
转基因(Bn-csRRM2)高产棉花的荒地生存竞争能力
2013-11-22 05:40马小艳姜伟丽马亚杰
生物安全学报 2013年4期
马小艳,彭 军,姜伟丽,马亚杰,马 艳
中国农业科学院棉花研究所,棉花生物学国家重点实验室,河南 安阳455000
作物高产是农业科技工作者长期追求的目标。前人研究发现,水稻开花控制基因FCA 的RNA 识别基序(RNA recognition motif,RRM)过量表达具有增大细胞和提高产量的功能(Hong et al.,2007);同时,在油菜Brassica napus L.中过量表达油菜FCA的第2 个RRM 结构域基因(Bn-csRRRM2)也表现出细胞和整个植株的增大(孙凡,2010)。这说明FCA-RRM2 可能参与细胞大小的调控,同时,其在进化过程中表现出高度的保守性。Sun et al.(2012)将Bn-csRRRM2 基因转入棉花获得的转基因棉花,与受体材料相比表现出植株和多种器官体积的增大,同时棉花的产量和品质显著提高,使其在农业生产中具有重大的经济价值。
然而,转基因作物在给人类带来巨大的经济和社会效益的同时,其对人类健康和生态环境可能造成的影响越来越受到人们的关注(宋新元等,2011;Andow & Zwahlen,2006)。对转基因作物进行环境安全性评价是其商业化种植之前的一项重要评价内容,其中,转基因作物的荒地生存竞争能力,即其转变为杂草的可能性,是转基因作物环境安全性评价的重要内容之一(刘娜等,2006;James,1996)。近年来,人们依据农业部953 号公告(中华人民共和国农业部,2007),对多种转基因棉花的荒地生存能力进行了评价,结果表明,转双价抗黄萎病基因(Glu+Chi)棉、转双价抗虫基因(cry1Ac +cry2Ab)棉和转基因(EPSPS+cry1Ac)抗草甘膦抗虫棉在与杂草的竞争中均处于弱势,生长势、种子活力及越冬能力等方面与非转基因植株差别不大,不能繁殖传代,无杂草化风险(马小艳等,2013;王振宇等,2010;张兴华等,2011、2012)。
本文以转基因高产棉为研究对象,依据农业部953 号公告,对其荒地生存竞争能力进行评价,以期为转基因高产棉的推广应用及转基因棉花环境安全评价技术的完善提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为转基因(Bn-csRRRM2)高产棉HN9311,受体材料中棉所12(CK1)及当地主栽品种转基因(Bt+CpTI)抗虫棉中棉所79(CK2)为对照品种,均由中国农业科学院棉花研究所育种研究室提供。
1.2 试验处理
试验于2013年在河南安阳中国农业科学院棉花研究所试验农场的杂草圃中进行。试验品种和对照品种各设4 个处理:分别于4月25日和5月25日播种,播种方式均为地表撒播和3 cm 深度点播2 种。每个小区播种40 粒种子,播种后未进行任何栽培管理。每个处理设3 次重复,随机排列,小区面积为1.6 m×3 m。
杂草调查:棉花播种前目测每个小区内杂草的覆盖度,即小区内全部杂草投影所覆盖小区面积的百分比。以后每隔1 个月调查1 次杂草覆盖度,至9月底结束。
棉花存活率及生长发育调查:棉花播种后每隔1 个月详细调查各小区的棉花株数,随机测量10 株棉花的株高、茎直径、果枝数、蕾铃数等生育指标;若小区内棉株少于10 株,则测量所有棉花的生育指标,至9月底结束。
1.3 统计分析
运用SPSS 13.0 软件对所有数据进行统计分析,采用单因素方差分析(One-way analysis,ANOVA)和Duncan's 差异显著性分析,检验同一时期不同小区内杂草发生情况的差异,以及棉花生长参数在不同品种之间的差异。
2 结果与分析
2.1 杂草发生情况
经Duncan's 差异显著性分析,2 种播种方式下,同一时期不同处理区间杂草的覆盖度无显著性差异(表1),且杂草覆盖度在整个调查期内呈现逐渐增大的趋势。4月底和5月底播种时,绝大多数杂草处于苗期,植株矮小,杂草平均覆盖度为30.0% ~65.0%,至7月底已达到100%,最终形成以小飞蓬Conyza canadensis (L.)Cronq.、狗尾草Setaira viridis (L.)Beauv.、马唐Digitaria sanguinalis (L.) Scop.和 女 菀 Turczaninowia fastigiata(Fisch.)DC.为绝对优势种的杂草群落。
2.2 4月底播种竞争能力表现
2.2.1 存活率 调查结果(表2)显示,4月底播种,撒播方式下棉花的出苗率高于深播,但2 种播种方式下棉花均延缓出苗,撒播方式下5月底仍未见棉花出苗。这主要是由于4月份降雨量较少,土壤干旱不利于棉花出苗,5月底至6月初的降雨促进了棉花的萌发。同一播种方式下,转基因高产棉花的存活率略高于受体材料,但两者差异不显著;而转基因高产棉花的存活率显著高于当地主栽对照品种中棉所79。上述结果表明,荒地4月底撒播和深播棉种,在不进行任何栽培管理的情况下,转基因高产棉花的存活率与受体材料和对照品种相比具有一定的竞争优势。
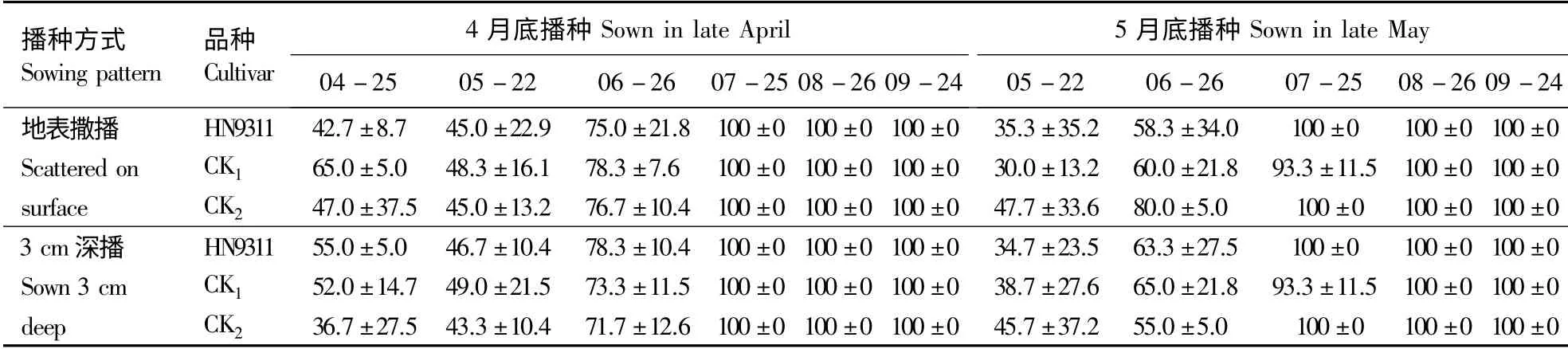
表1 2 种播种方式下小区杂草的覆盖度Table 1 Weed coverage in each district by two sowing patterns %

表2 2 种播种方式下棉花在荒地中的存活率Table 2 Cotton survival rates under two sowing patterns in non-cultivated land %
2.2.2 营养生长竞争力 由表3可以看出,若4月底播种,同一播种方式下,转基因高产棉花与两对照品种相比,株高和茎直径均无显著差异。从7月底至9月底,由于杂草的荫蔽作用,棉花生长缓慢,株高和茎直径无明显增加。这说明荒地4月底撒播和深播棉种,在不进行任何栽培管理的情况下,转基因高产棉花的营养生长与受体材料和对照品种相比无竞争优势。在杂草的荫蔽下,3 个品种在荒地中均生长缓慢,至9月底,棉花平均株高仅为9.7 ~32.9 cm,茎直径仅为1.9 ~4.7 mm。
2.2.3 生殖生长竞争力 4月底撒播方式下,至8月底,转基因高产棉花和对照品种中棉所79 仍未见果枝和蕾铃,仅见1 株受体材料有3 台果枝和3个花蕾。至9月底,3 个品种的个别植株均长出果枝,但果枝数较少,仅1 ~5 台,平均果枝数小于0.3台·株-1;转基因高产棉花未见花蕾,仅有1 株上见1 个大铃;同时,两对照品种的花蕾数也较少,平均为0.2 ~0.3 个·株-1,仅有1 株受体材料上有1个小铃(棉铃未出苞叶),无吐絮表现,而中棉所79未见棉铃(表4)。
由表4可见,同一时期同一品种深播处理的棉花比撒播处理生育进程略快。至8月底,深播方式下,除对照品种中棉所79 无果枝、蕾铃外,转基因高产棉花和受体材料均长出果枝和花蕾,但数量均较少,此时3 个品种均未见棉铃。至9月底,中棉所79 仍无果枝和蕾铃,而转基因高产棉花和其受体材料的部分植株长出果枝,但果枝数较少,平均果枝数小于0.5 台·株-1;虽然转基因高产棉花和其受体材料的蕾铃数多于撒播处理,但是仅见转基因高产棉花有2 株棉花各有1 个吐絮铃,且棉铃较小,直径仅2 cm 左右,棉籽发育不成熟,不能正常发芽繁殖。
上述结果表明,4月底撒播和深播2 种方式下,转基因高产棉与其受体材料和当地主栽棉相比,生殖生长的竞争优势不明显,在不进行任何栽培管理的环境下,无变为杂草的可能性。

表3 2 种播种方式下棉花的株高和茎直径Table 3 Cotton height and stem diameter under two sowing patterns
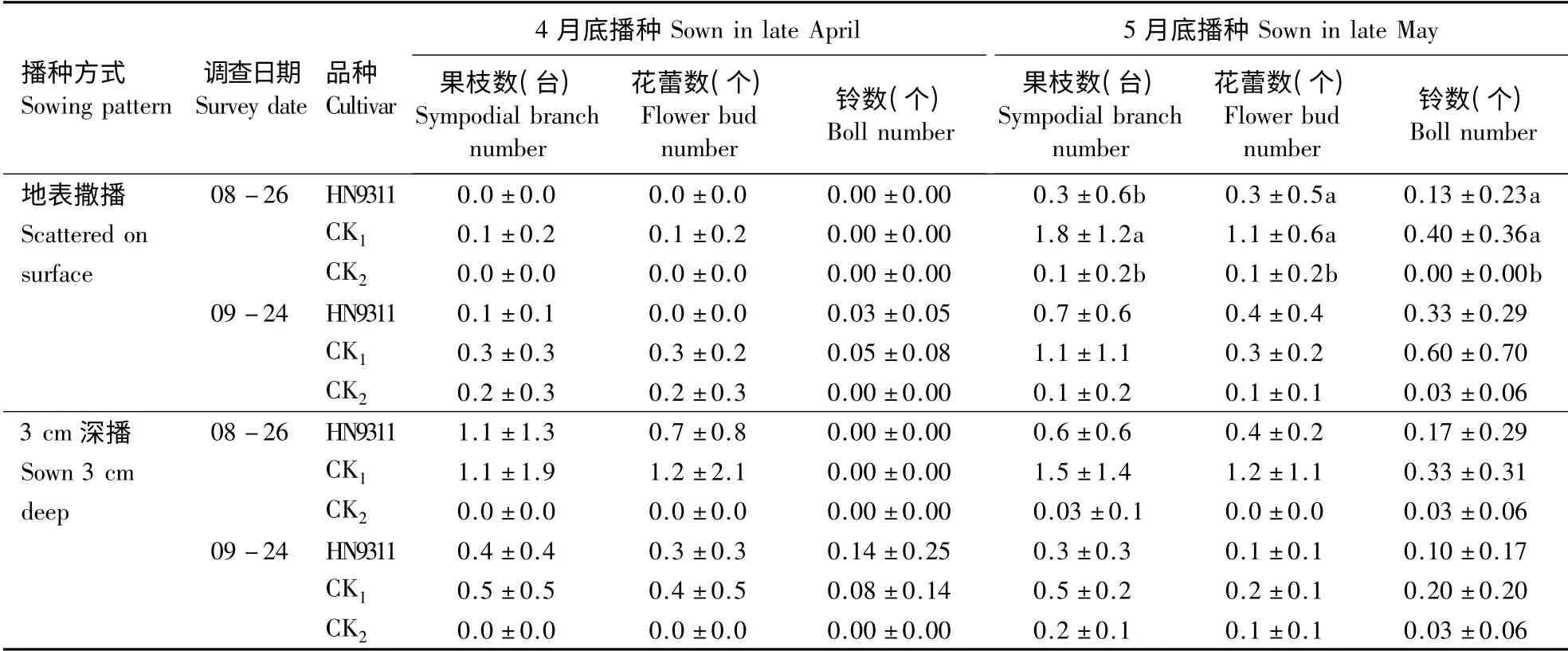
表4 2 种播种方式下棉花的果枝数、花蕾数和铃数Table 4 The number of sympodial branch,bud and boll under two sowing patterns
2.3 5月底播种竞争能力表现
2.3.1 存活率 5月底播种时,降雨量较大,土壤湿度大,有利于棉花的出苗,因此,5月底播种的棉花出苗情况好于4月底播种(表2)。撒播和深播方式下,转基因高产棉花与受体材料的存活率均(显著)高于对照品种中棉所79。随着杂草生长对棉花的荫蔽作用,3 个品种的存活率整体略呈下降趋势,但下降程度不大,至9月底,2 种播种方式下,棉花的存活率仍在50%左右。这说明荒地5月底撒播和深播棉种,在不进行任何栽培管理的情况下,与当地主栽品种相比,转基因高产棉花及其受体材料具有一定的存活竞争优势。
2.3.2 营养生长竞争力 同一时期,转基因高产棉花与受体材料的株高和茎直径相当,均大于对照品种中棉所79,但3 个品种间差异不显著(表3)。这说明外源高产基因的导入并未提高受体材料营养生长的竞争力,但荒地5月底撒播和深播棉种,在不进行任何栽培管理的情况下,与当地主栽对照品种相比,转基因高产棉花及其受体材料具有营养生长的竞争优势。
2.3.3 生殖生长竞争力 由表4可见,至8月底,撒播和深播2 种方式下,3 个品种均长出果枝和花蕾;但转基因高产棉花及其受体材料的果枝数和花蕾数要多于对照品种中棉所79,但三者间差异不显著;此时除地表撒播方式下常规品种未见棉铃外,其余处理均见棉铃。至9月底,撒播方式下,转基因高产棉及其受体材料的棉铃数均有所增多,对照品种部分植株也长出棉铃。上述结果表明,5月底撒播和深播2 种方式下,转基因高产棉与其受体材料和当地主栽品种相比,生殖生长的竞争优势不明显,在不进行任何栽培管理的环境下,无变为杂草的可能性。
3 讨论
植物的“杂草化”是指自然生长或人工栽培的植物,在新的人工生境中自然繁殖其种群并表现一定的生存竞争优势,从而转变成杂草的演化过程。转基因作物因通过基因工程手段获得新性状,其环境适应能力和生存竞争力可能发生改变,从而增加了作物演化成杂草的风险(钱迎倩和马克平,1998;强胜等,2010)。评估转基因作物和相应非转基因对照品种的生存竞争能力的强弱,如生长发育和繁殖能力、传播方式和传播能力、休眠期及生态适应性等指标,是判断植物是否具有杂草化潜力的主要方法。在相同环境条件下,竞争能力强的植物较易在栖息地占据生存空间,并入侵和改变栖息地环境,最终演变为杂草。
本研究通过比较转基因高产棉花与其受体材料和当地主栽品种在同一荒地环境中的存活、生长和繁殖情况,评价转基因高产棉花的荒地生存竞争能力,进而判断转基因高产棉花的杂草化风险。从试验结果可以看出,荒地4月底和5月底撒播和深播棉种,在不进行任何栽培管理的情况下,与对照品种相比,转基因高产棉花的存活率具有一定的竞争优势。从中后期棉花生长来看,2 种播种方式下转基因高产棉和2 个对照品种的长势均较弱,表现为生长缓慢,植株矮小,生育期延迟,无吐絮铃。因此,不同时期不同播种方式下,转基因高产棉在荒地无竞争优势,不能繁殖传代,不存在杂草化风险。刘娜,李葱葱,徐文静,董英山.2006.转基因作物环境安全性研究进展.分子植物育种,4(1):9-14.马小艳,马艳,彭军,姜伟丽,马亚杰.2013.转基因抗草甘膦抗虫棉的荒地生存竞争能力.生物安全学报,22(2):103-108.钱迎倩,马克平.1998.经遗传修饰生物体的研究进展及其释放后对环境的影响.生态学报,18(1):1-9.强胜,宋小玲,戴伟民.2010.抗除草剂转基因作物面临的机遇与挑战及其发展策略.农业生物技术学报,18(1):114-125.宋新元,张欣芳,于壮,李新海,张明.2011.转基因植物环境安全评价策略.生物安全学报,20(1):37-42.孙凡.2010.水稻OsPDCD5 基因的功能研究和应用及油菜csRRM2 结构域在棉花中的功能鉴定.杭州:浙江大学.王振宇,蒋媛媛,马奇祥,崔小伟,娄永尚,赵辉.2010.转抗病基因棉花荒地生存竞争能力研究.河南农业科学,(10):51-52,120.张兴华,田绍仁,乔艳艳,李捷,杨兆光.2011.转双价抗病基因棉花在荒地生存的竞争能力研究.江西棉花,33(1):42-50.张兴华,田绍仁,张天玉,李捷,乔艳艳.2012.转cry1Ac+cry2Ab 基因棉与转cry1Ac+EPSPS 基因棉荒地的生存竞争能力.生物安全学报,21(2):119-124.中华人民共和国农业部.2007.农业部953 号公告—12.2—2007(转基因植物及其产品环境安全检测 抗虫棉花第2 部 分:生 存 竞争 能力).http://www.moa.gov.cn/zwllm/tzgg/gg/200801/t20080110_952066.htm.
Andow D A and Zwahlen C.2006.Assessing environmental risks of transgenic plants.Ecology Letters,9:196-214.
Hong F,Attia K,Wei C,Li K,He G,Su W,Zhang Q,Qian X and Yang J.2007.Overexpression of the rFCA RNA recognition motif affects morphologies modifications in rice(Oryza sativa L.).Bioscience Reports,27:225-234.
James K.1996.Could transgenic super crops one day breed super weeds.Science,274:180-181.
Sun F,Liu C,Zhang C,Qi W,Zhang X,Wu Z,Kong D,Wang Q,Shang H,Qian X,Li F and Yang J.2012.A conserved RNA recognition motif (RRM)domain of Brassica napus FCA improves cotton fiber quality and yield by regulating cell size.Molecular Breeding,30:93-101.
猜你喜欢
东方少年(2022年27期)2022-11-24
中国棉花(2022年6期)2022-08-16
农业科技通讯(2022年6期)2022-06-21
新疆农业科学(2020年2期)2020-03-12
农药科学与管理(2019年9期)2019-11-23
中国诗歌(2018年6期)2018-11-14
——九江市浔阳区第一幼儿园发展略记
教师博览(2017年2期)2017-04-10
湖南农业(2016年3期)2016-06-05
中国果业信息(2013年6期)2013-01-22
北方作家(2012年5期)2012-12-18
