-ken, Japan)。
2.2 细弱褐毛藻(图2)
Halothrix gracilisLuan et Ding sp. nov.。
特征描述: 藻体褐色, 丛生, 高5~9 mm, 附着于其他海藻藻体表面。基部由匍匐状丝体组成基盘。直立丝体生于匍匐丝体上,下部细胞生有假根和短同化丝。短同化丝长100~250 µm, 由10~17个细胞组成, 细胞长 10~25 µm, 宽 10~12 µm, 长宽比为1~2.5。长同化丝细弱, 长0.5~0.8 cm, 中部稍粗, 向两端渐细; 下部细胞长12~20 µm, 宽20~38 µm, 长宽比为0.4~1; 中部细胞长45~55 µm, 宽35~55 µm, 长宽比为0.9~1.4; 上部细胞长25~75 µm, 宽15~25 µm, 长宽比为1.3~3。色素体盘状。多囊室着生于长同化丝细胞间, 由营养细胞形成, 长短不一, 长60~500 µm, 宽30~50 µm。单室囊不明。
习性和产地: 在中潮带大石沼中, 附着于海蒿子Sargassumconfusum体上, 生长于早春, 和短毛藻Elachistasp.混生。产于辽宁大连。中国特有种类。
模式标本产地: 中国大连( Dalian, China)。模式标本由栾日孝先生于1999年3月17日采自大连, 模式标本号为AST 9990070。

图2 细弱褐毛藻Halothrix gracilis Luan et Ding sp.nov. Fig. 2 Halothrix gracilis Luan et Ding sp.nov.
2.3 褐毛藻(图3)
Halothrixlumbricalis(Kuetzing) Reinke 1888[1]:19; Newton 1931[15]:132; Hamel 1931~ 1939[16]: 126; Okamura 1936[14]:147; Taylor 1957[17]: 143; Tseng et Cheng 1954[8]:110; Perestenko 1980[18]: 138; Tseng et al. 1983[9]:174; Scagel et al. 1986[19]:92; Luan 1989[10]:82; Yoshida et al. 1998[3]:255; Tseng et al. 2008[11]:329。
Ectocarpus lumbricalisKuetzing 1845[20]:233。
特征描述: 藻体黄褐色, 丛生呈束, 高0.5~3 cm。基部具有假根状藻丝, 交织成固着器。假根丝细胞长20~50 µm, 宽7~25 µm。直立部分由同化丝组成, 同化丝中部粗, 向两端渐细, 细胞宽而短; 下部细胞长7.5~12.5 µm, 宽25~32 µm; 中部细胞长18~30 µm, 宽55~85 µm; 上部细胞长12~25 µm, 宽20~50 µm。藻体大部分不分枝, 但近基部, 常有短分枝。色素体盘状。多室囊, 无柄, 密集围绕同化丝上部的一部分细胞上。
习性和产地: 生于潮间带, 多附着于大叶藻Zosteramarina(一种海草)和一种虾海藻属海草Phyllospadixsp.或其他藻体上, 多见于春季。产于辽宁大连、长海, 山东青岛、威海。
国外分布: 大西洋两岸的亚寒带及暖温带北部海区, 日本, 俄罗斯大彼得湾。
模式标本产地: 波罗的海(Baltic Sea)。
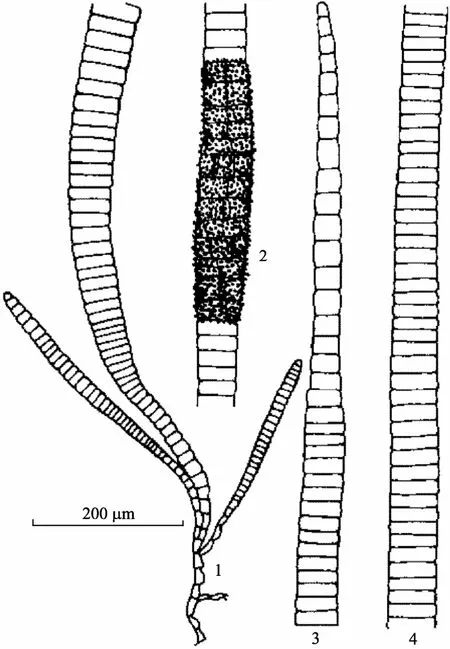
图3 褐毛藻Halothrix lumbricalis (Kuetzing) Reinke Fig. 3 Halothrix lumbricalis (Kuetzing) Reinke
2.4 垫状褐毛藻(图4)
Halothrix pulvinataLuan et Ding sp. nov.。
特征描述: 藻体黄褐色, 密集丛生, 高0.8~l cm。基部多由髓状的丝体组成厚薄不等的垫状部分。垫状部分为假膜构造, 通常较厚, 由髓状丝体组成。髓状丝体长1300~1700 µm, 细胞表面凹突不平。其下部细胞长70~170 µm, 宽9~25 µm, 长宽比为4~11; 上部细胞长40~165 µm, 宽20~35 µm, 长宽比为1.3~6.6。丝体基部生有少量的短假根丝。假根丝细胞长25~80 µm, 宽8~12 µm, 长宽比为3~6.7。同化丝着生于髓状丝体上部, 可分成长同化丝和侧丝(短同化丝)。长同化丝粗而长, 细胞横壁处稍缢缩; 下部细胞长11~20 µm, 宽40~50 µm, 长宽比为0.2~0.4; 中部细胞长25~80 µm, 宽55~70 µm, 长宽比为0.4~1.2; 上部细胞长25~160 µm, 宽15~40 µm, 长宽比为1.3~6。侧丝(短同化丝)着生于长同化丝的基侧, 不分枝或分枝, 较短, 长 200~1200 µm, 由16~40个细胞组成。侧丝下部细胞长12~13 µm, 宽12~20 µm, 长宽比为0.6~2.5; 中部细胞长15~40 µm, 宽20~40 µm, 长宽比为0.5~1; 上部细胞长8~25 µm, 宽10~22 µm, 长宽比为0.7~1.3。色素体球形。多室囊生于长同化丝间或顶部, 多连续, 生长。单室囊不明。
习性和产地: 在低潮线下附着于一种虾海藻Phyllospadixsp.叶面上, 和Protectocapusspeciosus混生, 在同化丝体上附有Acrohaetiumhumile。产于辽宁长海县海洋岛。
模式标本产地: 中国辽宁海洋岛( Haiyangdao, Liaoning, China)。模式标本由栾日孝先生于2003年3月 17日采自辽宁海洋岛, 模式标本号为AST039032A。
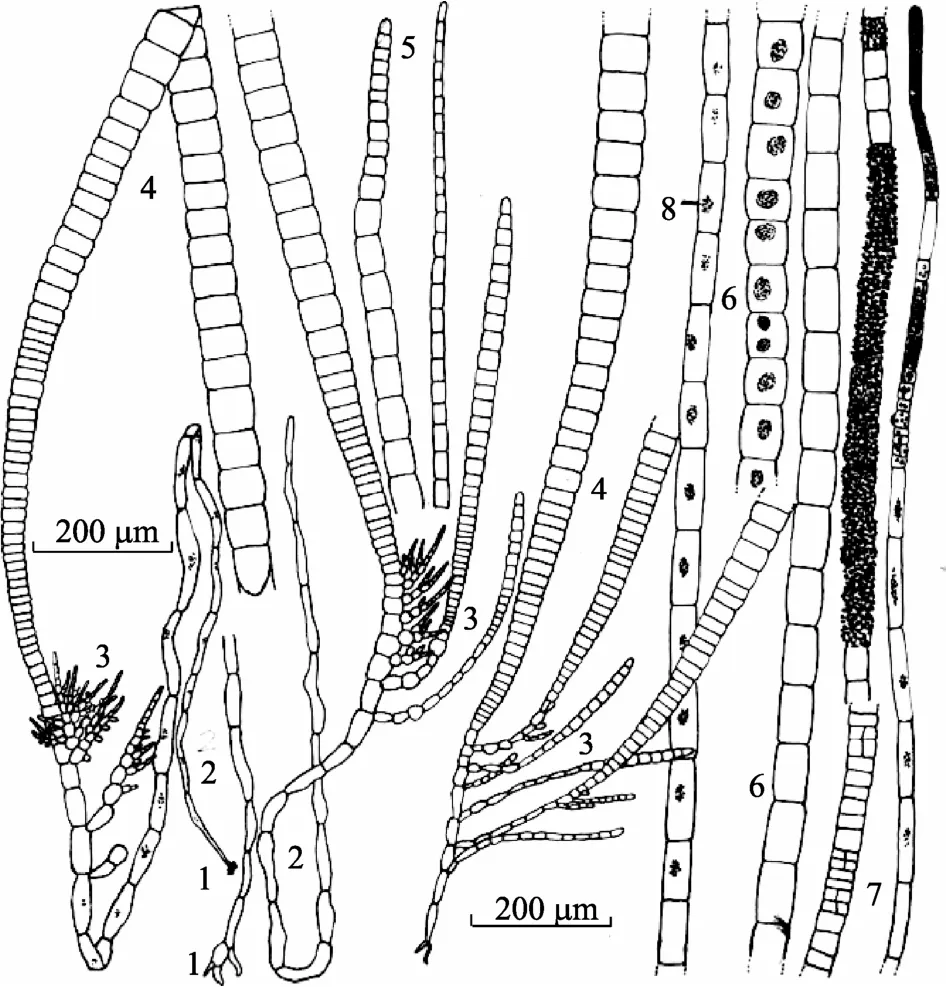
图4 垫状褐毛藻Halothrix pulvinata Luan et Ding Fig. 4 Halothrix pulvinata Luan et Ding
2.5 中国褐毛藻属Halothrix分种检索表

3 讨论
过去, 我国对海洋褐毛藻属的研究报道并不多, 仅报道了2个种。其中报道得较多的是褐毛藻H. lumbricalis(Kuetzing) Reinke[8-11], 而可疑褐毛藻H.ambiguaYamada仅报道一次[12]。本文的研究在此基础上, 发现了2个新物种, 即细弱褐毛藻H. gracilisLuan et Ding sp. nov.和垫状褐毛藻H. pulvinataLuan et Ding sp. nov.。
细弱褐毛藻H. gracilisLuan et Ding sp. nov.与扭曲褐毛藻H. tortuosaTakamatsu[3]在形态上很相似。其区别为前者长同化丝稍细, 直径15~55 µm, 中、上部细胞较长, 长宽比为1.3~3; 短同化丝的细胞较多, 10~17个, 具有匍匐丝体; 无单室囊; 附生在海蒿子Sargassum confusum的藻体上。而后者长同化丝稍粗, 直径20~55 µm, 中、上部细胞略短, 长宽比为约1~2; 短同化丝的细胞较少, 7~10个, 匍匐丝体不明; 具有单室囊, 附生在一种虾海藻属海草Phyllospadixsp.的植株上。
垫状褐毛藻H. pulvinataLuan et Ding sp. nov.主要特征是具有明显的垫部, 厚达1.3~1.7 mm, 由很多透明丝体组成, 丝体细胞13~20个, 细胞长70~170 µm, 宽9~25 µm, 长宽比为1.3~11; 长同化丝在基部多分枝等特征可与本属中其他种类相区别。
[1] Reinke J. Die braunen algen (Fucaceen und Phaeosporeen) der Kieler Bucht[J]. Berichte der Deutsche Botanischen Gesellschaft, 1888, 6: 14-20.
[2] Smith G M. Manual of phycology[M]. Waltham, Mass: Chronica Botanica Company, 1951.
[3] 吉田忠生. 新日本海藻誌 [M].東京:内田老鹤圃, 1998.
[4] Fletcher R L. Seaweeds of the British Isles, 3(1) [M]. London: British Museum, 1987.
[5] Womersley H B S. The marine benthic flora of Southern Australia II[M]. Adelaide:South Australian Government Printing Division,1987.
[6] Lee Y P. The circumscription of the genusHalothrixReinke (Chordariales, Phaeophyceae)[J]. Algae (The Korean Journal of Phycology), 2001, 16: 35-43.
[7] Skinner S. Australian and New Zealand species ofElachistaandHalothrix(Elachistaceae, Phaeophyta)[J]. Transactions of the Royal Society of South Australia, 1985, 109: 151-160.
[8] 曾呈奎, 郑柏林. 青岛海藻的研究I[J].植物学报, 1954, 3(1): 105-120.
[9] Tseng C K. Common Seaweeds of China[M]. Beijing: Science Press, 1983.
[10] 栾日孝. 大连沿海藻类实习指导[M]. 大连: 大连海运学院出版社, 1989.
[11] 曾呈奎. 黄渤海海藻[M]. 北京:科学出版社, 2008.
[12] 野田光藏(Noda). 中国东北区(满洲)の 植物誌 [M]. 东京: 風間書房, 1971.
[13] Yamada Y. Report on the biological survey of Mutsu Bay, 9. Marine algae of Mutsu Bay and adjacent waters. II[D]. [s.l.] Scientific Reports of the Tôhoku Imperial University, Biology, 1928, 3: 497-534.
[14] Okamura K. Nippon kaisô shi [Descriptions of Japanese algae] [M]. Tokyo: Uchida Rokakuho, 1936.
[15] Newton L. A handbook of the British seaweeds[M]. London: British Museum (Natural History), 1931.
[16] Hamel G. Phéophycées de France[M]. Paris: VII-XLVII, 1931-1939.
[17] Taylor W R. Marine algae of the northeastern coast of North America[M]. Ann Arbor: The University of Michigan Press, 1957.
[18] Perestenko L P. Vodorosli Zaliva Petra Velikogo [The seaweeds of Peter the Great Bay] [M]. Moscow: Akad. Nauk. SSSR. Bot. Inst., 1980.
[19] Scagel R F, Garbary D J, Golden L, et al. A synopsis of the benthic marine algae of British Columbia, northern Washington and southeast Alaska, Vol. 1 [M]. Vancouver: Department of Botany, University of British Columbia, 1986.
[20] Kützing F T. Phycologia germanica, d. i. Deutschlands Algen in bündigen Beschreibungen. Nebst einer Anleitung zum Untersuchen und Bestimmen dieser Gewächse für Anfänger[M]. Nordhausen: W. Köhne, 1845.
猜你喜欢
今日农业(2024年13期)2024-02-25 00:00:00
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
语文周报·教研版(2021年28期)2021-08-19 02:14:42
热带农业科学(2017年9期)2017-10-23 22:56:24
科技创新与应用(2017年20期)2017-07-15 20:43:02
数学理论与应用(2017年2期)2017-06-27 07:38:56
数学理论与应用(2017年2期)2017-06-27 07:38:54
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
西南农业学报(2016年4期)2016-05-17 05:41:45
数学理论与应用(2016年1期)2016-02-28 09:25:55
