
5 种壳斗科树种实生苗年生长规律研究
2013-11-05 06:39叶金山
江西农业大学学报 2013年3期
余 林,叶金山,朱 恒,周 诚*
(1.江西省林业科学院,江西 南昌330032;2.上饶市林业科学研究所,江西 上饶334000)
江西省的地带性植被是亚热带常绿阔叶林。由于长期的人为干扰,许多林分转变为针叶纯林,而研究表明针叶纯林长期经营存在退化的问题[1-2]。进入1980 年代后,江西省开始重视营造阔叶林,对阔叶树种培育的研究也加大了力度[3-4]。甜槠、石栎、麻栎、青冈和碟斗青冈等壳斗科树种作为江西省常绿阔叶林的重要组成树种,已被应用于改土和绿化造林。但是壳斗科树种苗期生长较缓慢,且缺乏成熟、系统的壮苗培育技术,这将影响其大面积人工栽培[5]。苗木的生长节律研究是森林培育的研究重点之一[6-19],掌握苗木的生长节律,确定其生长盛期及生长速度等,可以更好地科学培育、管理苗木,提高其经济、生态价值。
由于苗木的生长量是连续的,采用有序样本聚类法对1 a 生苗木生长期进行定量划分是一种科学、客观的方法,已被广泛应用[6-9]。以往研究表明,Logistic 模型能够较好地拟合植物生长过程,在苗木生长节律研究中也得到广泛应用[10,14]。因此,本文在定期观测甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗高年生长过程的基础上,运用有序样本聚类法和Logistic 模型定量分析了苗木的生长规律,旨在为扩大繁殖、培育良种壮苗提供理论依据和实践指导。
1 材料与方法
1.1 试验地概况
试验地设在江西省南昌市江西省林业科学院苗圃(E 28°44'34″,N 115°48'55″)。地处江西省中部偏北,属中亚热带湿润季风气候,年平均气温17.5 ℃,极端最高气温40.6 ℃,极端最低气温-9.3 ℃,年平均降水量1 754.9 mm,平均无霜期294 d,日照率43%。试验地地势平坦,排灌条件良好,圃地周围开阔,日照充足,土壤为红壤,肥力中等。
1.2 供试材料
试验所用种子于2006 年采自江西九连山自然保护区成片健壮的母树林。采种后先将受病虫损害及颜色不正常的种子剔除,然后浸入55 ℃温水中10 ~15 min,以杀死种子内害虫。经杀虫处理后的种子摊在阴凉干燥处,每天翻动2 ~3 次,晾干后沙藏。沙藏期间定期检查沙子湿度及种子是否发热发霉情况。
1.3 播种及苗期管理
2006 年冬季进行苗圃整地,做到三犁三耙,作业深度在20 cm 以上。按宽1.5 m、长10 m、高20 cm作床,苗床中央稍隆起以利于排水。2007 年2 月中旬用质量分数0.5%高锰酸钾和400 g/cm3的辛硫酸乳液喷洒床面进行土壤消毒。2007 年3 月上旬将沙藏的种子取出按行距30 cm,沟深8 ~10 cm 进行条播,播后覆土3 ~5 cm,以稻草覆盖,厚1 ~2 cm。苗期按常规管理[20-21]。
1.4 试验设计及调查方法
自苗木出齐后,每个树种选择生长健壮的小苗30 株为1 个小区,设置3 次重复,于每月中旬定期逐株测量苗高,至2007 年12 月15 日结束。
1.5 数据处理方法
应用SPSS 软件对本研究中5 种壳斗科树种1 a 生实生苗的苗高进行有序样本聚类,综合分析各树种实生苗生长情况,进而进行生长时期的划分。
本研究采用Logistic 模型模拟5 种壳斗科树种实生苗的苗高年生长动态,该模型的表达式为:

(1)式中:H 为苗高累积生长量(cm);t 为生长时间(d);K 为苗高年生长极限值(cm);a、b 为方程参数。用SPSS 软件对模型中参数K、a、b 进行估计。
依据文献[15]中方法,进行各树种苗高连日生长量的拟合:

(2)式中:tmax为苗高日生长量最大值的时间(d),称为速生点;t1和t2为苗高日生长量变化最大的2个值的时间(d),当t1<t <t2时称为苗木生长盛期。
2 结果与分析
2.1 苗高年生长节律
从出苗基本整齐起,根据定期观测的数据将生长期内各树种苗高的月净生长量结果绘于图1。
从图1 可以看出,各树种1 a 生实生苗的高生长动态均呈现“慢—快—慢”的生长节律。甜槠、石栎、麻栎、青冈和碟斗青冈实生苗的苗高年生长量平均值分别为12. 37、26. 69、61. 11、35.00 和43.76 cm。苗高生长高峰集中在6 月至10 月,但各树种间存在差异,麻栎和碟斗青冈在7 月份达到苗高生长高峰,苗高月净生长量为10.73 cm和7. 38 cm,各占生长期高生长量的17.55%和16.86%;甜槠、石栎和青冈苗高月净生长量出现在8 月,分别为2.98、4.65 和5.31 cm,各占生长期高生长量的24.05%、17.44%和15.17%。
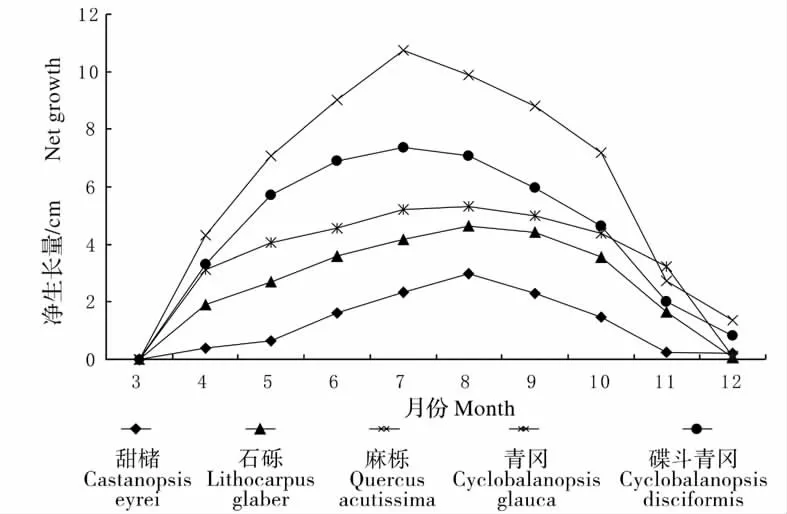
图1 5 种壳斗科树种苗高生长月净生长量Fig.1 Height net growth of five species of beech family

表1 5 种壳斗科树种苗高生长时期划分及生长情况比较Tab.1 Division and comparison of growth periods of five species of beech family
2.2 苗高各生长时期的划分
以甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗的苗高月净生长量为样本,利用有序样本聚类法进行苗高各生长时期的划分,将苗期划分为生长初期、生长盛期及生长后期3 个时期,具体见表1。
从表1 可以看出,5 种壳斗科树种苗高生长初期、生长盛期和生长后期各有不同。5 种壳斗科树种苗高生长初期在32 ~70 d,占整个生长期的14.88% ~25.36%,生长后期在46 ~94 d,占整个生长期的18.78% ~43.72%。各树种的生长盛期分别为:甜槠为6 月16 日至9 月12 日,石栎为5 月25 日至10月30 日,麻栎为5 月30 日至10 月25 日,青冈为6 月16 日至10 月15 日,碟斗青冈为5 月24 日至9 月24 日。麻栎在生长盛期苗高净生长量最大,达到41.71 cm,是甜槠生长盛期苗高净生长量的5.12 倍。各树种生长盛期占整个生长期的比率在41.40% ~64.90%,生长盛期苗高净生长量占总生长量的比率在55.10% ~68.25%。
2.3 苗高生长过程拟合
利用Logistic 模型拟合各树种苗高生长量随生长时间的变化规律,拟合得出的方程及检验结果见表2。
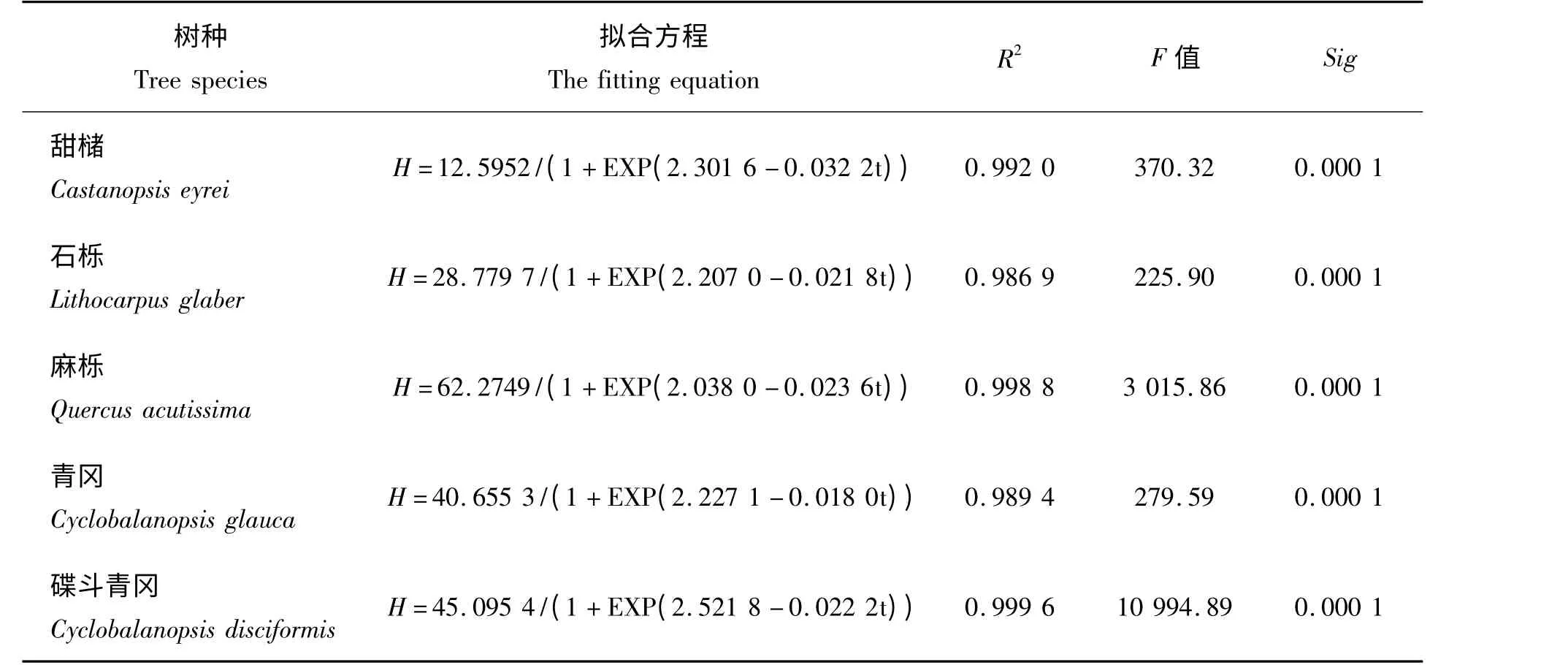
表2 5 种壳斗科树种苗高连续生长量的Logistic 方程Tab.2 The height growth simulated by Logistic model of five species of beech family
由表2 可见,各树种苗高连续生长量的Logistic 方程决定系数均大于0.98,其中碟斗青冈苗高连续生长量的Logistic 方程回归系数最大,达到0.999 6,说明苗高生长量与生长时间相关性显著。用F 值进行显著性检验,均达到极显著水平(Sig=0.000 1 <0.01),说明回归结果可靠,利用Logistic 模型可以反映出甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗的苗高生长过程。
2.4 苗高连日生长量拟合
甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗的苗高连日生长量变化情况见表3。
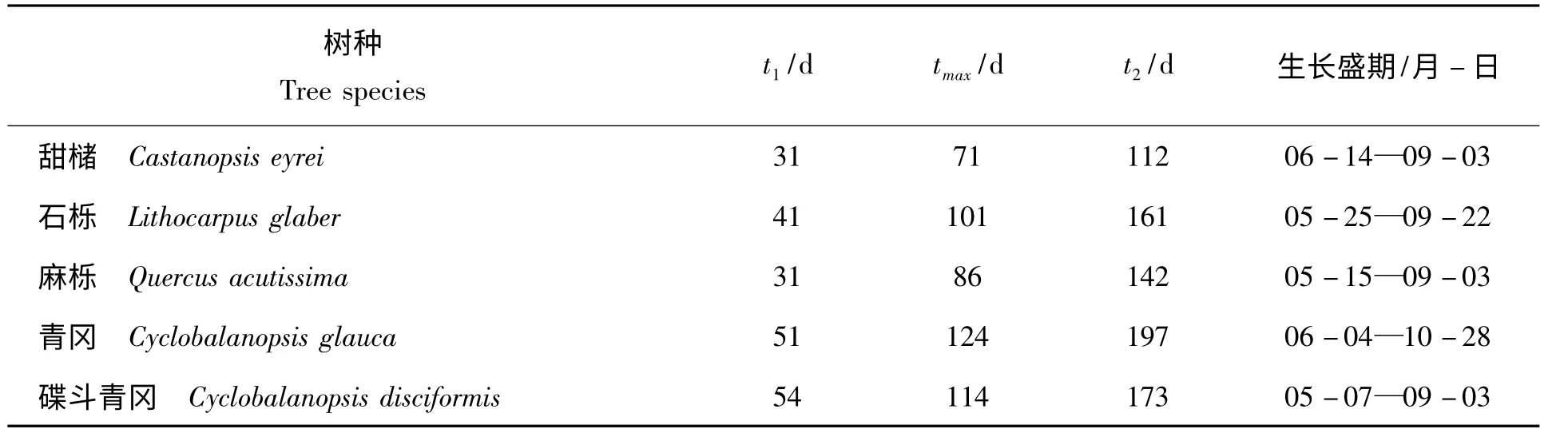
表3 5 种壳斗科树种连日生长量变化情况Tab.3 The height growth day increment by Logistic model of five species of beech family
表3 表明,5 种壳斗科树种1 a 生实生苗的苗高速生点(tmax)差异较大,最早的甜槠(71 d)与最晚的青冈(124 d)相差53 d。各树种苗高连日生长量的最大变化点差异也较大,t1在31 ~54 d,t2在112~197 d。t1和t2是连日生长量变化速率最快的2 个点,用t1和t2分别作为划分生长初期与生长盛期的分界点和生长盛期与生长后期的分界点,划分出的生长盛期与利用有序样本聚类法分析结果基本一致,进一步说明用Logistic 模型拟合各树种苗高生长过程是合适的。各树种生长盛期持续时间以青冈最长,达到146 d,其次为石栎,为120 d,最短的是甜槠,与青冈相差65 d。
3 结论与讨论
甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗的苗高年生长量平均值分别为12.37、26.69、61.11、35.00 和43.76 cm,与以往关于其他壳斗科树种苗高生长的研究结果存在差异[16,17,21],这可能是因为实生苗的生长状况除受到其自身的遗传因素影响外,还与种源地及播种地的地理位置和气候因子有一定的关系[18]。各树种1 a 生实生苗的高生长呈现出“慢—快—慢”的节律,生长高峰集中在6 月至10 月,且具有明显的阶段性。利用有序样本聚类法能够较好地将各树种的苗高生长划分为生产初期、生长盛期和生长后期3 个生长时期。
利用Logistic 方程拟合了5 种壳斗科树种1 a 生实生苗苗高生长量与生长时间的相关关系,方程决定系数均在0.98 以上,经检验均达到极显著水平。说明Logistic 方程可以反映出甜槠、石栎、麻栎、青冈和碟斗青冈1 a 生实生苗的苗高生长过程。对Logistic 曲线进行求导所确定的速生点和速生期,在苗期管理上具有重要意义[19]。本研究中5 种壳斗科树种间苗高的速生点、生长拐点和生长盛期持续时间差异较大,甜槠在71 d 达到日生长高峰,而青冈在124 d 达到日生长最大值,两者相差53 d。
[1]盛炜彤,范少辉.杉木人工林长期生产力保持机制研究[M].北京:科学出版社,2005.
[2]莫江明.鼎湖山马尾松林退化机理和恢复的生态学特征[D].广州:中国科学院华南植物园,2003.
[3]肖复明,黄晓春,曾志光.浅析江西阔叶树育种策略[J].江西林业科技,2003,30(6):23 -25.
[4]刘光正,曹展波,肖水清,等.江西9 个优良阔叶树种栽培试验[J].林业科技开发,2000,14(4):38 -40.
[5]黎建军,冯绍龙,刘鲁平,等.阔叶树种的研究现状与江西研究的发展方向[J].江西林业科技,2001(4):35 -38.
[6]彭方仁.有序聚类法在大叶樟苗高生长时期划分中的应用[J].林业科学研究,1989,2(5):501 -504.
[7]曹帮华.有序聚类和数学模拟法在刺槐苗年生长规律研究中的应用[J].山东农业大学学报,1998,29(4):487 -494.
[8]陈存及,李生,曹永慧,等.光皮桦苗高生长时期划分有序样本聚类分析[J].福建林学院学报,2002,22(3):197-200.
[9]林永英.卷斗青冈苗高生长时期有序聚类划分方法研究[J].福建林学院学报,2002,22(3):223 -226.
[10]郑仁华,杨宗武,施季森,等.福建柏优树子代苗期性状遗传变异和生长节律研究[J].林业科学,2003,39(专刊1):179 -183.
[11]郑益兴,彭兴民,赵保荣,等.印楝实生苗年生长规律研究[J].林业科学研究,2006,19(2):182 -187.
[12]郑益兴,冯永刚,彭兴民,等.印楝1 年生苗木生长节律与数量分级标准[J].南京林业大学学报:自然科学版,2008,32(3):25 -30.
[13]余道平,彭启新,李策宏,等.峨眉四照花育苗技术及苗高年生长规律研究[J].西北林学院学报,2008,23(1):111-114.
[14]吴国欣,王凌晖,俞建妹,等.降香黄檀幼苗年生长节律研究[J].浙江林业科技,2010,30(3):56 -60.
[15]杨斌.柳树苗期年生长模型的研究[J].西北林学院学报,2006,21(6):97 -99.
[16]游水生,王海为,廖祖辉.锥栗1 年生播种苗的年生长规律[J].福建林学院学报,2003,23(2):102 -105.
[17]万娜娜,吴晟,邹元熹,等.井冈山槠等阔叶树播种苗年生长规律研究[J].林业科技开发,2005,19(1):34 -37.
[18]赖文胜.长序榆一年生播种苗的年生长规律[J].南京林业大学学报,2001,25(4):57 -60.
[19]徐宏远,陈章水.不同密度I-69 杨树生长规律的研究[J].林业科学研究,1994,7(1):61 -66.
[20]罗坤水,杨春霞,林小凡,等.壳斗科树种育苗技术研究[J].江西林业科技,2008,35(6):6 -9.
[21]易观路,许方宏,罗建华,等.壳斗科7 种树种育苗及造林效果初步观测[J].贵州林业科技,2004,32(1):31 -34.
猜你喜欢
广东蚕业(2022年4期)2022-11-23
农业科技通讯(2022年1期)2022-01-17
生态学报(2021年6期)2021-04-25
湖北农机化(2020年16期)2020-10-21
江西农业(2020年13期)2020-09-08
山东林业科技(2019年2期)2019-06-03
——以新源县为例
新疆农业科技(2018年5期)2018-05-07
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
中国糖料(2016年1期)2016-12-01
