
抗菌肽在畜牧业上的应用研究及问题分析
2013-11-24 03:59王志强张爱忠姜宁丁翠华陈志宝
黑龙江八一农垦大学学报 2013年5期
王志强,张爱忠,姜宁,丁翠华,陈志宝
(1. 黑龙江八一农垦大学动物科技学院,大庆 163319;2. 东营职业学院教育系)
抗菌肽是生物界中广泛存在的一类生物活性多肽,是非专一性的免疫应答产物,具有广谱抗菌作用。如今发现的抗菌肽的数量如此的巨大以至于我们很难将其细致地分类,只能基于他们的二级结构进行大体分类,Zasloff[1]报道,目前已经发现了大约500 种抗菌肽分子。根据抗菌肽的来源不同可分为:昆虫抗菌肽,如天蚕素(cecropins);哺乳动物抗菌肽,如防御素(defensins);两栖动物抗菌肽,如蛙皮素(magainins)等;植物抗菌肽,如硫素(thionins);微生物抗菌肽,如短菌肽、乳酸菌肽等。此外还有大量人工设计的抗菌肽,例如,Cecropin Md-Che Rc[2]、Cecropin A-Melittin[3]等。
1 抗菌肽的理化性质及其生物学特性
天然抗菌肽通常是由30 多个氨基酸残基组成的碱性小分子多肽,水溶性好,分子量大约为4 000道尔顿左右。大部分抗菌肽具有热稳定性,在100 ℃下加热10~15 min 仍能保持其活性。多数抗菌肽的等电点大于7,表现出较强的阳离子特征。同时,抗菌肽对较大的离子强度和较高或较低的pH 值均具有较强的抗性。此外,部分抗菌肽还具备抵抗胰蛋白酶或胃蛋白酶水解的能力。
与传统抗生素相比,抗菌肽的抗菌作用有如下几个特点:(1)广谱抗菌,对病毒、革兰氏菌、真菌、寄生虫等都具有高效的杀灭作用;(2)作用时间短,大多数抗菌肽都能在5 min 之内起作用;(3)作用专一,具有靶向性,对正常细胞作用不明显;(4)无残留[4]。抗菌肽的以上特点使其成为新型添加剂的研究热点。
2 抗菌肽的杀菌作用机制
实验表明,几乎所有的抗菌肽都主要作用于细胞膜,可将其作用模式分为两类:(1)膜破坏性模式(membrane disruptive):主要包括桶-板模式(barrel stave)、环形孔模式(toroidal)、地毯模式(carpet)和胶束簇集合体模式(micellar aggregate models);(2)非膜破坏性模式(non-membrane disruptive),即胞内攻击目标型(intracellular targets)[5]。尽管学者们并没有取得广泛的共识,但AMPs 的穿膜机制模型已经被很多学者所认可。细菌细胞膜富含带负电荷的磷脂头基,如磷脂酰甘油(PG)、心磷脂(CL)和磷脂酰丝氨酸(PS)。相反,哺乳动物细胞膜富含两性电荷的磷脂(净电荷为中性)如磷脂酰乙醇胺(PE)、磷脂酰胆碱(PC)和神经鞘髓磷脂(SM)。此外,作为哺乳动物细胞膜主要组成成分的胆固醇能够降低AMPs 对脂质双层的稳定作用,或者可以直接与AMPs 相互作用而起到中和电荷的效应。以上说明,细胞膜的成分在AMPs 的作用机制中发挥着重要角色,故AMPs 可选择性的攻击细菌细胞膜而非哺乳动物细胞膜。
3 抗菌肽饲料添加剂在畜禽生产中的应用研究
3.1 在仔猪生产中的应用研究
现代养猪生产中大多采用早期断奶技术,因而过早中断了母源抗体的供给,导致仔猪出现腹泻等健康和生长方面的问题。
黄木家等[6]研究发现,在断奶仔猪饲粮中添加抗菌肽替代血浆蛋白粉,可以有效缓解仔猪早期断奶应激综合征。黄茂侠等[7]在仔猪基础日粮中添加0.6%~0.8%的复合抗菌肽制剂能够提高仔猪对腹泻和呼吸道疾病的抵抗力,其作用效果与添加300 mg·t-1的阿莫西林相当。另外,宋青龙等[8]在仔猪日粮中添加300 mg·kg-1的抗菌肽比添加100 mg·kg-1的吉他霉素(50%)的对照组显著提高了平均日增重和平均日采食量,显著改善仔猪腹泻和显著提高了试验前期(15、25 d)猪瘟抗体阳性率(P<0.05)。王建华等[9]研究了在仔猪日粮中分别添加抗菌肽10 mg·kg-1和蛋氨酸锌1 200 mg·kg-1,以及两者相同水平配伍对仔猪血液红细胞计数和血小板计数的影响,结果表明,3种饲粮对2 种血液常规指标的影响均显著(P<0.05),两者配伍对血小板计数有明显的协同(P<0.05),而对血红细胞计数影响不明显。马文锋等[10]在仔猪日粮中添加0.3%的抗菌肽替代0.2%的氧化锌,研究其对35日龄仔猪腹泻率、生产性能及血清猪瘟抗体的影响。结果表明,0.3%的抗菌肽能显著降低仔猪腹泻率及饲料增重比,并提高日增重(P<0.05)。另外,对血清猪瘟抗体水平也有一定的改善作用。
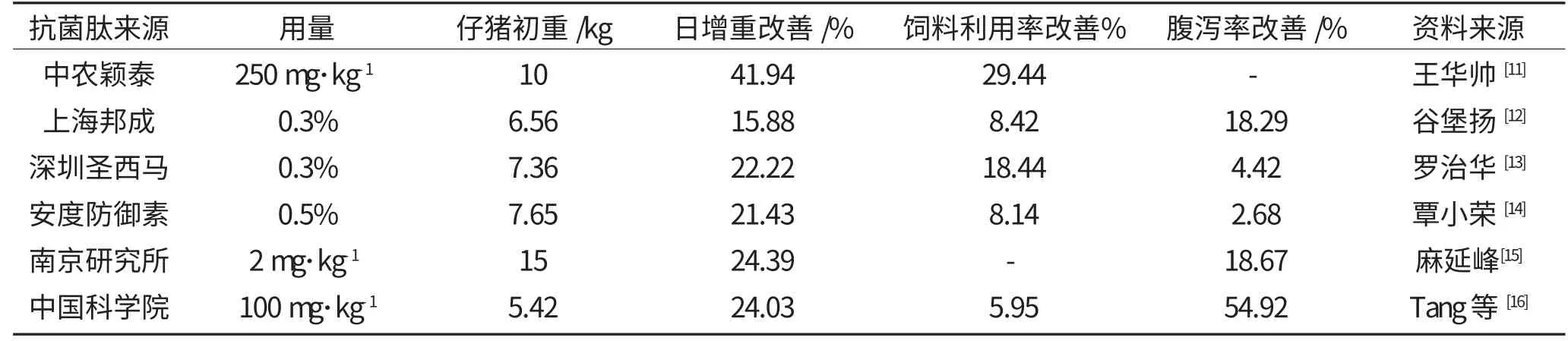
表1 部分抗菌肽产品及其在仔猪上的应用效果Table 1 The result of part of antimicrobial peptides'productions on piglet
以上的大量研究结果表明,在仔猪饲粮中添加抗菌肽,可以促进仔猪生长,改善饲料利用率,同时提高了仔猪的免疫功能,尤其在预防和治疗仔猪疾病方面的表现出色。
3.2 在母猪生产中的应用研究
天蚕素抗菌肽在母猪饲料上的应用试验近年来也比较多,论证了天蚕素对母猪的生产性能以及免疫水平等方面的影响。
李永新等[17]于母猪产前35 d 在饲料中添加抗菌肽饲料添加剂进行对比试验,观察母猪产仔及仔猪健康状况。结果表明,试验组母猪的窝均活仔重量有所提高,分娩、断奶的成活率分别提高了6.48%和16.10%,试验组比对照组提高259.08 元/窝的经济效益。潘行正等[18]研究抗菌肽制剂对母猪生产性能与健康水平的影响,结果表明,添加抗菌肽制剂的试验组母猪死产率仅为5.77%,而使用抗生素的对照组的死产率为12.75%,抗菌肽组哺乳期和保育期的仔猪死亡率为0.34%、1.31%,而抗生素组分别为2.35%和3.65%。可见抗菌肽制剂对提高母猪生产性能及仔猪健康水平效果显著。李波等[19]从母猪分娩前30 d 到产后21 d 在日粮中添加400 kg·mg-1天蚕素抗菌肽,对照组添加400 kg·mg-1的阿莫西林和300 kg·mg-1的支原净,试验组的腹泻率显著低于对照(P<0.05),而活仔率和健仔率均极显著高于对照(P<0.01)。郭强等(2012)[20]在日粮中添加200 mg·kg-1的天蚕素抗菌肽,试验结果与正常猪场保健程序相比,健仔数明显增加(P<0.05),仔猪腹泻率降低并提高了日增重。皮灿辉等[21]在母猪日粮中使用“肽轻松”抗菌肽制剂,研究其对母猪猪瘟抗体水平的影响,其试验结果显示,抗菌肽促进猪瘟抗体水平提高,且随着使用时间的延长猪瘟抗体水平增加。由此可见,抗菌肽能提高母猪免疫力,提高其健康水平。
综上所述,天蚕素抗菌肽对母猪的繁殖性能、免疫机能以及疾病预防等方面都具有良好的应用效果,并能在影响母体的情况下,间接对初生仔猪也产生益处。这不但提高了养猪的效益,并且为生产绿色健康的猪肉产品提供了新的途径。
3.3 在家禽生产中的应用研究
3.3.1 抑菌杀菌及维持菌群平衡
抗菌肽具有广谱抗菌活性,对多种细菌和真菌起作用,有些抗菌肽还能对抗原虫和病毒。陈红羽等[22]研究了抗菌肽Cecropin AD 对肉鸡大肠杆菌K88 攻毒状态下,日粮中添加抗菌肽对其肠道微生物和血浆IL-6 的影响。结果表明,日粮中添加了300 mg·kg-1Cecropin AD 的试验组与空白组比较,cecropin AD 能有效降低肉鸡盲肠大肠杆菌的数量(P<0.01),缓解由灌服大肠杆菌引起的应激反应,低于金霉素(80 mg·kg-1)处理组(P>0.05)。血浆IL-6 的分泌水平为Cecropin AD 组>金霉素组>空白组,且Cecropin AD 组与其他两组差异显著(P<0.05)。这表明Cecropin AD 可以抑制大肠杆菌数目、控制炎性反应来保证肉鸡生产效率。陈晓生等(2006)[23]在肉鸭日粮中添加Cecropin AD 制剂发现,对金霉素产生耐药性的大肠杆菌在抗菌肽的作用下抑制效果明显(P<0.05)。显然,抗菌肽能显著地抑杀体内大肠杆菌,对促进肠道微生物菌群平衡、保障动物生长性能的提高有重要作用,抗菌肽调节鸡、鸭肠道微生物菌群平衡的能力优于金霉素。温刘发等(2009)[24]研究发现Cecropin AD 对肉鸡柔嫩艾美耳球虫也具有良好的抑制效果。
3.3.2 提高家禽生产性能
马倩等(2012)[25]将鲎素抗菌肽分别以0.1 g·kg-1和0.1 g·kg-1+0.5 亿酵母活菌·kg-1制剂的形式添加到日粮中进行芦花鸡饲养试验,其研究结果表明:在芦花鸡日粮中添加鲎素抗菌肽,能明显提高芦花鸡的生产性能,且添加抗菌肽制剂效果更好。同时,抗菌肽制剂和抗菌肽能促进芦花鸡体内蛋白质的合成,增加蛋白质的沉积。在芦花鸡生长初期,抗菌肽制剂促进脂肪的沉积。卢宇等[26]将抗菌肽s807 分50~300 mg·kg-1等5 个剂量,分别在不同的时间段添加到颗粒料和粉料给10~35 日龄肉仔鸡饲喂7 d 以上,结果肉鸡的死淘率从16.5%下降至7.88%;每天添加饲喂可使肉鸡平均增重11.0~16.3 g,生产性能增加18.6%~22.1%,其试验证实抗菌肽可以显著地提高雏鸡成活率,促进肉仔鸡生长。吕尊周等[27]研究了枯草芽孢杆菌中提取的抗菌肽粗制品对蛋鸡产蛋性能和蛋品质的影响,其研究结果显示,蛋鸡日粮中添加400 mg·kg-1的抗菌肽粗提物就能显著提高淡季的产蛋率和蛋黄相对重(P<0.05)。
3.3.3 调控家禽机体免疫功能
在抗菌肽作用机理的基础研究中人们已经发现其对机体免疫功能产生的重要作用。在家禽的应用研究中,抗菌肽对免疫指标的影响较大。可能与实验动物、抗菌肽种类和质量等因素有关,不同试验的结果之间有时存在一定差异。
陈晓生等[28]对不同日龄肉鸭日粮添加抗菌肽制剂的试验结果显示,IGF-I 水平与金霉素组均显著升高,甲状腺素水平T3 显著升高而T4 降低,这与金霉素组相反。而在马倩等[29]的研究中,抗菌肽能显著提高14 日龄芦花鸡T4 水平(P<0.05)以及84 日龄的IGF-I 水平,而42 日龄的T3 水平则显著降低;抗菌肽制剂对血清生长激素水平无显著影响(P<0.05)。吕尊周等[30]研究了抗菌肽对蛋鸡血清免疫指标及脾脏白细胞介素2 mRNA 表达量的影响,其研究结果表明,抗菌肽能提高血清IgA、IgG 的含量,当添加量为400 mg·kg-1时达到显著水平(P<0.05)。
姚俊等[31]研究了外源性抗菌肽——猪小肠抗菌肽对SPF 鸡肠道黏膜免疫功能的影响,结果显示外源性猪小肠抗菌肽能显著提高SPF 鸡的肠道黏膜上皮淋巴细胞数、肥大细胞数、杯状细胞数以及sIgA 等免疫参数。刘莉如等[32]的研究结果显示,饲粮添加天蚕素抗菌肽可显著改善海兰褐仔公鸡小肠黏膜上皮细胞形态,同时也增加了小肠免疫活性细胞的数量。这些研究成果为动物肠道免疫防御能力的提高提供了新思路。
3.4 在奶牛疾病预防和治疗方面的应用研究
奶牛乳房炎是一种发病率极高的奶牛常见病,多种病原微生物如病毒、细菌、真菌、支原体、放线菌甚至寄生虫等都可使奶牛患病。有些奶牛乳房炎极难治疗。奶牛患乳房炎后身体质量下降,不仅导致产奶质量下降,产奶量降低,造成经济损失,而且对人的健康也构成威胁。抗生素在治疗奶牛乳房炎的过程中极易导致残留,且容易产生耐药性。方维焕等[33]的研究表明乳铁素(Lactoferricin)是一种广谱抗菌多肽,但同时却参与乳房链球菌对乳腺上皮细胞的粘附,其经过试验探讨认为,制备乳铁素结合蛋白可能是免疫预防乳腺炎的一种有效途径。乳酸链球菌素(Nisin)是一种小分子抗菌肽,对革兰氏阳性菌和支原体有很好的抑制效果。曹立亭等[34]对患临床型乳房炎的奶牛进行乳房灌注Nisin 治疗后,患病乳区牛奶乳脂肪、乳蛋白、乳糖和非脂乳固体含量都呈现上升趋势,其中,乳糖含量除停药后2 d 的治疗乳区外,均在4.7%以上,表明奶牛乳房炎病例经Nisin 乳房灌注治疗后,受损乳腺组织修复较快,乳腺上皮细胞合成乳成分的能力大幅度提高。徐名能等(2011)[35]在奶牛日粮中添加50 g·(头·d)-1的抗菌肽制剂(肽轻松C),试验结果显示抗菌肽制剂显著提高了乳脂率(P<0.05),并降低了61.43%的牛乳体细胞数(P<0.01)。
3.5 在肉兔生产中的应用研究
雷岷等[36]的研究表明,200 mg·kg-1左右的抗菌肽能显著降低料重比,并显著提高肉兔胸腺指数和脾脏指数。另外,抗菌肽还改善了肉兔小肠黏膜形态,并对盲肠内的有益菌增殖有促进作用[37]。
3.6 在水产动物生产中的应用研究
抗菌肽在水产饲料中的应用日益广泛,其促生长效果显著,已得到了实验和生产的验证。黄自然等[38]在南美白对虾中进行的试验表明,每千克饲料添加2~4 mL 抗菌肽提高了对虾的孵化率、出苗率和收获量,且高于5 mg·kg-1金霉素组。黄沧海等[39]在罗非鱼幼鱼饲料中添加抗菌肽,对比黄霉素发现对促进鱼体增重的效果没有显著差异,且两者的促生长效果优于喹乙醇。喹乙醇可在体内蓄积,对大多数动物有明显的致畸作用,对人也有潜在的危害,因此在水产养殖方面,用抗菌肽替代抗生素,不仅可以提高鱼体的免疫抗病力,而且可以避免抗生素在鱼体中的蓄积,提高水产品质量。
4 抗菌肽开发与应用中存在的问题
大量的研究表明,抗菌肽在畜牧业上的应用必将为畜牧业的发展带来新的推动力。但要将抗菌肽全面推向市场还需要进一步的研究,其主要的问题有:(1)商品化成本较高,基因工程生产技术有待进一步突破。(2)抗菌肽多为异源性,尚缺少药效动力学和毒理学等的基础研究数据,用药安全问题有待进一步解决。而更重要的问题是,尽管有报道称抗菌肽很难产生耐药性[1],但抗菌肽的耐药性问题仍然是限制其开发应用的核心问题。例如,细菌使用各种策略来避免被抗菌肽杀死[40]。金黄色葡萄球菌可分泌蛋白酶降解抗菌肽,还可以改变膜的转运作用从而对蛙类抗菌肽产生耐药性[41]。还可将细胞质内的D-丙氨酸运输到细胞膜表面的磷壁酸,通过将碱性氨基介入到细胞膜上来降低负电荷数,还能通过MprF 用L-赖氨酸来修饰其阴离子膜,增加其正电荷数量[42]。克雷伯菌属可以通过荚膜多聚糖限制抗菌肽与细胞膜上靶位的交互作用[43]。沙门菌属将2-羟基肉豆蔻酸盐或将棕榈酸盐加到脂质a 上,然后在细胞膜上增加这些经修饰的脂质a,从而降低了外细胞膜的流动性,进而减少了与抗菌肽的疏水性交互作用。而增加的疏水力矩则减轻或消除了抗菌肽插入细菌细胞膜并在上面形成针孔结构的能力,另外,细胞膜蛋白质残基的修饰和更改也在进行中。在一些革兰氏阴性菌中,通过修饰外膜蛋白产生了对抗菌肽的耐受力[42]。一种无法分类的流感嗜血杆菌将抗菌肽运输到细胞内的特殊位置并将其降解掉;H. 流感改变其细胞膜,使其看起来“好像”被抗菌肽攻击了,从而避免了更多抗菌肽的攻击[44];两种转运蛋白:ATP 结合匣式转运子和抗结瘤细胞分裂流出泵分别能将抗菌肽运进和运出细胞[45],这都与细菌对抗菌肽的耐药性相关。另外,细菌还产生蛋白质水解酶来降解抗菌肽[46]。
以上这些举例表明细菌耐药性是可以自然进化的,这导致了人们越来越担忧抗菌肽的大量使用会使细菌耐药性的发展更快、更完善。有些研究还表明,在有些情况下,抗菌肽作为治疗药物使用后也可能使机体本身产生的类似抗菌肽不再对细菌有效[47]。
因此,抗菌肽的应用研究成果固然可喜,但对于抗菌肽的抗菌机制以及耐药性等问题的基础性研究也已经迫在眉睫。
[1] Zasloff,Michael. Antimicrobia l peptides of multicellular organisms[J]. Nature,2002,415:389-395.
[2] 孙艳发,张爱忠,姜宁. 杂合抗菌肽Cec Md-Che Rc 的分子设计及其克隆载体的构建[J]. 黑龙江八一农垦大学学报,2009,21(6):47-50.
[3] 储卫华,李大力,汪信. 天蚕素A-蜂毒素杂合肽基因表达载体的构建及其分泌表达[J]. 南京理工大学学报,2007,31(2):257-261.
[4] 陈星星,陈宏惠,郑新添. 抗菌肽的研究进展及其在畜牧业上的运用[J]. 福建畜牧兽医,2011,23(1):20-23.
[5] 吴德伟. 抗菌肽的功能及作用机理[J]. 科技信息,2008,33(22):92-93.
[6] 黄木家,刘文娟,李永新. 抗菌肽制剂替代血浆蛋白粉对断奶仔猪生长性能及健康状况的影响[J]. 中国饲料,2011(3):43-44.
[7] 黄茂侠. 抗菌肽对预防断奶仔猪腹泻和呼吸道疾病的作用[J]. 中国畜禽种业,2011(6):72-74.
[8] 宋青龙,王洪彬,张广民,等. 日粮中添加天蚕素抗菌肽对仔猪健康及生产性能的影响[J]. 动物保健,2012,48(13):52-58.
[9] 王建华,伍淳操. 抗菌肽和蛋氨酸锌对断奶仔猪部分血液常规指标的影响[J]. 贵州农业科学,2010,38(3):127-130.
[10] 马文锋,焦喜兰,李振田,等. 抗菌肽替代氧化锌对仔猪生产性能及血清猪瘟抗体水平的影响[J]. 中国饲料,2011(19):22-24.
[11] 王华帅,贾山松,雷东风. 天蚕素抗菌肽对断奶仔猪生长性能的影响[J]. 养猪,2011(2):11.
[12] 谷保扬,郑云峰,陆克文. 肽健宝对断奶仔猪生产性能的影响[J]. 中国饲料添加剂,2010(8):32-33.
[13] 罗治华,周洪波,苏向东. 抗菌肽饲料添加剂对断奶仔猪生产性能的影响[J]. 当代畜牧,2009(7):23-24.
[14] 覃小荣,刘丁健,曾其恒. 抗菌肽对保育猪生产性能与健康水平的影响[J]. 饲料研究,2011(4):6-8.
[15] 麻延峰,王宏艳,周文仙. 抗菌肽制剂对提高金华猪生长性能的效果[J]. 浙江农业科学,2010(4):82-89.
[16] Tang Z R,Yin Y l,Zhang Y M,et al. Effects of dietary supplementation with an expressed fusion peptide bovine lactoferrcin -lactofe rrampin on performance,immune function and intestinal mucosal morphology in piglets weaned at age 21d[J]. British Journal of Nutrition,2009(101):998-1005.
[17] 李永新,王强,周洪波,等. 抗菌肽饲料添加剂对母猪生产性能的影响[J].饲料工业,2009,30(15):19-21.
[18] 潘行正,黄正明,李永新. 抗菌肽制剂对母猪死产率和仔猪成活率的影响[J].现代农业科技,2010(12):285-286.
[19] 李波,杨利,易学武,等. 日粮中添加天蚕素抗菌肽对母猪繁殖性能的影响[J]. 中国畜牧兽医,2011,38(1):26-29.
[20] 郭强,何涛. 妊娠母猪日粮中添加天蚕素抗菌肽对繁殖性能及产后仔猪生长性能的影响[J].抗菌肽应用技术,2012(12):54-58.
[21] 皮灿辉,王凯,谢灯养,等. 抗菌肽制剂对母猪猪瘟抗体水平的影响[J]. 养猪,2008(5):11-13.
[22] 陈红羽,谯仕彦.日粮中添加抗菌肽cecropin AD 对肉鸡大肠杆菌攻毒条件下肠道菌群和白介素-6 水平变代研究[J]. 抗菌肽应用技术,2012(5):51-53.
[23] 陈晓生,张辉华,周庆国,等. 抗菌肽对肉鸭雏鸭期肠道主要微生物菌落的影响[J]. 兽药与饲料添加剂,2006,11(4):4-6.
[24] 温刘发,李国清,马海涛,等. 抗菌肽Cecropin AD 抗肉鸡柔嫩艾美耳球虫效果的研究[J].中国动物保健,2009(7):95-96.
[25] 马倩,赵衍铜,柏明娜,等. 抗菌肽对芦花鸡生产性能和血清生化指标的影响[J]. 饲料研究,2012(3):27-30.
[26] 卢宇,王素珍,王凯. 抗菌肽s807 提高肉鸡生产性能的研究[J]. 广东畜牧兽医科技,2010,35(4):17-21.
[27] 吕尊周,袁肖笑,朱志刚,等. 抗菌肽粗提物对蛋鸡产蛋性能蛋品质血清生化指标和肠道菌群的影响[J]. 中国畜牧杂志,2012,48(15):47-52.
[28] 陈晓生,刘为民,温刘发,等. 抗菌肽替代抗生素对肉鸭生产性能及血清代谢激素水平的影响[J]. 中国家禽,2005,27(5):7-9.
[29] 马倩,赵衍铜,柏明娜,等. 抗菌肽对芦花鸡血清激素水平的影响[J]. 饲料工业,2012,33(9):17-20.
[30] 吕尊周,袁肖笑,蔡兆伟,等.抗菌肽对蛋鸡血清免疫指标及脾脏白细胞介素2 mRNA 表达量的影响[J]. 动物营养学报,2011,23(12):2183-2189.
[31] 姚俊,王德成,佘锐萍,等. 猪小肠抗菌肽对SPF 鸡肠道黏膜免疫功能的影响[J]. 科学与技术工程,2009(17):4918-4924.
[32] 刘莉如,杨开伦,滑静,等. 抗菌肽对海兰褐仔公鸡小肠黏膜形态结构及免疫活性细胞数量的影响[J]. 动物营养学报,2013,25(1):190-197.
[33] 方维焕,张晓峰. 乳铁素参与乳房链球菌对乳腺上皮细胞的粘附[J]. 中国兽医学报,2003,23(1):35-37.
[34] 曹立亭,胡松华. 奶牛临床型乳房炎Nisin 抗菌肽治疗后日产奶量及主要乳成分的变化[J]. 中国兽医学报,2010,30(12):1686-1689.
[35] 徐名能,黄木家,李永新,等. 日粮添加抗菌肽制剂对荷斯坦牛乳中体细胞数的影响[J]. 中国奶牛,2011(16):41-43.
[36] 雷岷,郭志强,任永军,等. 天蚕素抗菌肽对肉兔生产性能和免疫器官指数的影响[J]. 中国饲料,2012(9):25-28.
[37] 郭志强,杨奉珠,雷岷,等. 抗菌肽对肉兔小肠黏膜形态、盲肠菌群和免疫功能的影响[J]. 动物营养学报,2012,24(9):1778-1784.
[38] 黄自然,黄国庆,张敬炬. 新型饲料添加剂抗菌肽养殖对虾的效果[J]. 广东蚕业,2006(3):23-28.
[39] 黄沧海,李波,王冬冬. 抗菌肽对罗非鱼幼鱼生长性能的影响[J]. 中国畜牧杂志,2009,45(23):53-56.
[40] Perron G G,Zaslof M,Bell G. Experimental evolution of resistance to an antimicrobial peptide[J]. Proc Biol Sci,2006,273:251-256.
[41] Brogden K A. Antimicrobial peptides:pore formers or metabolic inhibitors in bacteria [J]. Nature Reviews Microbiology,2005,3(3):238-250.
[42] China B,N'guyen B T,De Bruyere M,et al. Role of Yad A in resistance of Yersinia enterocolitica to phagocytosis by human polymorphonuclear leukocytes [J]. Infection and immunity,1994,62(4):1275-1281.
[43] Campos M A,Vargas M A,Regueiro V,et al.Capsule polysaccharide mediates bacterial resistance to antimicrobial peptides [J]. Infection and Immunity,2004,72(12):7107-7114.
[44] Catherine L,Shelton,Forrest K. Sap transporter mediated import and subsequent degradation of antimicrobial peptides in Haemophilus [J]. PLos Pathogens,2011,7(11):102-135.
[45] Nikaido H. Multidrug efflux pumps of gram-negative bacteria [J]. Journal of Bacteriology,1996,178(20):5853-5859.
[46] Whitelock J M,Murdoch A D,Iozzo R V. Underwood,P.A. The degradation of human endothelial cell-derived perlecan and release of bound basic fibroblast growth factor by Stromelysin, Collagenase, Plasmin, and Heparanases [J]. Journal of Biological Chemistry,1996,271(17):10079-10086.
[47] Habets,Michelle,Brockhurst. Therapeutic antimicrobial peptides may compromise natural immunity [J]. Biology Letters,2012,8(10):1098-1203.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13
中国动物保健(2021年4期)2021-07-21
今日农业(2020年18期)2020-12-14
数学小灵通·3-4年级(2018年12期)2018-01-26
中成药(2017年4期)2017-05-17
中国蚕业(2017年1期)2017-04-08
吉林农业(2016年23期)2016-12-22
广东饲料(2016年5期)2016-12-01
国外医药(抗生素分册)(2016年5期)2016-07-12
中国中医药现代远程教育(2014年11期)2014-08-08
