
氮磷对青岛大扁藻和牟氏角毛藻单养和共培养生长的影响
2013-12-03 07:23徐善良
生物学杂志 2013年1期
邵 波,徐善良,2,齐 闯,邹 秀
(1.宁波大学海洋学院,宁波315211;2.宁波大学应用海洋生物技术教育部重点实验室,宁波315211)
微藻是养殖水体生态系统的重要组成部分,其种群生长、演替对养殖水体的生态质量具有重要的影响。研究表明,在复杂的自然水域中,微藻种群的生长除了受到生物因素的影响,还受到温度[1-3]、盐度[4-5]、光照[2,4,6-7]等诸多非生物因素的影响,但是研究者们普遍认为微藻的生长繁殖以及种间竞争更是受限氮磷营养盐条件不仅是决定微藻生长繁殖的主要因子,而且是关系微藻种间竞争的关键因素之一[8-9]。扁藻和硅藻是海洋环境中生理生态特性有显著差异的类群,是养殖水体中重要的饵料微藻,在贝类苗种生产上具有广泛的应用。为此,作者选取青岛大扁藻(Platymonas helgolandica var.tsingtaoensis)和牟氏角毛藻(Chaetoceros muelleri Lemmerman)为对象,进行单培养和共培养的比较实验,研究两种藻在共培养模式下,对不同氮磷质量浓度的响应,以了解氮磷对两类藻的调节模式。
1 材料方法
1.1 藻种
青岛大扁藻和牟氏角毛藻由宁波大学海洋生物工程重点实验室种质库提供。
1.2 微藻培养
实验用海水经脱脂棉过滤后,煮沸消毒,盐度27,pH值8.2。培养温度20℃,光照强度3000 lx,光暗周期 L :D=14 h:10 h,培养时间14 d。
1.3 氮磷营养盐起始添加量设置
分别以KNO3和KH2PO4作为氮源和磷源进行单因素实验,每个因素设置5个水平,具体质量浓度值见表1。以下以 N1、N2、N3、N4、N5、P1、P2、P3、P4、P5 代表各种处理。

表1 N、P初始添加浓度值(mg/L)Table 1 The initial concentrations of nitrogen and phosphorus(mg/L)
1.4 单培养实验
取指数生长期微藻进行接种培养,青岛大扁藻和牟氏角毛藻的接种密度分别为3.0×104cell/mL、12.0×104cell/mL。按照NMB3#培养液配方添加除N(或P)外的元素,用微量移液枪添加N(或P)达到每个水平要求的浓度,每个水平重复3次。
1.5 共培养实验
两种藻的起始接种密度以及营养盐的添加方案同单培养实验。每个水平也都设置3个重复。
NMB3#培养液的母液配方:KNO3100 g;KH2P0410 g;MnSO40.25 g;FeSO4·7H2O 2.5 g;Na2EDTA 20 g;VB120.5 μg;VB15 μg,溶解于 1000 mL 蒸馏水。以 1 :1000海水比例使用。
1.6 细胞计数
隔天取1 mL藻液,用Lugol氏液固定,血球计数板计数。每个样品计数3次,取其平均值。
1.7 数据处理
统计分析在EXCEL2010和spss 17.0中进行。
2 结果
2.1 青岛大扁藻和牟氏角毛藻在不同氮磷处理下的生长特征
青岛大扁藻和牟氏角毛藻在不同氮磷处理下的生长曲线见图1。图中可见,单培养模式下,不同的氮质量浓度处理直接导致青岛大扁藻生长状况有所差异。比较发现,随着氮质量浓度的提高,青岛大扁藻进入静止期的时间相应地延后,生长繁殖所能达到的最大细胞密度相应地增大。在单培养模式下,不同磷质量浓度处理以及共培养模式下,青岛大扁藻表现出相同的规律。至于牟氏角毛藻,在两种培养模式下其最大藻密度都呈现单峰型,即最大藻密度先随着营养盐质量浓度的升高而变大,随后到达峰值,之后与营养盐浓度呈负相关。同时也可发现,低氮或者低磷处理下,在培养后期牟氏角毛藻的细胞密度逐渐减小;在高氮组也发生同样的现象,而此时,青岛大扁藻历经阶梯式增长后,呈现快速增殖的趋势。经配对样本t检验分析,结果显示,在单培养和共培养模式下,两种藻的生长存在显著差异(P<0.05)。

图1 青岛大扁藻和牟氏角毛藻在不同氮磷处理下的生长曲线Fig 1 The growths curve of Platymonas helgolandica var.tsingtaoensis and Chaetoceros muelleri Lemmerman in different nitrogenous and phosphorus concentration
2.2 不同氮磷处理对最大藻密度的影响
由图2、3知,共培养模式下,不同氮磷处理对青岛大扁藻和牟氏角毛藻的最大藻密度有显著差异(P<0.05)。在不同氮处理下,青岛大扁藻在低氮组的最大细胞密度明显小于中氮和高氮组,并且在KNO3浓度为110 mg/L时取得极大值,但是与KNO3浓度为70 mg/L、90 mg/L组没有显著差异;而牟氏角毛藻在低氮和高氮组的最大藻密度相对较低,在KNO3浓度为70 mg/L时取得最大值,与低氮和高氮组存在高度显著差异(P<0.01)。
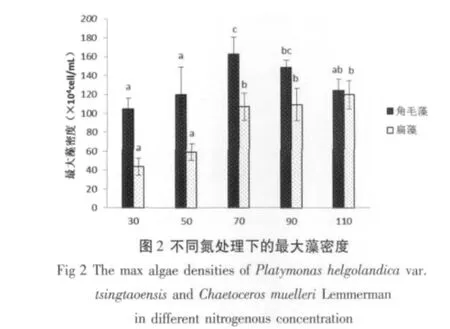

在不同磷处理下,青岛大扁藻也在高浓度组取得最大值,在低浓度时最大藻密度相对较低。牟氏角毛藻在KH2PO4浓度是5 mg/L取得最大值,与低磷处理存在显著差异(P<0.05),与高磷处理相近(P>0.05)。
2.3 不同氮磷比(N/P)对两种藻生长的影响
不同氮磷比对共培养时两种藻终细胞密度的影响见表2。由表可见,不同的氮磷比对两种藻终细胞密度都有显著影响(P<0.05)。当N/P<9.43,青岛大扁藻终细胞密度随N/P值增大处于快速增加的状况;当N/P值在9.43~14.81时,青岛大扁藻的终细胞密度随N/P值增大而平缓增加,并在N/P值是14.81时取得最大值;当N/P>14.81,青岛大扁藻的终细胞密度随N/P值增大呈现减少的趋势。而牟氏角毛藻只有在N/P<9.43和N/P>26.93时,其终细胞密度的变化表现出与青岛大扁藻相似的规律;当N/P值在9.43~14.81范围时,牟氏角毛藻的变化趋势与青岛大扁藻截然相反,在N/P值为14.81时,取得较小值;当14.81<N/P<26.93,牟氏角毛藻的终细胞密度又出现一波随N/P值增大而快速增加的现象。同时还可以发现,无论低氮高磷还是高氮低磷都不利于青岛大扁藻和牟氏角毛藻获得较大的终细胞密度。
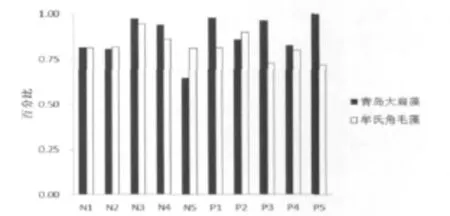
图4 共培养时两种藻的细胞数与单培养时的百分比Fig 4 The ratio of each last algae densities in mixed culture when compared with that cultured alone

表2 不同氮磷比下两种藻的终细胞密度Table 2 The last algae densities of Platymonas helgolandica var.tsingtaoensis and Chaetoceros muelleri Lemmerman in different N/P
2.4 两种藻的相互作用
以不同处理为X变量,共培养时两种藻的终细胞密度占各自单培养时终细胞密度的比值为Y变量绘图。由图4可见,无论何种处理,青岛大扁藻和牟氏角毛藻都受到对方不同程度的抑制。在N5处理下,结合图1,共培养的青岛大扁藻此时正处于快速增长期,而且藻密度高于其他处理,但是其终细胞密度仅为单培养时的64.30%,为其他各处理中最低,受到牟氏角毛藻最为强烈的抑制,是牟氏角毛藻对青岛大扁藻抑制作用的0.79倍。在N1、N2处理下,共培养的青岛大扁藻和牟氏角毛藻对对方的抑制能力相当。在N3处理下,青岛大扁藻受到牟氏角毛藻微弱的抑制作用,也可见青岛大扁藻对牟氏角毛藻的抑制作用也较微弱。在P5处理下,青岛大扁藻的终细胞密度与单培养时相当,而牟氏角毛藻的百分比最低,是71.64%。
3 讨论
3.1 营养盐浓度与微藻生长的关系
有学者提出海洋中浮游植物所需氮、磷营养盐下限分别为无机氮 0.08 mg/L,无机磷 0.018 mg/L[10]。本实验氮磷的最低值分别是4.16 mg/L、0.23 mg/L,高于所需下限,所以即使在低氮或者低磷处理下两种藻也表现出一定的生长性,但是也可以轻易地发现,低氮或者低磷处理下,两种藻都较早地进入静止期,最大藻密度也较低,这可能是因为低氮或者低磷组营养盐更快到达所需下限,最终形成营养盐限制,影响微藻的生长;在低氮或者低磷处理下,两种藻互相之间的抑制作用不及高氮或者高磷组,因此初步认为在低氮或低磷处理下,影响微藻生长繁殖的最直接因素是营养盐缺乏,而在高氮或者高磷处理下,培养后期影响微藻生长最直接要素是两种藻的互相抑制作用。研究发现,青岛大扁藻能分泌克生物质[11],在后期可能更能显示出竞争优势性。同时发现,当营养盐浓度达到一定值后,不同处理获得的最大细胞密度(或者终细胞密度)非常接近。单培养模式下,两种藻获得最大细胞密度的营养条件并不相同。陆茸[12]也发现当营养盐浓度达到一定浓度时,再增加营养盐不能明显地促进微藻的生长,营养盐的浓度越高并不一定有利于藻类生长[13],过高反而有抑制作用[13-14]。此外,两种藻获得最大藻密度的营养盐条件并不相同。Christian[15]认为微藻只有在特定的合适的营养盐范围才能大量繁殖,因为不同的微藻对最适氮磷营养盐的浓度有所不同。
3.2 N/P值对微藻生长的影响
微藻的生长不仅与氮磷营养盐浓度有关,而且当水体中氮磷营养盐达到一定水平,受到N/P值的影响。N/P过高或过低均不利于微藻的生长,因为每种微藻对氮磷结构比具有特殊的需求。本研究显示,在低N/P条件下,青岛大扁藻的生长比高N/P的好,与孙育平等[16]的研究相似,这可能是因为青岛大扁藻对磷有较高的要求[17],当N/P>16,磷易成为限制因子。而牟氏角毛藻在较高的N/P时具有更好的适应性,与梁伟峰等[14]的研究较为相近。
3.3 共培养模式对两种藻的影响
氮磷营养盐是影响微藻生长的重要因子之一,依照Monod方程,随着可利用氮磷营养盐的减少,对氮磷亲和性高的微藻容易成为优势种,而另一种的数量不会增加,两者发生竞争排斥[18]。研究显示,扁藻竞争能力明显强于眼点拟微绿球藻(Nannochloropsis ocutala)[19];青岛大扁藻会强烈地抑制强壮前沟藻(Amphidinium carterae Hulburt)的生长[11],因此在共培养的条件下,最大藻密度大多低于或接近于单种培养时其中一种微藻的细胞密度[11,20-21],但也有明显高于单种培养时最大细胞密度的现象[21-22]。本研究结果显示,实验期间共培养时青岛大扁藻和牟氏角毛藻的最大藻密度普遍低于单培养时的最大藻密度,与陈洁等[19]有类似的规律,这可能是因为,随着氮磷营养盐的消耗,促使青岛大扁藻和牟氏角毛藻争夺有限氮磷营养盐资源,各自的生长都受到对方的抑制。
[1]李 锋,葛长字,方建光,等.不同温度和接种密度下亚心形扁藻增殖的初步研究[J].海洋水产研究,2007,28(6):61-66.
[2]杨东方,陈生涛,胡 均,等.光照、水温和营养盐对浮游植物生长重要影响大小的顺序[J].海洋环境科学,20007,26(3):201-207.
[3]周洪琪,Renaud S M,Parry D L,等.温度对新月菱形藻、铲状菱形藻和杷夫藻的生长、总脂肪含量以及脂肪酸组成的影响[J].水产学报,1996,20(3):235-240.
[4]钱振明,邢荣莲,汤 宁,等.光照和盐度对8种底栖硅藻生长及其生理生化成分的影响[J].烟台大学学报(自然科学与工程版),2008,21(1):46-52.
[5]谢立民,林小涛,许忠能,等.不同类型虾池的理化因子及浮游植物群落的调查[J]. 生态科学,2002,22(1):34-37.
[6]石 娟,潘克厚.不同光照条件对小新月菱形藻和等鞭金藻8701生长及生化成分的影响[J].中国水产科学,2004,11(2):121-128.
[7]王爱军,王修林,王江涛,等.光照对东海赤潮高发区春季硅藻生长的影响[J]. 中国海洋大学学报,2006,36:173-178.
[8]Pratt C M.Competition between Skeletonema costatum and Olsthodiscus luteus in Narraaganesett Bay and in culture[J].Limnol Oceanogr,1966,11:447.
[9]朱从举,齐雨藻,郭昌弼.铁、氮、磷、维生素B1和B12对海洋原甲藻的生长效应[J]. 海洋与湖沼,1994,25(2):166-172.
[10]孙 耀,李 峰,李 健,等.虾塘水体浮游植物群落特征及其与营养状况的关系[J].海洋水产研究,1998,19(2):45-51.
[11]郝雯瑾,王 悠,唐学玺.两种海洋微藻——强壮前沟藻与青岛大扁藻之间的相互作用研究[J].中山大学学报(自然科学版),2008,47:98-105.
[12]陆 茸.东海围隔实验中营养盐对浮游植物生长的影响及其动力学研究[D].中国海洋大学,2005.
[13]牛明改.水体富营养化藻类资源竞争与种群演替规律的初探[D].苏州大学,2003.
[14]梁伟峰,李卓佳,陈素文,等.微藻群落在氮、磷比率与硅酸盐含量的生态位研究[J].农业环境科学学报2008,27(2):773-777.
[15]Christain E W.Planktonic bloom-forming Cyanobacteria and the eutrophication of Lakes and rivers,Freshwater Bio1ogy,1988,20:279-287.
[16]孙育平,赵曰水.不同氮、磷浓度下亚心形扁藻的生长及水中氮、磷变化[J]. 水产科学,2011,30(4):197-201.
[17]Tilman D,Sterner R W.Invasions of equilibria:tests of resource competition using two species of algae.Oecologia,1984,61:197-200.
[18]崔启武,刘家冈.生物种群增长的营养动力学[M].北京:科学出版社,1991.
[19]陈 洁,段舜山,李爱芬,等.眼点拟微绿球藻与扁藻在不同接种比例下的竞争[J]. 海洋科学,2003,27(5):73-76.
[20]谢志浩,肖 慧,蔡恒江,等.不同起始数量对赤潮异弯藻和东海原甲藻种间竞争的影响[J].海洋环境科学,2008,27(5):462-465.
[21]蔡恒江,唐学玺,张培玉,等.不同起始密度对3种赤潮微藻种间竞争的影响[J]. 生态学报,2005,25(6):1331-1335.
[22]周成旭,马 斌,汪飞雄,等.海洋原甲藻与三角褐指藻混合培养条件下的种群生长与氮磷营养盐变化[J].海洋科学,2006,30(12):58-61.
猜你喜欢
古今农业(2022年1期)2022-05-05
宁夏医学杂志(2020年3期)2021-01-21
天然产物研究与开发(2018年4期)2018-05-07
水利技术监督(2017年6期)2017-12-19
环境保护与循环经济(2017年3期)2017-09-26
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
中国医疗美容(2015年5期)2015-02-03
食品工业科技(2014年23期)2014-03-11
湖泊科学(2014年2期)2014-02-27
