
生物受激发光的管流实验研究
2013-10-21 00:49:48吴荣华马治国宗思光王江安
发光学报 2013年10期
曹 静,吴荣华,马治国,宗思光,王江安
(海军工程大学电子工程学院,湖北 武汉 430033)
1 引 言
生物受激发光是海洋生物体受流场刺激而产生的发光现象。在破碎的波浪[1]、舰船的尾流[2]、运动生物[3]的海域等很多情况下都可以观测到这类生物发光,而且当海洋中生物体浓度超过100 个/L 时,生物体的发光强度就足以能够照亮周围运动的物体。生物光尾流是舰船等运动物体在经过富含发光生物体的海域时在其周围以及尾部由于流场的剧烈扰动而使生物体发光、形成一条发光带的现象,因此通过对生物光尾流的检测就可实现对水中目标的探测及跟踪。生物受激发光与生物光尾流的研究涉及海洋生物学、海洋光学、流体力学、地球物理学等多学科。
海洋中约有80%以上的生物都会发光,受激发光生物体的种类也很繁多,其中最典型的受激发光生物体是腰鞭毛虫。本文选取了在海洋中分布广泛且能在实验室培养的3 种腰鞭毛虫(多边膝沟藻、夜光虫、月牙形梨甲藻),通过自主设计的管流实验装置研究了流场的刺激对海洋生物发光的影响,分析了流场与生物受激发光之间的相关性,从而为生物光水中探测技术的研究提供了一定的参考。
2 管流场中的水动力特性分析
影响生物发光的因素有许多,例如流体的扰动、油污等化学物质的干扰、电磁信号的干扰、激光信号的干扰等都有可能使发光生物体发光,但水中运动物体刺激生物发光的最主要原因是物体机械运动引起的流场水动力特性变化。为了研究使用生物光尾流探测技术的可行性,就必须研究发光生物受水动力扰动的发光特性与流场刺激的相关性。根据国外已有的研究经验,我们选择了管流实验来具体研究受激发光与流场刺激的相关性。采用管流实验的原因主要有以下5 点[4]:
(1)管流实验中的流场特性很容易通过测量管中的流速和压力差来进行描述;
(2)管流能提供一个宽范围的(有时甚至是重叠在一起的)层流和湍流刺激;
(3)流场中的发光生物体能够持续不断地通过管流装置中的水箱进行补充;
(4)流场中生物的细胞体只会受到流场的短时刺激;
(5)当发光生物进入管子或者逆流运动通过管子时,生物发光的级别都不会出现明显的降低。
因此,通过管流实验的研究能够细致地了解发光生物在不同水动力扰动下的发光特性,为后面的生物光尾流研究打下坚实的实验基础。
2.1 进口段的特性
流体刚进入管内时,整个管截面上的速度U均匀,但管壁上的速度为0。随着流体向前流动,由于流体粘性作用,管壁附近的速度降低,截面上的速度分布不断变化,直至离管口一定距离Le后不再改变。速度分布随轴向距离变化的管段称为进口段。进口段内发展着的流动与绕流时壁面附近的流动具有相似之处。边界层厚度δ 在进口处为0,此后沿管长逐渐增加。在边界层之外是径向速度梯度为0 的区域,这一区域沿流动方向不断缩小,流体的流速不断增加,依次可由U0、U1、U2、U3变至Umax,以保持截面平均速度不变,即流量不变。无粘性流区域消失后,充分发展的速度分布即可形成。通常所说的圆管中的流动就是这种达到平衡状态的流动。当达到平衡状态的流动为层流时,进口段也是层流;当平衡态为湍流时,在进口段从层流向湍流变化,要引起边界层的转捩。进口段长度Le与雷诺数Re 的关系为[5]:
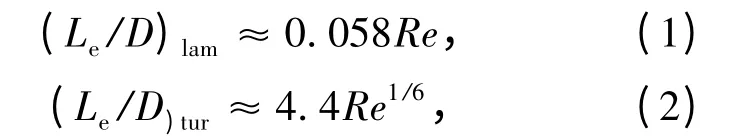
其中D 是圆管的直径,而雷诺数Re 是流体的密度ρ、圆管截面上的平均速度Uavg和流体的动力粘性系数μ 的函数,即

从式(1)和(2)可以看出,若完全发展的流动为层流,Le随Re 线性变化;但是当流场为湍流时,Le随雷诺数Re 的变化就小,这主要是由湍流混合作用的边界层发展过快引起的。
当管流场中的下游没有速度分布时就认为流场达到了平衡。由于没有平均加速度,管流中剪应力是压力差、柱状流场半径的函数,即

那么在管壁R 处其剪应力大小为:

从而流场内任意一点的剪应力与壁剪应力的关系为

不管流场是湍流还是层流,这种剪应力的线性分布都是普遍适用的[5],从而得到管流截面上的平均剪应力为壁剪应力的2/3,即
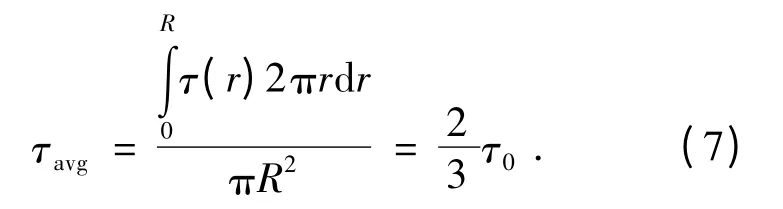
2.2 管流中的层流
当管内流场为层流时,剪应力还可表示为
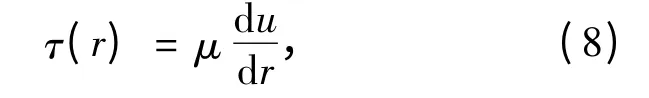
其中u(r)是半径r 处沿着管轴向的速度。
每单位长度L 上的压降量用ΔP(ΔP >0)表示,则管内流动沿程压降的无量纲的阻力系数,也称为达西摩擦系数(λ),其表达式为[5]:
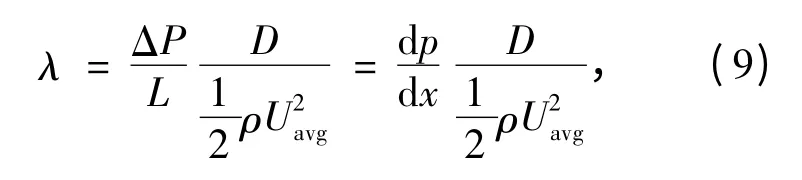
又根据层流的体积流量公式以及与平均速度的关系,可知:

将式(10)和(3)代入式(9),可得层流中的管摩擦系数为:

从式(9)可以看出管摩擦系数与壁面的粗糙程度无关。由式(5)、(9)和(11)可得层流中管壁面的剪应力为:

2.3 管流中的湍流
圆管内的内部湍流不带自由边界,虽然其边界条件的湍流边界层不同,但其流动结构仍然是由与边界层一样直接受壁面影响的壁区域以及由上游来的受积累影响的管中央区域构成,这样就可经常采用与边界层类似的解析方法。在充分发展的管内湍流方面,虽然离开壁面的区域也象边界层湍流那样存在非湍流部分,但没有间歇性,称它为湍流核心区域。


由于公式(13)用纯理论是求解不出来的,在工程应用中可用一种更简便的方法计算平均速度和阻力系数,即幂函数形式的平均速度分布:
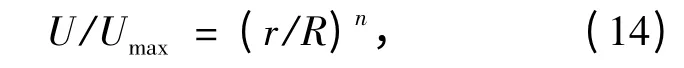
其中指数n 随Re 的变化为

在4 ×103≤Re≤105范围内,对于光滑圆管,布拉修斯给出了λ 的经验公式:
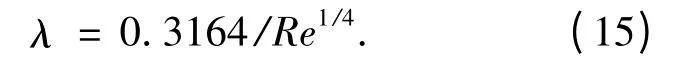
联立式(5)、(9)和(15),从而得到湍流场中的壁剪应力为
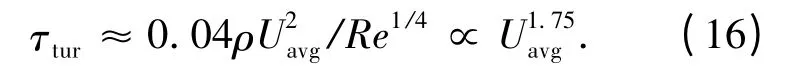
在层流状态,阻力系数随雷诺数的增加而减小。当流场由层流向湍流转变时,阻力系数没有确定规律,表面剪应力随环境因素变化很大。在流动达到完全湍流状态后,阻力系数继续随雷诺数的增大而减小,不过减小得比较缓慢。在充分发展的圆管湍流中,壁面粗糙度对阻力系数的影响较大。在给定雷诺数下,相对粗糙度增加,阻力系数增大;当相对粗糙度增加到一定量后,流动进入完全粗糙状态。在完全粗糙区,阻力系数不随雷诺数变化,变为常数。
3 管流实验装置及方案
管流的实验装置[5]如图1 所示,上端是一个容积25 L 的水箱,它连接到一个收缩形状的喷嘴龙头和一个由光滑铜管以及直径为0.65 cm 的玻璃管制成的出水管,管子尾部装有用来控制流速的流量计和阀门。压力测量值通过装有水银的压差计来显示。生物发光的定量测量采用的是光子计数模式的滨松光电倍增管,放置在管子底部距离入口65 cm 的位置。

图1 管流装置示意图和实物图Fig.1 The equipment of pipe flow experiment
实验均设置在生物体能够发光的黑暗期,且所有的实验都是在晚上九点后的暗室中进行,从而减少了背景光噪声对生物微弱光测量的干扰。为了让发光藻适应黑暗的环境并在测量前不受任何机械扰动,每次实验前需要提前1 h 将含有生物体的海水缓慢注入水箱中静置,同时将实验室的温度控制在22 ℃左右以保证生物体的活性。此外,为了减小生物体浓度不均匀、流场不稳定等其他因素产生的随机误差,同一流速下的生物发光测量一般要持续100~250 s,然后对测量时间内的发光量取平均值,从而得到该流速下单位时间内生物受激发光的光子数。
4 管流实验结果与分析
4.1 管流水动力特性的实验测量
在分析管流不同流速下流场的水动力特性时,用流量计和压差计分别测量流场的流速、压强差两个状态参量。把流速测量值代入式(3)可计算出流场的雷诺数,把压强差测量值代入式(5)可算出壁剪应力值,再将流速、压强差代入公式(9)可得到流场的达西摩擦系数,其实验测量结果与理论值如图2 所示。图2(a)给出了处于不同状态(层流、湍流、完全发展的或过渡期)的流场雷诺数与摩擦系数的关系,其中直线代表理论值,单点代表每个不同流速下的实验测量值。由上节的理论分析可知,当流场处于层流时,雷诺数与摩擦系数满足式(11)所示的线性关系。测量结果表明,当Re<2 200 时,实验结果与理论值吻合得较好,这与Latz 等人1999年在文献[8]和文献[9]中给出的结果相符。而随着雷诺数的增大,流场变得不稳定,此时的流场处于过渡期,测量值会偏离理论值。尽管在染色实验中已经证实,当雷诺数Re=7 000 时,流场还可能处在层流状态,但是其流场是不完全发展的。湍流区的实验测量值与经验公式(15)符合得较好。图2(b)给出了层流时不同平均流速下壁剪应力的测量值与理论值间的关系,从图中可以看出,当Uavg<0.35 m/s时,壁剪应力随流场的平均流速线性增加,并与式(16)的理论值相吻合。当Uavg>0.35 m/s 时,测量值会略微大于理论值,这主要是由高雷诺数层流下压强差的测量值大于理论值引起的[8]。

图2 管流的水动力特性。(a)雷诺数与摩擦系数的关系;(b)层流场中平均流速与壁剪应力的关系。Fig.2 Hydrodynamic characterization of pipe flow.(a)Relationship of Re and the friction coefficient.(b)Relationship of the mean flow speed and the wall shear stress.
4.2 低速管流刺激下的生物受激发光
图3 给出了3 种腰鞭毛虫的浓度约为30 个/mL 时,在流速为45 mL/s(流场壁剪应力τwall=0.21 N/m2)的流场刺激下受激发光量随时间的变化情况,其中光子计数器的积分时间为100 ms。从图中可看出3 种生物体在同条件刺激下的发光次数不同,按从少到多排列依次为多边膝沟藻、夜光虫、月牙形梨甲藻,其中月牙形梨甲藻在120 s 内的发光次数和发光强度都明显高于另外两种。据国外的研究表明,腰鞭毛虫受到刺激后可多次发光,依据种类的不同可从2、3 次到几十次不等,每次发光的持续时间从几毫秒到几百毫秒变化[4]。实验中由于受设备的限制,为了减小误差,采样时间最小只能设置为100 ms,所以测量所得的光子数可能是单个生物体的单次发光,也可能是多次发光。另外,由于实验数据是管流场下的刺激发光,所以进入探测器的光子是由水箱源源不断地流出的未被刺激的新鲜生物体发出的,也就是说,在计数器的一次积分时间内的光子数还有可能是多个生物体产生的发光。为了使不同流速下生物发光量的研究有可比性,下面设计了单种生物体浓度相同时不同流速流场刺激下的受激发光测量,只考虑相同刺激条件下单位时间内的光子数,不涉及生物体数目对发光的影响。

图3 3 种藻在低速层流刺激下的发光随时间的变化。(a)多边膝沟藻;(b)夜光虫;(c)月牙形梨甲藻。Fig.3 Representative time series of bioluminescence of alga in laminar flows.(a)Lingulodinium polyedrum.(b)Pyrocystis noctiluca.(c)Pyrocystis lunula.
4.3 生物发光与流场剪应力间的相关性研究
多边膝沟藻的浓度约为600 个/mL,夜光虫和月牙形梨甲藻的浓度相同约为50 个/mL,测量不同流速下单位时间内的生物发光情况(每一个流速下的测量值均是在该流速下长时间测量的平均值),结果如图4 所示。图中的各个实心圆点代表完全发展的层流刺激下的生物体发光,空心点为湍流刺激下的发光情况,中间的空白区域为未完全发展的过渡区域。由前面管流的水动力学分析可知过渡区域是层流湍流都存在的情况,不利于分析其对发光的影响,所以没有进行实验测量。

图4 发光藻受激发光与管流壁剪应力的关系。(a)多边膝沟藻;(b)夜光虫;(c)月牙形梨甲藻。Fig.4 Relationship between stimulated bioluminescence and shear stress of developed pipe flow.(a)Lingulodinium polyedrum.(b)Pyrocystis noctiluca.(c)Pyrocystis lunula.
从图4 中可以看出,3 种生物体的受激发光存在壁剪应力阈值,即只有当壁剪应力大于某一值时(图中虚线),生物体才能受激发光,否则只能低水平地自发发光;而当剪应力大于临界值后,3 种生物体的发光强度对数值在层流范围内都随壁剪应力对数值的线性增加而线性增大。根据该组测量结果,用最小二乘原理对对数数据进行线性回归分析,可得多边膝沟藻、夜光虫和月牙形梨甲藻受激发光的壁剪应力阈值分别为0.29,0.06,0.047 N/m2,其发光量随壁剪应力变化的指数分别为2.8(R2=0.94)、2.6(R2=0.97)和3.4(R2=0.98)。也就是说对多边膝沟藻而言,只有当流场中的剪应力大于0.29 N/m2时,生物体才能够受激发光;当剪应力大于阈值0.29 N/m2后,生物受激发光的光子数I 与流场剪应力τ 的关系为I=aτ2.8(a 为常数)。
4.4 实验结果讨论
Anderson 等人在1988年的研究指出,生物体的受激发光能够在流场应力、加速度以及压强等物理量突变的地方发生。后来,Rhor 以及Latz 等人的研究表明,不是由于流场物理量的突变使生物发光,而是流场内剪应力本身的刺激使生物体发光[10]。我们的研究也正是基于此开展的,通过测量的流场流速、压强差和生物发光量,以及计算出的相应位置剪应力,得出单位时间内的受激发光量在双对数坐标下随剪应力的增加而线性增大。这一结论与目前该领域的国外研究相吻合。Latz 等人在二十世纪九十年代开展了大量的不同海域腰鞭毛虫受激发光的现场实测,并采用线性回归拟合的方法计算了引起受激发光的剪应力阈值(也是本文中采用的方法)。其中,对多边膝沟藻的研究较为透彻,实验测得多边膝沟藻的剪应力阈值为(0.32 ±0.03)N/m2,发光量随剪应力变化的指数为3.4 ±0.4(R2=0.94)[8]。从图4中的实验结果可以看出本文与他们的研究相符,但是由于受时间和实验条件的限制,每种情况下的实验次数非常有限,从而无法对实验结果进行统计学分析。从实验的测量值还可以看出,无论是层流还是湍流都能刺激生物体发光,通常情况下湍流刺激产生的生物发光量要大于层流情况,这是由于湍流会使发光生物体的数量增加,而不是使单个生物体的发光量增大。
5 结 论
通过自主设计的管流刺激生物发光实验系统开展了流场刺激与生物受激发光相关性的实验研究,结果表明:引起生物体发光的剪应力存在剪应力阈值,只有当剪应力大于阈值,生物体才能被激发发光;根据实验中选取的腰鞭毛虫种类的不同,剪应力阈值约为0.05~0.29 N/m2;在层流范围内,双对数坐标系下生物体的受激发光量随着剪应力的增加而线性增大。
[1]Stokes M D,Deane G B,Latz M I,et al.Bioluminescence imaging of wave-induced turbulence[J].J.Geophys.Res.:Oceans,2004,109(C1):C01004-1-8.
[2]Rohr J,Hyman M,Fallon S,et al.Bioluminescence flow visualization in the ocean:an initial strategy based on laboratory experiments[J].Deep-Sea Res.Ⅰ,2002,49:2009-2033.
[3]Rohr J,Latz M I,Fallon S,et al.Experimental approaches towards interpreting dolphin-stimulated bioluminescence[J].J.Exp.Biol.,1998,201:1447-1460.
[4]Latz M I,Nuaen J C,Rohr J.Bioluminescence response of four species of dinoflagellates to fully developed pipe flow[J].J.Plankton Res.,2004,26(12):1529-1546.
[5]William F H,John A B.Outline of Theory and Problems of Fluid Dynamics[M].Beijing:Science Press,2002.
[6]Zhang Z S,Cui G X.Hydrodynamics[M].Beijing:Tsinghua University Press,2006.
[7]Cao J,Wang J A,Wu R H.Stimulated bioluminescence by fluid shear stress associated with pipe flow[J].J.Phys.:Conference Series,2011,277(1):012003-1-5.
[8]Latz M I,Rohr J.Luminescent response of the red tide dinoflagellate lingulodinium polyedrum to laminar and turbulentflow[J].Limnol.Oceanogr.,1999,44(6):1423-1435.
[9]Rohr J,Schoonmaker J,Losee J,et al.Flow visualization in the ocean-implications of laboratory bioluminescence experiments[C]//OCEANS'99 MTS/IEEE,Riding The Crest into The 21st Century,Seattle:IEEE,1999,1:145-156.
[10]Rohr J,Losee J,Anderson G.The response of bioluminescent organisms to fully developed pipe flow[R].Technical Report 1360,August 1994.Naval Command,Control and Ocean Surveillance Center,USA.
猜你喜欢
防爆电机(2022年5期)2022-11-18 07:40:18
汽车实用技术(2022年9期)2022-05-20 06:00:26
四川轻化工大学学报(自然科学版)(2021年1期)2021-06-09 06:12:18
科学(2020年1期)2020-08-24 08:07:58
广东医科大学学报(2020年4期)2020-08-24 07:11:10
当代水产(2019年7期)2019-09-03 01:02:24
安徽工业大学学报(自然科学版)(2014年4期)2014-07-11 01:45:46
遗传(2014年3期)2014-02-28 20:59:04
河南科技(2014年23期)2014-02-27 14:18:52
河南科技(2014年13期)2014-02-27 14:11:25

- 发光学报的其它文章
- Synthesis and Fluorescence Properties of Terbium and Samarium Complexes with 2,4,6-Tri(2-pyridyl)-1,3,5-triazine
- ICP 刻蚀工艺对LED 阵列电流输运特性的影响
- Er3+/Yb3+ 共掺KLaF4纳米晶的制备和上转换发光
- CdSe/ZnSe/ZnS 量子点在单晶太阳能电池中的应用
- Transient Analysis of InGaN/GaN Light-emitting Diode with Varied Quantum Well Number
- N 掺杂纳米ZnO 的制备及光催化性能