
攀西黑山羊群体的AFLP遗传多样性研究
2013-12-01 06:23:52韦雷飞钟金城武志娟姜雪鸥张利亚
家畜生态学报 2013年2期
李 娜,杨 鹰,万 洁,韦雷飞,钟金城*,武志娟,姜雪鸥,张利亚
(1.动物遗传育种学国家民委-教育部重点实验室,西南民族大学,四川 成都 610041;2攀枝花市农林科学研究院,四川 攀枝花 617061)
扩增片段长度多态性(Amplified Fragment Length Polymorphism,AFLP)标记技术是由荷兰科学家Zabeau和Vos在PCR和RFLP技术基础上发展起来的一种DNA多态检测方法[1]。AFLP具有多态性丰富,可靠性好,分辨率高,方便快速等优点,已广泛应用于植物(小麦、水稻、拟南芥等)遗传育种研究中,在动物和微生物遗传育种研究中也有一些应用。Kim[2]等利用AFLP标记和表型分类法对海胆属的动物进行分类,发现海胆属内亲缘关系较近,种群间遗传距离为种内的3倍。Jeremy等[3]结合微卫星和AFLP标记构建5种罗非鱼-Oreochromis niloticus [wild type(on)和 red (Ron) strains]、O. aureus(Oa)、O. msambicus(Om)、Sarotherod on galilaeus(Sg)合成群体的遗传图谱,此图谱在培育罗非鱼时跟踪耐寒、耐盐及鱼体质量等数量性状位点的变动,在分子水平标记起关键作用。因而AFLP在构建物种分子遗传图谱,种质资源鉴定,遗传多样性研究等方面意义显著。
我国黑山羊主要分布在海拔2 500 m以下、气候温暖、雨水充分的地区。黑山羊产肉性能好,繁殖率高,生长发育快,养殖前景优良。四川省攀枝花市及周边地区黑山羊品种丰富,之前的研究主要以生产性能方面为主,很少在DNA分子水平上展开。本研究采用AFLP分子标记技术分析该地区及周边黑山羊品种(群体)的遗传多样性,以期为该地区黑山羊品种的进一步选育、保种提供理论依据。
1 材料与方法
1.1 样本采集
本试验选取攀枝花市的米易(MY,n=15)、盐边(YB,n=15)、龙潭(LT,n=11)、中坝(ZB,n=4),以及周边地区的云岭(YL,n=40)、乐至(LZ,n=20)、金堂(JT,n=20)、会理(HL,n=40)、建昌(JT,n=20)等5个黑山羊品种(类群),共185个健康个体(表1)。颈静脉采血,抗凝处理,带回实验室备用。
1.2 DNA提取
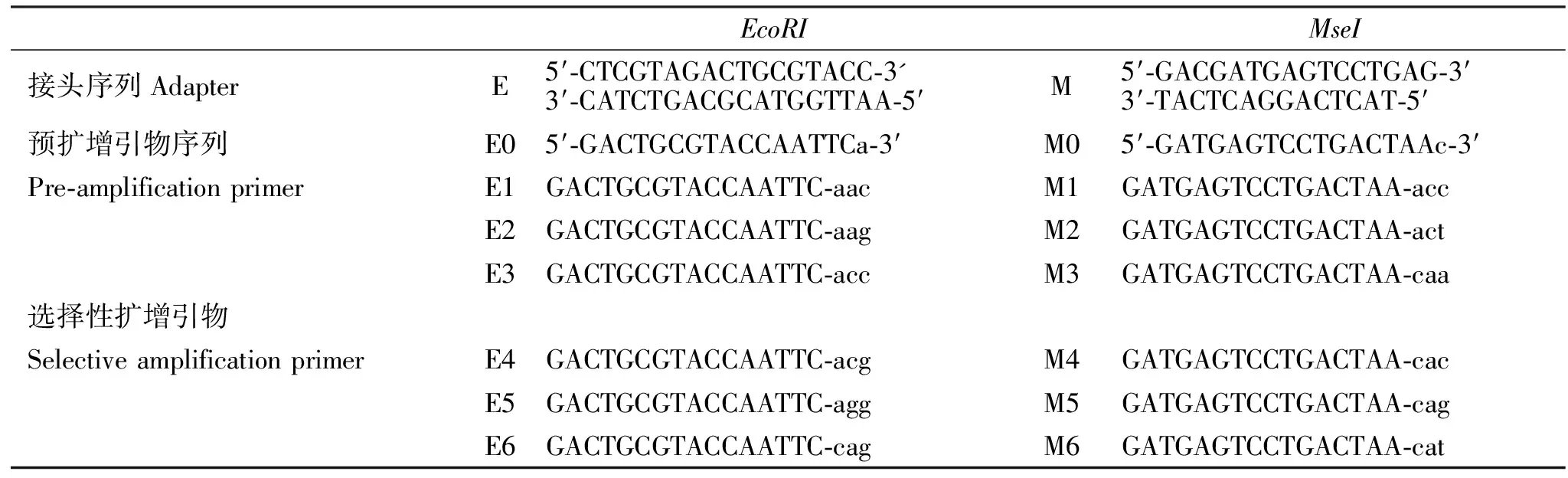
表1 接头、预扩增与选择性扩增引物序列Table 1 Adapters and primer sequences used for pre-amplification and selective amplification
基因组DNA提取参照《分子克隆指南》(第三版)(上册)哺乳动物DNA的快速分离法[4],所提取DNA置于-20 ℃冰箱保存。
1.3 AFLP分析
AFLP方法参照相关文献[5-9],并作少许修改。
1.3.1 酶切 0.5 μLMseI、0.5 μLEcoRI、4 μL 10Х酶切缓冲液、10 μL基因组DNA、加去离子水至40 μL,37 ℃酶切3 h,70 ℃灭活15 min。
1.3.2 连接 取酶切产物10 μL、1 μLMseI接头、1 μLEcoRI接头、1 μL T4-连接酶、2 μL buffer、5 μL ddH2O,16 ℃过夜,70 ℃灭活10 min,将连接产物置于-20 ℃冰箱,备用。连接接头为E,M(表1)。
1.3.3 预扩增 将1 μL连接产物、1 μL Primer Ep、0.1 μL Primer Mp、12.5 μL mastermix、6.4 μL ddH2O混合离心后, 进行PCR反应(94 ℃预变性 3 min, 94 ℃变性45 s, 56 ℃退火60 s, 72 ℃延伸80 s, 共 30~35个循环, 最后延伸10 min,4 ℃保存),用0.5 %琼脂糖电泳观察结果。预扩增产物稀释30倍,置于-20 ℃温度下,备用。须扩增引物为E0、M0(表1)。
1.3.4 选择性扩增 将1 μL稀释后预扩增产物、1 μL Primer E、0.16 μL Primer、12.5 μL M mastermix、10.34 μL ddH2O混合离心后, 进行PCR反应(94 ℃变性30 s, 56 ℃退火60 s, 72 ℃延伸60 s, 共15~20个循环,每个循环温度降低0.7 ℃,然后再90 ℃变性,56 ℃退火30 s,72 ℃延伸60 s,持续23个循环,72 ℃延伸7 min)。选择性扩增引物为E1~E6,M1~M6(表1)。
1.4 聚丙烯酰胺凝胶电泳检测扩增结果
预扩增产物用5%聚丙烯酰胺凝胶电泳检测,每个泳道点样2 μL,200 V预电泳5 min,85 V电泳1 h,0.2%硝酸银溶液染色,甲醛氢氧化钠显色液显色,凝胶成像系统拍照记录。
1.5 统计分析
1.5.1 条带记录 根据各样本AFLP标记结果,选取清晰且重复性好的条带,清晰可重复的记为“1”,否则记为“0”。
1.5.2 多态性与遗传多样性指数计算 根据所记录的 “0,1”矩阵,利用POPGENE和DCFA软件统计群体多态标记数、多态频率遗传多样性指数Shannon。
1.5.3 遗传距离的计算 根据条带的“0,1”矩阵,利用POPGENE和DCFA软件统计遗传距离。
1.5.4 聚类分析 根据遗传距离,利用Mega5.0软件,采用算术平均的非加权组对法(Unweighted Paie-Group Method with Arithmetic Averaging,UPGMA)和邻接法(Neighbor-Joining Method, NJ)构建群体系统进化树[10]。
2 结果与分析
2.1 AFLP扩增结果
15对AFLP引物扩增结果表明:多态频率范围为93.33%~100%,平均值99.01%,共扩增出1 011个条带,其中多态条带1 003个,不同引物组合扩增条带不同(表2、图1)。
引物组合E2-M1、E2-M3、E2-M4、E5-M3、E3-M2、E5-M1扩增条带较多,平均扩增条带数分别为41、38、38、30、30、29,AFLP多态性丰富。攀枝花本地黑山羊扩增多态条带最多,多态位点频率为85.42%,建昌黑山羊次之,乐至黑山羊最少,多态位点频率为25.99%,5个群体平均多态频率为42.54%(表3),表明这些黑山羊群体具有较丰富的遗传多样性。
2.2 遗传多样性指数
M.100 bp DNA Marker,1-9.依次表示选择性引物组合E2-M3、E2-M4、E2-M5、E2-M6、E3-M4、E3-M5、E3-M6、E4-M1、E5-M1对攀枝花龙潭2号个体DNA限制性片段扩增的电泳图谱
M.100 bp DNA Marker,1-9 stand for AFLP electrophorsis bands of restrict fragment of black goat LT2 by selective primer combinations E2-M3、E2-M4、E2-M5、E2-M6、E3-M4、E3-M5、E3-M6、E4-M1、E5-M1 correspondingly.
5个黑山羊群体的遗传多样性指数变异范围为0.0286~0.2131,表明这些黑山羊群体具有较丰富的遗传多样性(表4)。平均遗传多样性指数依次为建昌黑山羊(0.1346)>攀枝花黑山羊(0.1329)>金堂黑山羊(0.1230)>云岭黑山羊(0.1201)>乐至黑山羊(0.1101),说明建昌黑山羊遗传多样性最高,攀枝花黑山羊次之,乐至黑山羊遗传多样性最低。
2.3 遗传距离
5个群体中,群体间遗传距离范围为0.0062~0.0281,攀枝花与建昌之间的遗传距离最小(DR=0.0062),云岭与攀枝花间的遗传距离次之(DR=0.0128),乐至与金堂间遗传距离最大(DR=0.0281),金堂与其它4个类群遗传距离较大(DR=0.0173-0.0281)(表5)。说明攀枝花与建昌的亲缘关系最近,云岭与攀枝花亲缘关系较近,乐至与金堂的亲缘关系最远,金堂与其它4个类群遗传距离均比较远。
2.4 聚类分析
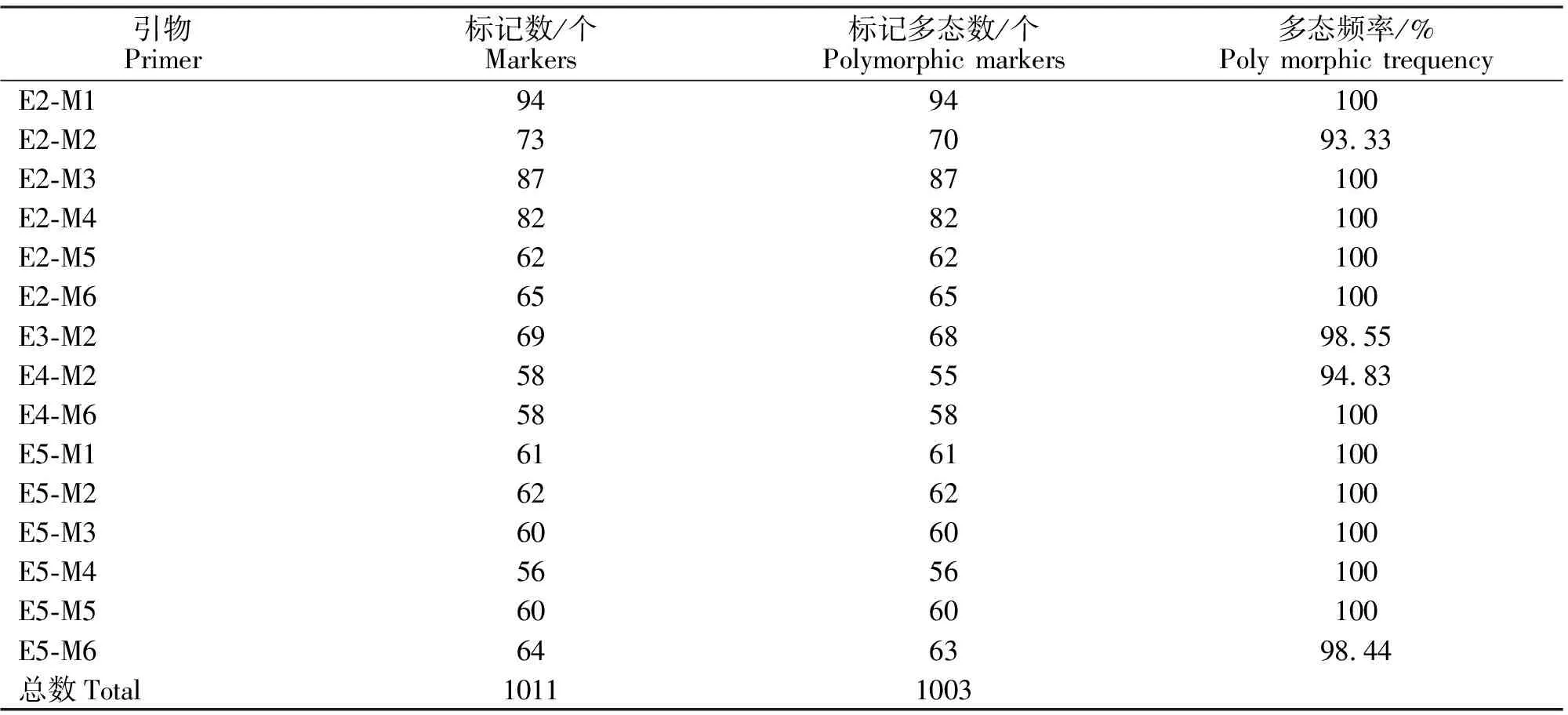
表2 选择性引物及5个群体的AFLP扩增结果Table 2 The selective primers and the result of AFLP among 5 black goat populations
根据遗传距离,采用UPGMA法和NJ法构建遗传系统树状图, 攀枝花与建昌聚为一类,攀枝花,建昌与云岭、乐至聚为一类,金堂与其它4个亲缘关系均比较远,两种聚类方法所得结果基本一致(图2、图3),说明聚类结果的精确性。
3 讨 论
3.1 攀西黑山羊的AFLP遗传多样性
AFLP标记技术采用不同类型限制性内切酶和不同数目选择性碱基,理论上可产生无限多标记数,并且覆盖整个基因组,被认为是指纹图谱中多态性最丰富的一项技术,目前常用的限制性内切酶组合是EcoRI和MseI[5-6]。
苟本富等[11]研究了AFLP在检测小香羊遗传多样性方面应用的可行性,用PstI酶切,10条引物共获得113条AFLP标记,群体相似系数为0.913(0.814-0.980),为小香羊的遗传稳定性提供了相关参数。本研究用15对引物对5个群体进行AFLP标记,共扩增出1 011个条带、1 003个多态条带,平均多态频率为99.01%,表明黑山羊群体间和群体内遗传多样性丰富。建昌遗传多样性最高,其次是攀枝花、金堂、云岭黑山羊,乐至黑山羊遗传多样性最低,说明建昌、攀枝花本地黑山羊遗传纯度较低,乐至黑山羊遗传变异纯度低,种质均匀性好,遗传纯度高。建昌黑山羊遗传多样性结果与李利等[14]的结果不一致,原因可能是所用分析方法不同。
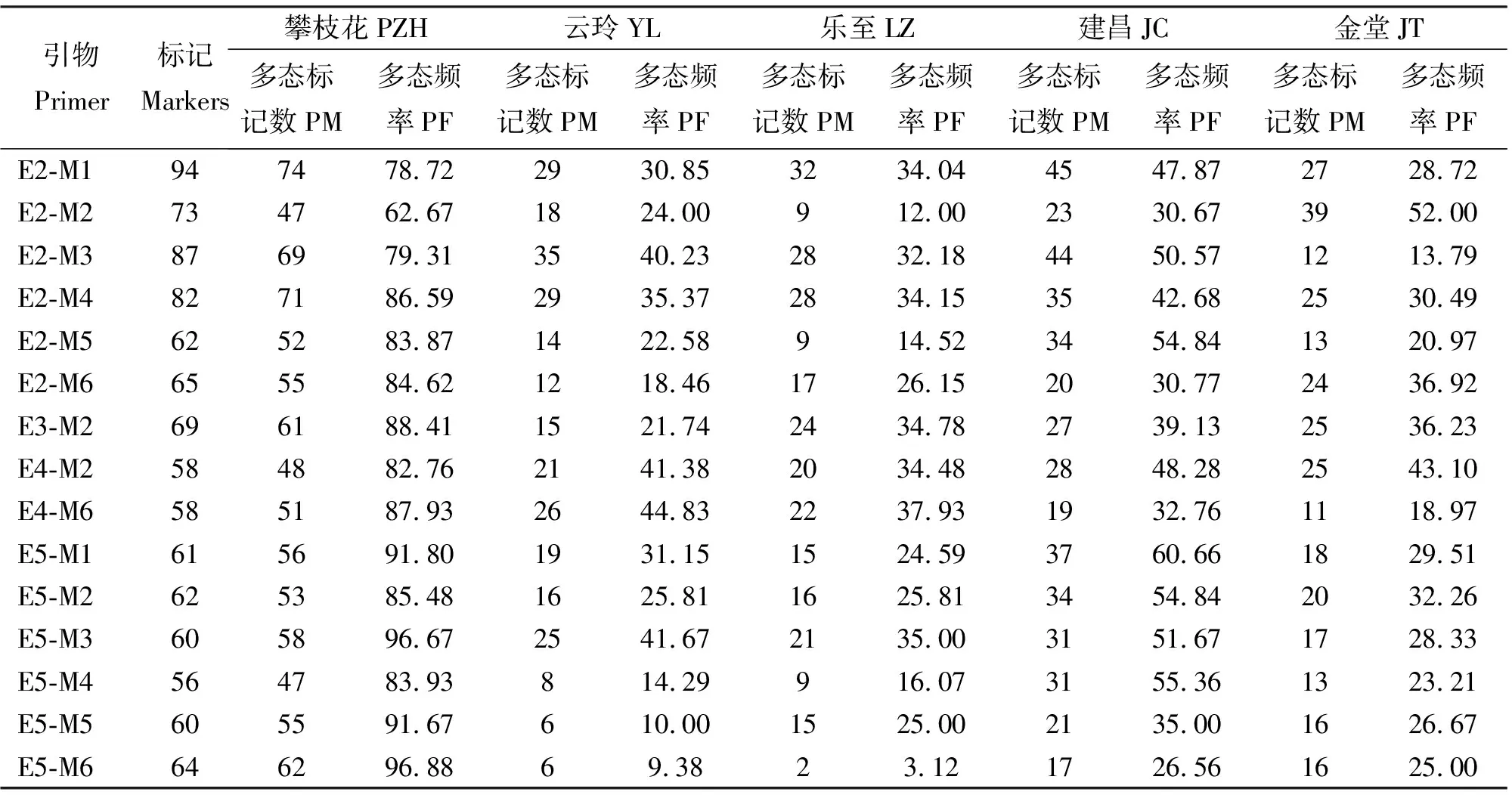
表3 5个黑山羊群体的多态位点数及多态频率Table 3 The number of polymorphic loci and polymorphic frequence among 5 black goat populations
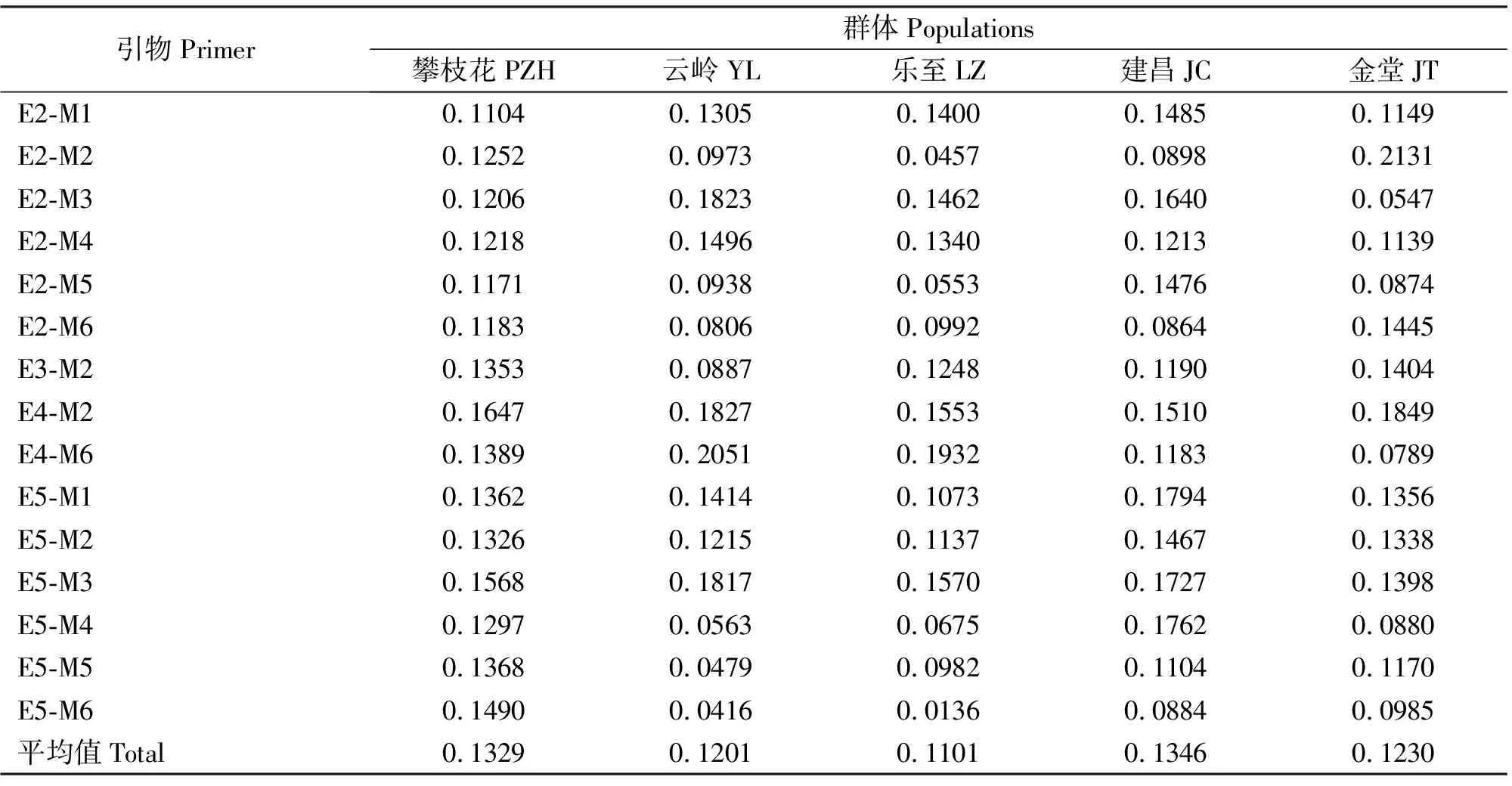
表4 5个黑山羊群体遗传多样性指数Table 4 The genetic diversity index of 5 black goat populations
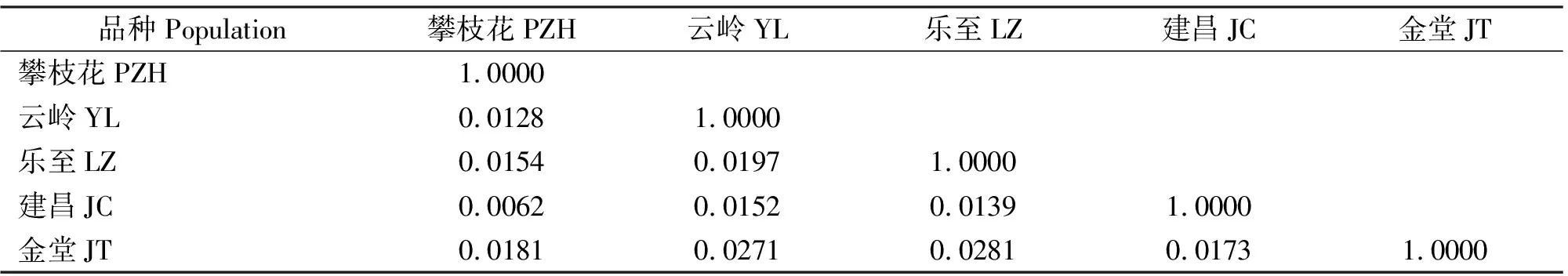
表5 5个黑山羊群体的遗传距离Table 5 The genetic distance of 5 black goat populations
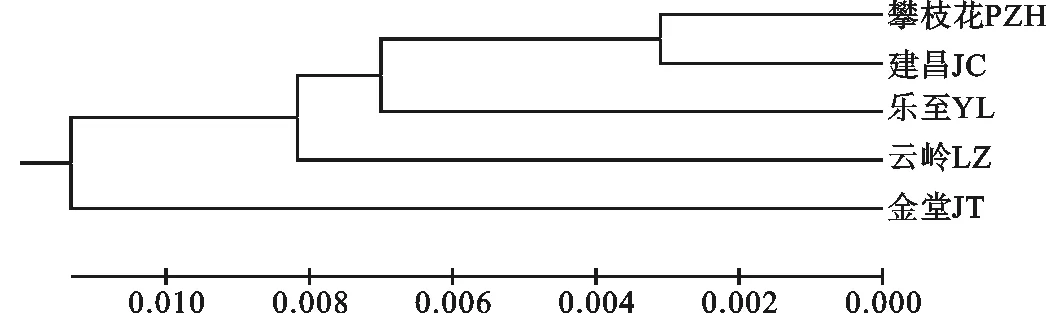
图2 5个黑山羊群体遗传距离的UPGMA聚类关系Fig.2 The cluster of 5 black goat populations based on UPGMA

图3 5个黑山羊群体遗传距离的NJ聚类关系Fig.3 The cluster of 5 black goat populations based on NJ
曹少先[12]等用AFLP标记对波尔山羊、徐汇山羊和海门山羊进行多样性分析,徐淮山羊与海门山羊具有最近的亲缘关系,而波尔山羊与徐淮山羊的亲缘关系较波尔山羊与海门山羊的亲缘关系近。肖非[13]等对新疆6个绵羊品种进行AFLP分析,所得遗传相似系数和聚类结果与各绵羊品种的地理分布及现实状况相吻合。本研究结果:PZH与JC聚为一类,PZH,JC与YL、LZ聚为一类, JT与其它4个亲缘关系均比较远。其中LZ和JT间遗传距离较小,与王杰等[7]的结果一致。会理县距离攀枝花市较近,位于攀枝花市东北方;龙潭乡与中坝乡距离较近,位于攀枝花市以南;云南云岭黑山羊分布较接近攀枝花市;金堂县所处位置最远,说明分类结果与群体地理分布、生态条件基本一致。
3.2 攀枝花市黑山羊资源的保护与利用
家畜遗传资源是畜牧业生产的工具,对其进行调查、正确分类有助于家畜资源的合理开发利用。四川各地黑山羊生产性能及遗传特征的研究,为各地黑山羊品种鉴定提供科学依据,也为黑山羊的选育、保种及开发利用奠定基础[15]。攀西地区拥有丰富的黑山羊种质资源,通过AFLP标记技术对攀枝花本地、云岭、乐至、金堂、建昌5个黑山羊群体进行研究,结果表明黑山羊品种间和群体内遗传多样性丰富,为攀枝花市本地黑山羊品种的鉴定和更好地开发和利用提供理论依据。
参考文献:
[1] Vos P, Hogers R, Bleeker M, et al. AFLP :anew technique for DNA fingerprinting[J]. Nucleic Acids Research,1995,21:4 407-4 414.
[2] Kin D H, Herber D, Still D W. Genetic diversity of Echinacea species based upon amplified fragment length polymorphism markers[J]. Genome, 2004,47(2):102-111.
[3] Jeremy J A, Seki S, Cnaan M A, et al. Breeding new strains of tilapia: development of an artificial center of origin and linkage map based on AFLP and microsatellite loci[J]. Aquaculture, 2000,185:43-56.
[4] Sambrook J, Russell D W. 分子克隆指南[M].3版.北京:科学出版社, 2002:483-485.
[5] 周延清, 杨清香, 张改娜. 生物遗传标记与应用[M]. 北京:化学工业出版社,2008:170-184.
[6] 周延清. DNA分子标记技术在植物研究中的应用[M]. 北京:化学工业出版社,2005:166-185.
[7] 王 杰, 马正花,王 永. 成都麻羊与7个山羊品种(群体)的AFLP遗传多样性研究[J].西南民族大学学报:自然科学版,2008, 34(4):688-693.
[8] Santos L M, Gama R A , Eiras A E,et al. Genetic differences based on AFLP markers in the mosquito species Anopheles darlingi collected in versus near houses in the region of Porto Velho, RO, Brazil[J]. Gentics and Molecular Research, 2010,9(4):2 254-2 262.
[9] Wang Da-Wei, Li Yu, Li Zhou-Qi. Identification of a male-Specific amplified fragment length polymorphism (AFLP) and a sequence characterized amplified region (SCAR) marker in eucommia ulmoides oliv[J]. International Journal of Molecular Science, 2011,12:857-864.
[10] Liu Wei-dong, Li Hong-jun, Bao Xiang-bo, et al. Genetic differentiation between natural and hatchery stocks of japanese scallop (Mizuhopecten yessoensis) as revealed by AFLP analysis[J]. International Journal of Molecular Science, 2010,11:3 933-3 941.
[11] 苟本富, 叶华虎,魏 泓, 等. AFLP标记在小香羊遗传多态性检测中的应用[J]. 动物学杂志,2003,38(5):45-49.
[12] 曹少先, 杨利国, 姜勋平, 等. 波尔山羊和江苏本地山羊的AFLP和RAPD 分析[J].中国农业科学, 2002, 35(10):1 291-1 296.
[13] 肖 非,付 永,石 廷, 等. 新疆6个地方绵羊品种遗传多样性AFLP分析[J]. 中国畜牧兽医, 2009,36(10):93-98.
[14] 李 利, 刘成建, 张国俊, 等. 用5个微卫星标记分析四川7个地方山羊品种的遗传关系[J]. 中国畜牧杂志,2009,45(3):8-11.
[15] 周光明, 李 强, 付昌秀, 等. 四川黑山羊品种(品群)资源现状及其发展对策[J]. 中国草食动物,2005(S):64-66.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:10:34
走向世界(2022年18期)2022-05-17 23:03:14
系统工程学报(2021年4期)2021-12-21 06:21:08
四川蚕业(2020年3期)2020-07-16 08:09:32
新世纪智能(高一语文)(2020年12期)2020-06-01 08:14:20
韩国语教学与研究(2020年3期)2020-03-17 08:08:28
校园英语·中旬(2019年11期)2019-11-26 10:01:06
西部皮革(2016年23期)2017-01-03 06:37:16
中国医药导报(2015年27期)2015-02-28 22:08:01
中国神经精神疾病杂志(2013年1期)2013-03-11 20:23:33
