
扁颅蝠和褐扁颅蝠体表寄生革螨的宿主选择
2013-12-17 09:16:50张光良唐占辉洪体玉谭梁静沈琪琦张礼标
Zoological Research 2013年1期
张光良,唐占辉,洪体玉,杨 剑,曾 玉,谭梁静,刘 奇,陈 毅,沈琪琦,张礼标,*
1. 广东省昆虫研究所,广州 510260;
2. 东北师范大学 环境与生态学院,长春 130024;
3. 海南医学院,海口 571199
包括翼手目在内的大多数哺乳动物是许多体表寄生虫的天然寄主。许多学者对体表寄生虫与宿主身体状况的关系进行了研究,并提出了两种不同的观点。一些研究表明寄生虫对宿主身体状况有明显影响(Brown & Brown, 1986, 2004; Bergströma et al,1999; Neuhaus, 2003; Whiteman & Parker, 2004),而另一些研究则认为宿主身体状况与其寄生虫感染的负荷量是相互独立的,两者之间无必然相关性(Perez-Orella & Schulte-Hostedde, 2005; Tompkins et al, 1996)。有研究发现,在长耳鼠耳蝠(Myotis bechsteinii)种群内,高健康指数(体重与前臂长的比值较高)个体携带更多的寄生虫,即寄生虫负担与宿主健康指数正相关(Reckardt & Kerth, 2009)。
体表寄生虫(包括体表寄生革螨)大多表现不同程度的宿主专一性(宿主特异性)。专一性高的寄生虫常有一个主要宿主,可能还有很少几个偶尔寄生的宿主(Tripet et al, 2002),即使是广寄生的寄生虫也通常表现出对某一或某一些宿主的偏爱选择(Tripet & Richner, 1997; Soler et al, 1999; Johnson et al, 2002)。广寄生种类的生境区域较其主要宿主的生境更为宽广(Tripet & Richner, 1997; Poulin,2007),而专一性寄生种类能有效利用宿主的微气候和生活史以减小竞争(Timms & Read, 1999; ter Hofstede & Fenton, 2005; Poulin, 2007)。由于生境隔离及寄生虫生活史策略,蝙蝠体表的寄生虫通常属于专一性较强的寄生种类(Dick et al, 2003; Dick,2007; Shatrov, 1992)。虽然对蝙蝠体表革螨类寄生虫的专一性存在争议(Bruyndonckx et al, 2010; Guiller& Deunff, 2010),但野外统计学分析及室内不同寄主物种的交叉感染研究均表明大多蝙蝠体表寄生革螨属于较强的寄主专一性种类,且常常表现一种寄生革螨只寄生一种蝙蝠(Dittmar et al, 2006;Seneviratne et al, 2009; Tian et al, 2009)。Bruyndonckx et al (2009)比较研究了不同群落的马耳他鼠耳蝠(Myotis punicu)及其体表鼠耳蝠革螨(Spinturnix myoti)的线粒体变异,发现革螨线粒体变异与其宿主线粒体和原子核变异紧密相连,认为这两个相互作用的物种在进化树上为伴同演化(co-differentiation)。
宿主专一性可能是一系列过程/作用的结果:宿主隔离阻碍寄生虫感染潜在的新宿主,寄生虫较弱的散布能力降低其分布到同域并存潜在宿主的可能性,且其生理、形态及行为上的适应将进一步减小其在新寄主体表寄生的机会(Dick & Patterson,2007; Reed & Hafner, 1997; Timms & Read, 1999)。实验室探究寄生虫寄生专一性的主要策略之一是给予寄生虫在不同潜在宿主间选择的机会(Esbérard et al, 2005; Giorgi et al, 2004)。寄生虫选择性感染能给予它们最高舒适的宿主或宿主种群,且在首选宿主体表存活的可能性更大(Giorgi et al, 2004)。
广西南宁地区共同栖宿在相同竹林内的扁颅蝠(Tylonycteris pachypus)和褐扁颅蝠(T. robustula)经常更换栖宿竹筒,它们的体表寄生虫革螨均表现寄主专一性,且不同性别、年龄和繁殖状态的宿主体外寄生革螨负荷量也具有明显差异(Zhang et al,2010)。本文通过野外调查研究了扁颅蝠和褐扁颅蝠体外寄生革螨负荷量与其宿主健康状况之间的关系,同时通过室内感染行为实验,探讨了体外寄生革螨对其原宿主性别以及非原宿主的选择偏好,以期进一步了解体表寄生革螨对于宿主选择的规律性。
1 材料与方法
1.1 研究地点
研究地点位于广西壮族自治区龙州县,地处南亚热带南沿,气候暖和湿润,常年无霜,年均温22.2 °C,平均海拔~200 m,喀斯特地貌显著。扁颅蝠和褐扁颅蝠均栖息在村庄及附近的刺竹(Bambusa stenostachya)竹筒内,可在不同时间轮流使用同一个栖息竹筒,但极少出现两者混居的现象(Zhang et al, 2004)。
1.2 蝙蝠及寄生革螨的收集
蝙蝠样本于2010年8—10月采自广西壮族自治区龙州县县城附近的村庄。捕捉两种扁颅蝠潜在栖宿竹筒内的全部蝙蝠个体,同一个栖宿竹筒的蝙蝠装在同一个布袋内带回临时实验室。前期调查发现,扁颅蝠体表寄生虫为拟雷氏巨刺螨(Macronyssus pararadovskyi),褐扁颅蝠体表寄生虫为雷氏巨刺螨(Macronyssus radovskyi)(Tian et al,2009)。用镊子和软刷子尽量取下蝙蝠体表的可见寄生虫(革螨类)放在培养皿中,并用于感染无体表寄生革螨蝙蝠个体。
1.3 寄生革螨离开寄主后的存活时长
从扁颅蝠体表选取 10只拟雷氏巨刺螨放入培养皿中,每10 min计数一次存活革螨个体数,直至所有个体死亡。重复7次,为确定感染实验的最短时长提供依据。由于无法从褐扁颅蝠体表获得足够的雷氏巨刺螨,我们用拟雷氏巨刺螨离开宿主后的存活时长作为雷氏巨刺螨感染实验最短时长的参考。
1.4 两种扁颅蝠的寄生革螨感染实验
1.4.1 种内感染
分别选取 7只寄生革螨(拟雷氏巨刺螨和雷氏巨刺螨)对清除过寄生虫的原宿主物种(扁颅蝠和褐扁颅蝠)雌、雄个体进行感染实验。将载有寄生革螨的培养皿置于20 cm×15 cm×10 cm的塑料盒子中央,待寄生革螨均爬行至培养皿边缘时将一雌一雄蝙蝠放入塑料盒,盖上培养皿盖及塑料盒盖。1 h后分别计数感染在雌、雄蝙蝠体表的寄生革螨数量,总量以最后计数的寄生革螨个体总数为准。将感染数据转换为百分比(Christe et al, 2007; Esbérard et al, 2005),即实验结束时雌、雄蝙蝠个体体表寄生革螨数量分别占寄生革螨总数的百分比。扁颅蝠重复32次,褐扁颅蝠重复30次。
1.4.2 种间交叉感染
通过室内行为实验探究寄生虫寄生特异性的主要策略是给予寄生虫在不同潜在寄主间选择的机会。由于在实验期间无法获得足够的雷氏巨刺螨,因此仅以拟雷氏巨刺螨感染扁颅蝠与褐扁颅蝠。选取7只拟雷氏巨刺螨对2只已经完全清除寄生革螨的蝙蝠(相同性别的成年扁颅蝠与褐扁颅蝠)进行感染,实验操作过程同上。1 h后分别计数 2只蝙蝠体表的寄生革螨数量,总量以最后计数的革螨个体数为准,并分别计算每只蝙蝠体表革螨数量的百分比(Esbérard et al, 2005; Christe et al, 2007),重复27次。
1.5 寄生革螨数量与寄主身体状况
虽然对于动物健康指数的计算尚无一致观点(Green, 2001; Schulte-Hostedde et al, 2005),但是我们以蝙蝠体重与前臂长的比值作为健康指数(Lewis,1996; Lučan, 2006; Laurenco & Palmeirim, 2007),并用于分析扁颅蝠和褐扁颅蝠身体状况与其体表寄生革螨数量的关系。由于具有相同革螨数量的重复个体较多(特别是无革螨感染个体所占比例较大,扁颅蝠:33.0%,褐扁颅蝠:44.1%),携带相同数量革螨蝙蝠个体的健康状况指数取平均数来表示。
1.6 数据统计与分析
记录每个蝙蝠栖宿群的个体数量及单个样本的物种、性别、体重、前臂长及体表寄生革螨数量等数据,并用SPSS17.0进行分析。用t-test来分析种间及种内感染差异。采用 Spearman相关分析检验不同性别个体健康指数与寄生革螨负荷量的相关性。
2 结 果
2.1 寄生革螨离开寄主后的存活时长
拟雷氏巨刺螨离开宿主扁颅蝠后不能很好存活。实验结果显示在温度24.7 °C,湿度72%条件下,拟雷氏巨刺螨离开寄主~70 min后开始死亡,100~120 min内死亡速率最高,140 min后,7次重复实验的寄生革螨均已没有存活个体(图1)。
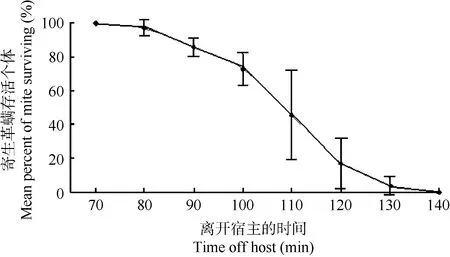
图1 拟雷氏巨刺螨离开宿主的存活时间Figure 1 Survival time of Macronyssus pararadovskyi off host
2.2 寄生革螨感染实验结果
种内感染行为实验显示2种体表寄生革螨均更倾向于感染原宿主雄性个体。扁颅蝠雌、雄个体感染率分别为(42±12)%及(58±12)%(t=-3.6, df=31,P<0.01),褐扁颅蝠雌、雄个体感染率分别为(37±11)%及(63±11)%)(t=-6.1, df=26, P<0.001)(图2A, B)。
拟雷氏巨刺螨种间感染实验显示,原宿主扁颅蝠个体被感染的寄生革螨数量(71±13)%显著高于褐扁颅蝠(29±13)%(t=9.1, df=29, P<0.001)(图 2C)。
2.3 寄生革螨负荷量与寄主健康状况
两种扁颅蝠的寄生革螨负荷量呈现较明显的非连续性波动,雌、雄扁颅蝠寄生革螨负荷量范围分别为0~47及0~32,雌、雄褐扁颅蝠则分别为0~12及 0~13。雌、雄扁颅蝠健康指数范围分别为 0.122~0.160及 0.123~0.153,而雌、雄雄性褐扁颅蝠则分别为为 0.161~0.193及 0.161~0.212。
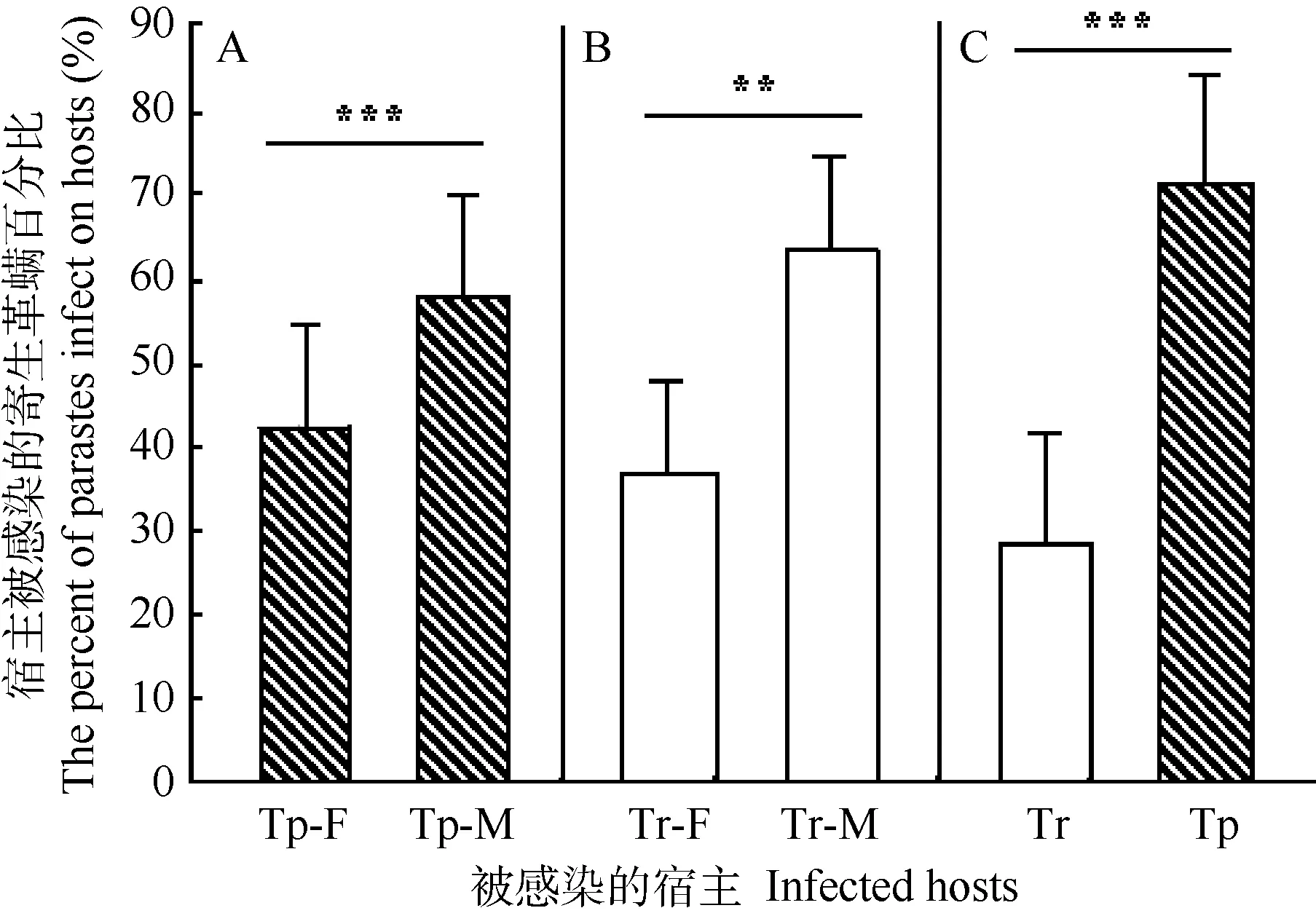
图2 扁颅蝠和褐扁颅蝠体表寄生革螨感染百分比Figure 2 Percentage of infected mites on T. pachypus andT. robustulaA:拟雷氏巨刺螨感染雌、雄原宿主扁颅蝠;B:雷氏巨刺螨感染雌、雄原宿主褐扁颅蝠;C:拟雷氏巨刺螨感染原宿主扁颅蝠和非原宿主褐扁颅蝠。Tp:扁颅蝠;Tr:褐扁颅蝠, F :雌性,M:雄性;**:P<0.01,***:P<0.001。A: Macronyssus pararadovskyi infected original host T. pachypus; B: M.radovskyi infected original host T. robustula; C: intercross infection between original host T. pachypus and novel host T. robustula with M. pararadovskyi.Tp: Tylonycteris pachypus; Tr: Tylonycteris robustula, F: Female, M: Male.**: P<0.01, ***: P<0.001.
Spearman相关检验显示,雄性扁颅蝠及雌、雄褐扁颅蝠的健康指数与寄生革螨负荷量没有显著相关性(P>0.05),而雌性扁颅蝠健康指数与体表寄生革螨数量存在一定的正相关(Spearman: rs=0.55,P<0.01,n=24)(图3B),即健康指数较大的雌性扁颅蝠个体携带较多革螨。
3 讨 论
3.1 寄生革螨的专一性及其对宿主性别的选择偏好
蝙蝠体表的螨类寄生虫不能长时间离开宿主且扩散能力较弱,许多种类的寄生虫终生寄生在同一个宿主体表;偶然因素导致其脱离宿主后,需要快速探测并选择潜在寄主,而原宿主物种通常更有利于其生存和繁衍(Thomas et al, 2000)。我们的存活实验表明,扁颅蝠体外的拟雷氏巨刺螨离开宿主被置于玻璃培养皿后,70 min开始出现死亡,2 h后则全部死亡。
寄生虫对原宿主感染行为实验表明,寄生虫感染趋势与野外自然条件下相似,即野外条件下高感染数量的类群(如雄性个体)在室内感染时也同样表现高感染量(Giorgi et al, 2004; Christe et al, 2007)。我们的感染实验表明,扁颅蝠和褐扁颅蝠的体外寄生革螨趋于选择雄性宿主,与野外自然条件下的结果(Zhang et al, 2010)一致,说明与雌性个体相比,这两种扁颅蝠的雄性个体均为其体外寄生革螨更适合的宿主。
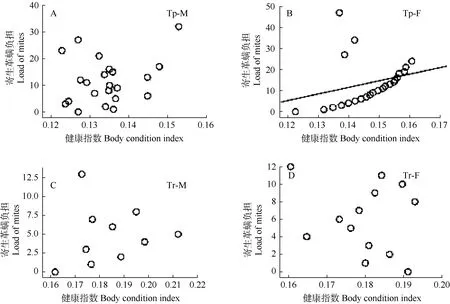
图3 扁颅蝠和褐扁颅蝠体表寄生革螨数量与健康指数的关系Figure 3 Relationship between mite load and body-condition index of Tylonycteris pachypus (Tp) and Tylonycterisrobustula (Tr) F: Female, M: Male
蝙蝠体表寄生虫的专一性说明其与宿主之间进化过程的同步性(Bruyndonckx et al, 2010)。然而大多关于寄生虫特异性寄生的观点都是通过大量的区系统计得到的结论,关于其机制尚存在较大分歧。目前的主要观点有两个,第一是寄生虫的转移能力决定其专一性,第二个是由于寄生虫在新的宿主体表不能很好存活,从而使得寄生虫表现出很强的专一性。蝙蝠体表的寄生虫螨类不能长时间离开宿主,且扩散能力较弱,导致许多种类的寄生螨终生寄生在同一宿主体表。而当寄生虫由于偶然因素脱离了宿主时,常能根据潜在宿主细微的化学信息差别选择有利宿主。革螨类能够利用蝙蝠的振动、阴影及 CO2浓度等因素辨别有利的潜在寄主(Sonenshine, 1993; Cox et al, 1998; Osterkamp et al,1999; Devine et al, 2000)。我们设计的交叉感染实验给予寄生革螨选择两种潜在宿主的机会以检验寄生革螨再次感染宿主的选择趋势,虽然扁颅蝠与褐扁颅蝠亲缘关系较近,且常常轮流使用同一个栖宿竹筒,但野外观察发现这两种蝙蝠极少同时共用同一个栖宿竹筒(Zhang et al, 2004),拟雷氏巨刺螨与雷氏巨刺螨出现宿主间转换的机会极少;即使是偶然脱落掉在蝙蝠栖宿竹筒内的革螨也无法幸存并进驻新宿主。实验结果显示,扁颅蝠体表拟雷氏巨刺螨显著趋于选择其原宿主,且对潜在宿主褐扁颅蝠则不表现明显选择喜好,这可能与其专一性寄生有关。寄生虫选择一个新的寄主物种可能导致其快速死亡,而选择原宿主则更有利于其生存(Giorgi et al, 2004)。两种扁颅蝠虽然生活在同一区域,但它们的栖宿群却只由同一物种的个体组成,这种栖宿方式使扁颅蝠体外革螨类寄生虫失去了在潜在宿主物种间转移的可能。我们认为,扁颅蝠体外寄生革螨类的特异性寄生特性是长期进化的结果。褐扁颅蝠体外寄生革螨的专一性有待进一步验证。
3.2 身体健康状况与寄生革螨负荷量的关系
蝙蝠体表寄生虫负荷量有明显季节及性别差异(Christe et al, 2000; Zhang et al, 2010)。蝙蝠个体在食物短缺的季节常表现体重下降。一些研究发现在寄生虫爆发期,身体状态较差的个体常寄生相对较大数量的寄生虫。而在一定时期,寄生虫负荷量又与健康指数表现正相关性。由此可见,寄主健康状态与寄生虫数量之间的关系可能是多种多样的(Møller, 2000; Weddle, 2000; Zahn &Rupp, 2004; Lučan, 2006; Laurenco & Palmeirim,2007)。
本研究检验了成年扁颅蝠和褐扁颅蝠健康指数与体表寄生革螨负荷量的关系,发现在雌性扁颅蝠个体寄生革螨负荷量与健康指数存在一定正相关性,与Reckardt & Kerth (2009)在长耳鼠耳蝠种群发现的趋势一致,可能是由于较好健康条件更受到体表寄生虫的青睐(Christe et al, 2003)。但是,雄性扁颅蝠和雌、雄褐扁颅蝠的体外寄生革螨负荷量与宿主健康指数无相关性。扁颅蝠和褐扁颅蝠的体外寄生革螨负荷量相对较低(Zhang et al., 2010),宿主选择策略可以解释扁颅蝠和褐扁颅蝠体外寄生革螨负荷量与其宿主健康指数无相关性或正相关性。负荷量较低时,寄生革螨通常不会对其寄主带来直接伤害,寄主的防御投入亦通常不会随寄生革螨的增加而增加,寄生革螨通常会选择有利于其生存繁衍的宿主个体,即身体状态较好,健康指数较高的个体(Giorgi et al, 2001)。
致谢:感谢韦力、张伟、叶建平和朱光剑在野外调查中给予的帮助。
猜你喜欢
中成药(2022年4期)2022-06-14 08:23:42
四川蚕业(2022年1期)2022-06-06 02:04:02
大科技·百科新说(2022年4期)2022-05-30 10:48:04
林业科学研究(2021年3期)2021-07-11 03:33:46
紫禁城(2019年2期)2019-04-28 12:10:06
中成药(2018年11期)2018-11-24 02:57:38
实验与检验医学(2017年1期)2017-03-13 03:46:31
中国卫生标准管理(2015年1期)2016-01-14 03:41:21
中国医疗美容(2015年2期)2015-07-19 10:11:59
安徽医药(2014年4期)2014-03-20 13:13:25

- Zoological Research的其它文章
- Blockage of glucocorticoid receptors during memory acquisition,retrieval and reconsolidation prevents the expression of morphineinduced conditioned place preferences in mice
- Seasonal variations in the energy budget of Elliot’s pheasant(Syrmaticus ellioti) in cage
- Seed caching and cache pilferage by three rodent species in a temperate forest in the Xiaoxinganling Mountains
- Proximity association in polygynous western black crested gibbons(Nomascus concolor jingdongensis): network structure and seasonality
- 中国鸟类亚种新记录——黑冠黄鹎
- 虹鳟LECT2的酵母表达、纯化及生物活性分析