
五种彩叶植物光合日变化研究
2013-11-18 07:33:10邵怡若郭淑红杨振意
湖南林业科技 2013年1期
张 柔, 邵怡若, 薛 立, 郭淑红, 杨振意
(华南农业大学林学院, 广东 广州 510642)
五种彩叶植物光合日变化研究
张 柔, 邵怡若, 薛 立*, 郭淑红, 杨振意
(华南农业大学林学院, 广东 广州 510642)
采用美国生产的LCI便携式植物光合测定仪测定自然生长的红龙草、黄金榕、变叶木、红背桂和鹅掌柴等5种彩叶植物的净光合速率(Pn)、气孔导度(Gs) 、细胞间隙CO2浓度(Ci)、蒸腾速率(Tr)和气孔限制值(Ls)等光合特征值的日变化。结果表明:红龙草、黄金榕和变叶木的Pn、Gs和Tr日变化曲线呈“单峰”型,其Pn、Gs和Tr在10:00达到高峰;红背桂和鹅掌柴的Pn、Gs和Tr日变化曲线呈“双峰”型,其Pn、Gs和Tr分别在10:00和16:00达到高峰,且第1个高峰值大于第2个高峰值。5 种彩叶植物的Ci日变化均先降后升,红龙草、黄金榕和变叶木的Ci在14:00出现低谷,红背桂和鹅掌柴的Ci在10:00出现低谷。5 种彩叶植物的Ls日变化均先升后降,红龙草和变叶木的Ls在14:00达最高值,黄金榕的Ls在12:00达最高值,红背桂和鹅掌柴的Ls在10:00达最高值。
彩叶植物; 光和特性; 日变化
彩叶植物是指叶色与自然绿色有明显区别,并且具有良好观赏价值植物的总称[1],具有色彩鲜艳、观赏期长的特点,有着观花植物无可比拟的优越性。彩叶植物易于形成大色块景观,可以增加城市的色彩,在现代园林中有重要的地位[2-3]。近年来,随着人们审美水平的提高,在我国一些大中城市开始尝试应用彩叶植物营造景观[4-8]。光合作用是植物最基本的生命活动,是将太阳能转换为化学能的过程,是植物内最重要的化学反应,也是地球上规模最大的生物合成过程[9-10],国内已有较多的报道[11-13]。红龙草 (Altemantheraficoideacv.ruliginosa)、黄金榕(Ficusmicrocarpa)、变叶木(Codiaeumvariegatumvar.pictum)、红背桂(Excoecariacochinchinensis)和鹅掌柴(Scheffleraoctophylla)是我国南方常用的彩叶植物,具有生长速度快、适应性强的特性,广泛栽培于校园、广场、花台、庭园、游乐区以及高楼大厦中,以美化环境[14-18]。然而有关其生态生理学方面的研究,特别是光合特性的研究鲜有报道[14-19]。我们对这5 种彩叶植物的光合日变化特性进行研究,以期为彩叶植物的应用提供参考。
1 材料与方法
1.1试验地概况
试验地设在华南农业大学林学院。该地区位于113°18′E,20°06′N,其气候属于南亚热带典型的季风海洋气候,具有温暖多雨、光热充足、温差较小、夏长冬短等气候特征,年平均气温、最冷月(1月)和最热月平均气温分别为21.8 ℃、13.3 ℃和28.1 ℃。年降雨量1714.4 mm,4—9月降雨量占年降雨量的82%,年均相对湿度79%[20]。
1.2试验材料
供试材料为红龙草、黄金榕、变叶木、红背桂和鹅掌柴,均于2010年春季栽植,生长环境和日常管理均一致,生长良好,无病虫害。
1.3光合特性日变化的测定方法
2012年10月2日,采用美国LI-COR公司生产的Li-6400便携式光合系统分析仪,选取生长状况相近的5种彩叶植物苗木各1株,每1株选取中部外围的5片功能叶进行测定。测定当天天气晴朗,测定时间为08:00—18:00,每2h测定1次,每1片叶重复测3次,取平均值。测定指标包括:净光合速率(Pn)、气孔导度(Gs) 、细胞间隙CO2浓度(Ci)、蒸腾速率(Tr)和气孔限制值(Ls)。气孔限制值[21](Ls) = 1-Ci/Ca(Ca为大气中CO2浓度)。
1.4数据处理
用Excel 2005和SAS9.0统计软件进行数据处理和制图。
2 结果与分析
2.15种彩叶植物净光合速率的日变化
净光合速率(Pn)是光合系统功能的直接体现,也是植株光合系统工作正常与否的指标[22]。5种彩叶植物Pn的日变化曲线如图1所示。红龙草、黄金榕和变叶木的Pn日变化曲线呈“单峰”型,红背桂和鹅掌柴的呈“双峰”型。从08:00—10:00,5种植物的Pn呈快速上升趋势,且都达到了显著差异(P<0.05),其中,变叶木的增幅最大,达527%;鹅掌柴的增幅最少,仅66%。红龙草、黄金榕和变叶木的Pn在10:00左右,出现1天中的最高峰,峰值分别为9.06、9.43 和4.49μmol/(m2·s);10:00过后呈下降趋势,且没有出现明显的“午休现象”。红背桂和鹅掌柴的Pn在10:00左右出现1天中第1个高峰,峰值分别为4.64和4.81μmol/(m2·s);14:00左右出现第2个高峰,峰值分别为3.45和3.11μmol/(m2·s),且第2个峰值明显低于第1个峰值(P<0.001) ;在12:00左右出现低谷,表现出明显的“午休现象”,14:00之后均呈下降趋势。5 种彩叶植物的Pn在18:00左右达到1天中的最低值,且为负值,说明此时植物的光合速率低于呼吸速率。通过比较净光合速率的日均值发现,5 种彩叶植物的光合作用强弱顺序为: 红龙草> 黄金榕>鹅掌柴> 红背桂> 变叶木。

图1 5种彩叶植物净光合速率的日变化Fig.1 Diurnal variation of net photosynthetic rate of the five colorful plant species
2.25种彩叶植物气孔导度(Gs)的日变化
植物通过改变气孔开度来控制其与外界CO2和水汽的交换,从而调节自身的Pn、Tr和呼吸等重要的生理过程,以适应环境。5 种彩叶植物气孔导度(Gs)的日变化规律(见图2)与其净光合速率(Pn)的日变化规律相似。红龙草、黄金榕和变叶木的Gs日变化曲线呈单峰型,红背桂和鹅掌柴的呈双峰型。1天之中,随着气温与光照强度的增强,叶片的气孔相应扩大,Gs也不断增大。红龙草、黄金榕和变叶木的Gs在10:00左右出现1天中的最高峰,峰值分别为0.14、0.18和0.06mol/(m2·s);10:00过后呈下降趋势。红背桂和鹅掌柴的Gs在10:00左右出现1天中第1个高峰,峰值分别为0.06和0.08mol/(m2·s),随着外界温度进一步升高,受高温的影响,气孔逐渐闭合,气孔导度相应降低,在12:00左右达到低谷, 这与植物光合“午休”时间相一致;12:00(高温)之后,外界温度逐渐降低,叶片气孔又重新张开,气孔导度又开始升高,于14:00左右出现第2个高峰,峰值分别为0.04和0.05mol/(m2·s);14:00之后均呈下降趋势。
2.35种彩叶植物细胞间隙CO2浓度的日变化
细胞间隙CO2浓度(Ci)是影响植物光合作用的一个重要因素,能控制光合作用的直接合成碳源。 5 种彩叶植物Ci的日变化规律(见图3)与其净光合速率(Pn)和气孔导度(Gs)的日变化规律大致相反。5 种彩叶植物的Ci均先降低后升高,红龙草、黄金榕和变叶木的Ci在14:00左右达到最低值,分别为183.56、200和172.16μmol/mol,红背桂和鹅掌的Ci在10:00左右达到最低值,分别为195.79和206.2μmol/mol。
2.45种彩叶植物蒸腾速率的日变化
蒸腾作用是植物对水分吸收和运输的一个主要动力,能促进植物对矿物质的吸收和运输,以保证叶肉细胞光合作用的原料供给。气孔蒸腾是植物进行蒸腾作用的主要方式。如图4所示,5 种彩叶植物的蒸腾速率(Tr)的日变化曲线与其净光合速率(Pn)和气孔导度(Gs)的日变化曲线相似。红龙草、黄金榕、变叶木和红背桂的Tr日变化曲线均呈“单峰”型,先升后降;鹅掌柴的呈“双峰”型,为先升后降,然后再一次升降。这一结果与蒸腾速率主要受叶片气孔调节有关。

图2 5种彩叶植物气孔导度的日变化 Fig.2 Diurnal variation of stomatal conductance of the five colorful plant species

图3 5种彩叶植物细胞间隙CO2浓度的日变化Fig.3 Diurnal variation of intercellular CO2 concentration of the five colorful plant species

图4 5种彩叶植物蒸腾速率的日变化Fig.4 Diurnal variation of transpiration rate of the five colorful plant species
2.55种彩叶植物气孔限制值的日变化
气孔限制值(Ls)在一定程度上反映了植物叶片对大气CO2相对利用效率的大小,与细胞间隙CO2浓度密切相关。5 种彩叶植物的Ls日变化(见图5)表现出与细胞间隙CO2浓度(Ci)相反的规律,均呈先升后降。红龙草、黄金榕和变叶木的Ls在14:00时左右达到最大,而此时其Ci则处于1天中的最低值;红背桂和鹅掌柴的Ls在10:00左右达到最大,而此时其细胞间隙CO2浓度最小。
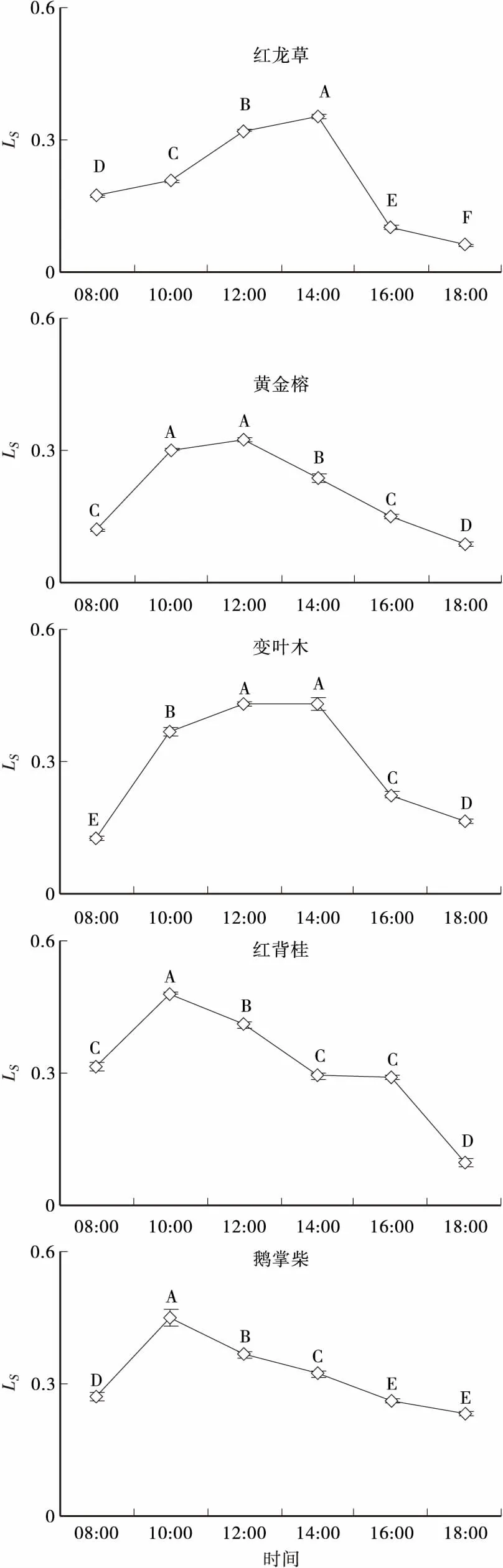
图5 5种彩叶植物气孔限制值的日变化Fig.5 Diurnal variation of leaf Ls of the five colorful plant species
3 结论与讨论
(1)红龙草、黄金榕和变叶木的净光合速率(Pn)日变化曲线均呈“单峰”,Pn在10:00左右出现1天中的最高峰之后呈下降趋势;红背桂和鹅掌柴的Pn日变化曲线呈“双峰”,并且这2种植物表现出明显的光合“午休”现象。这与刘弘等[23]对紫叶李(Prunuscerasiferacv.atropurpurea) 和李晓征等[24]对金叶含笑(Micheliafoveolata) 的研究结果相一致。主要原因是强光、高温、低湿和土壤干旱等条件引起气孔部分关闭和光合作用光抑制。植物光合“午休”现象是由于部分气孔关闭引起的气孔限制和叶肉细胞自身活性下降导致非气孔限制2种原因引起的[25-27]。红龙草、黄金榕和变叶木的Pn在10:00之后下降,在10:00—14:00,其细胞间隙CO2浓度(Ci)下降,气孔限制值(Ls)增加,此时的下降是由气孔限制引起的;在14:00之后Ci上升,Ls下降,此时的下降是由非气孔限制引起的。
(2) 红龙草、黄金榕、变叶木、红背桂和鹅掌柴等5 种彩叶植物的蒸腾速率(Tr)与气孔导度(Gs)日变化规律基本一致,其净光合速率和蒸腾速率均与气孔导度有一定的相关性。蒸腾作用既受外界因子的影响,也受植物体内部结构和生理状况的调节,是一个复杂的生理过程。蒸腾作用的正常进行,气孔开放,有利于光合作用中CO2固定。光照是影响蒸腾作用的最主要的外界条件,光对蒸腾的影响首先是引起气孔开放,其次是提高大气和植物体的温度,增加叶内外蒸气压而加速蒸腾。
[1] 袁涛. 彩叶植物漫谈[J].植物杂志,2001(5):12-13.
[2] 于晓南,张启翔. 彩叶植物多彩形成的研究进展[J].园艺学报,2000,27(增刊):533-538.
[3] 徐华金,张志毅,王莹. 彩叶植物研究开发现状及展望[J].四川林业科技,2007,28(1):44-49.
[4] 孔令玉,任景景,余佳,等. 彩叶植物在海南园林中的应用[J].热带农业科学,2012,32(3):60-64.
[5] 张学鑫. 彩叶植物在深圳园林绿化中的应用[J].现代农业科技,2010(12):214-216.
[6] 朱素平,徐杰元. 彩叶植物在城市园林绿化中的应用[J].现代农业科技,2011(14):269-270.
[7] 李玉萍,李宏,夏和宝. 彩叶植物资源及其在南京园林中的应用[J].金陵科技学院学报,2006,22(1):95-100.
[8] 刘维华,李梅. 彩叶植物的观赏价值及应用前景探讨[J].四川林勘设计,2005(2):25-27.
[9] 沈允刚. 地球上最重要的化学反应——光合作用[M].北京:清华大学出版社,2000.
[10] 沈允钢, 施教耐. 动态光合作用[M]. 北京:科学出版社,1998:154-174.
[11] 郭淑红,薛立, 张柔,等. 4种幼苗对低温胁迫的光合生理响应[J].华南农业大学学报, 2012,33(3): 373-377.
[12] 杨振意,薛立,张柔,等. 干旱对南五味子和海南红豆幼苗气体交换参数的影响[J]. 湖南林业科技, 2012, 39(1):21-23.
[13] 张柔,薛立,郭淑红,等. 南五味子和海南红豆对淹水胁迫的光合生理响应[J].湖南林业科技,2012,39(1):24-26.
[14] 廖友媛. 红背桂光合特性日变化研究湖南工业大学学报[J].2008,22(6):36-38.
[15] 权宏,施和平. 红龙草叶片的组织培养及其植株再生[J].园艺学报,2005,32 (4):735-737.
[16] 权宏. 红龙草的组织培养及发根农杆菌对红龙草的遗传转化研究[D]. 广州:华南师范大学,2004.
[17] 林钧. 黄金榕扦插繁殖技术研究[J].青海农林科技,2012(1):12-14.
[18] 王育选,于娜. 变叶木组培快繁技术研究现代农业科学[J].2008,15(1):34-36.
[19] 刘嘉君,王志刚,刘炳响,等. 四种彩叶树种光合特性研究安徽农业科学[J].2011,39(8): 4967-4970.
[20] 薛立,张柔,奚如春,等. 华南地区6 种阔叶幼苗叶片形态特征的季节变化[J].生态学报,2012,32(1): 123-134.
[21] Berry J , Bjorkman O. Photosynthetic response and adaptation to temperature in higher plants[J]. Ann. Rev. Plant Physiol. , 1980,31:491-543.
[22] 周建,杨立峰,郝峰鸽,等. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报, 2009, 29(1):0136-0142.
[23] 刘弘,马杰,李保印. 三种彩叶植物的光合特性日变化研究[J].河南职业技术师范学院学报, 2004, 32(2): 33-35.
[24] 李晓征,彭峰,徐迎春,等. 不同遮荫下多脉青冈和金叶含笑幼苗叶片的气体交换日变化[J] . 浙江林学院学报, 2005, 22 (4): 380-384.
[25] 许大全. 光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[26] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[27] Farquhar G D , Sharkey T D. Stomatal conductance and photosynthesis[J]. Ann. Rev. Plant Physiol., 1982, 33: 317-345.
(文字编校:唐效蓉)
Diurnalchangesofphotosyntheticcharacteristicsoffivecolorfulplantspecies
ZHANG Rou, SHAO Yiruo, XUE Li*, GUO Shuhong, YANG Zhenyi
(College of Forestry, South China Agricultural University, Guangzhou 510642, China)
Diurnal changes of net photosynthetic rates, stomatal conductance, intercellular carbon dioxide concentration, transpiration rates and stomatal limits of photosynthetic characteristics ofAltemantheraficoideacv.ruliginosa,Ficusmicrocarpa,Codiaeumvariegatumvar.pictum,ExcoecariacochinchinensisandScheffleraoctophyllawere studied under natural conditions by using LCI portable photosynthesis system. The results showed that, the diurnal changes ofPn,GsandTrofA.ficoideacv.ruliginosa,F.microcarpaandC.variegatumvar.pictumemerged a single-peak curves with the peak at 10:00, whereas those ofE.cochinchinensisandS.octophyllaexhibited double-peaks curves with the first peak at 10:00 and the second peak at 16:00, and the first peak was higher than the second peak. The diurnal changes ofCsof five colorful plants decreased followed by an increase. The minimumCiofA.ficoideacv.Ruliginosa,F.microcarpaandC.variegatumvar.pictumoccurred at 14:00 while that ofE.cochinchinensisandS.octophyllaappeared at 10:00.The diurnal changes ofLsof five colorful plants increased followed by an decrease, which reached maximum at 14:00 forA.ficoideacv.ruliginosaandC.variegatumvar.pictum, at 10:00 forE.cochinchinensisandS.octophyllaand at 12:00 forF.microcarpa, respectively.
colorful plants; photosynthetic characteristics; diurnal changes
2012-10-10
2013-01-16
广东省林业局资助项目(F09054)。
张 柔(1986-),女,山东省荷泽市人,硕士生,主要从事森林培育和树木生理学研究。
* 为通迅作者
Q 945
A
1003-5710(2013)01-0008-06
10. 3969/j. issn. 1003-5710. 2013. 01. 003
猜你喜欢
剑南文学(2022年1期)2022-10-22 10:35:50
意林(2021年19期)2021-10-27 14:22:57
食品安全导刊(2021年21期)2021-08-30 08:22:08
农民致富之友(2020年14期)2020-05-25 02:29:40
故事作文·高年级(2019年8期)2019-08-20 17:53:42
现代园艺(2017年22期)2018-01-19 05:07:24
现代园艺(2017年23期)2018-01-18 06:58:08
信息记录材料(2016年4期)2016-03-11 15:22:57
作文周刊(中考版)(2009年40期)2009-08-24 08:02:42
烹调知识(2004年6期)2004-01-05 09:27:00
