
4种蒿属植物的光合光响应曲线及其拟合模型
2013-10-17 12:20:42姬明飞丁东粮吴寿方梁少华邓建明
草业科学 2013年5期
姬明飞,丁东粮,吴寿方,梁少华,邓建明
(1.草地农业生态系统国家重点实验室 兰州大学生命科学学院,甘肃 兰州730000;2.南阳师范学院生命科学与技术学院,河南 南阳473061;3.甘肃省陇南市食品药品监督管理局,甘肃 陇南746000)
植物的光合作用是整个生态系统的原初动力,长久以来关于光合效率指标的变化、联系以及控制和调节机理,一直是生态学关注的焦点[1-3]。植物的光合作用是一个复杂的生物物理化学过程,受到环境生态因子和生理因素的共同影响。其中植物净光合速率与光合有效辐射强度之间关系的研究被认为是研究植物光合生理过程对环境响应的基础[4-5]。自Blackman在1905年提出第一个光合作用光响应模型以来,国内外学者建立了多种光响应模型[6-10]。然而,各种方法所拟合出的光合响应参数在与实测值的比较上表现出一定的差异,所以关于不同模型的优缺点存在很大的争议[11-14]。因直角双曲线模型和非直角双曲线模型可以精确地预测在常规双曲线和Blackman光饱和点[15]之间的光响应曲线,最受关注[16]。虽然与光合相关的文献中经常提及光合模型拟合出的指标和参数[17-18],但是对于这些指标和参数缺乏深刻的理解,出现不同模型指标混用的现象,导致在光合拟合模型选择时缺乏可信度[19]。
蒿属(Artemisia)在全世界约有380种,我国分布有180余种和44个变种。蒿属植物分布很广,在温带和热带山区、草原、荒漠、高山和亚高山草原等生态环境中起着重要作用[20]。蒿属植物用途十分广泛,大多具有较高的粗蛋白、粗脂肪含量,且适口性较好,是理想的牧草,也是我国重要的水土保持和防风固沙植物[21]。此外,该属许多种类可以入药,具有消炎、止血、利尿、解毒,抗癌等多种功效[22]。我国蒿属植物资源丰富,进一步研究和发掘其生理变化特征,对于开发蒿属植物的用途具有重要的指导意义。本研究旨在通过两种典型的光响应拟合模型在4种蒿属植物中的应用及其光合效率参数的变化来探讨光响应曲线模型的适用性,同时也为蒿属植物今后作为主要的药用植物的引种、栽培提供科学的参考。
1 材料与方法
选取毛莲蒿(A.vestita)、蒙古蒿(A.mongolica)、牡蒿(A.japonica)以及灰苞蒿(A.roxburghiana)4种蒿属代表性植物为研究材料,将其种植于泥炭土和珍珠岩等体积比混合的花盆内,并随机摆放于兰州大学草地农业生态系统国家重点实验室榆中试验站的全日光温室大棚中(35°56.61′N,104°09.08′E,海拔1 750 m)。在整个试验过程中,平均日照时间为10 h,一天内温度变化基本维持在20~25℃,CO2浓度基本与大气浓度持平,保证足够的空间以防止植物之间的遮阴。
1.1 光响应曲线的测定方法 所有待测植株经1 h光活化后,每种选取3~5株,每株选取3片当年生的成熟叶片用于光响应曲线的测定。根据先前的预试验并结合西北地区的太阳辐射,光合有效辐射强度分别设定为0、40、80、150、200、400、600、800、1 200和1 600μmol·m-2·s-1。利用便携式光合作用 分 析 系 统 LI-6400(Li-Cor Inc,Lincoln NE,USA)中的内置红蓝光源叶室(Li-6400-02B LED)按照1 600到0μmol·m-2·s-1顺序控制光合有效辐射强度,测定不同光合有效辐射梯度下的净光合速率(Amax)。在测定过程中,其它外界环境基本保持一致,即CO2浓度控制在400μmol·L-1,叶片的温度基本控制在28℃,相对湿度控制在52%。
1.2 拟合模型
1.2.1 直角双曲线模型 在1913年Michaelis-Menten就提出了直角双曲线的模型[7],表达式为:

式中,A为总光合作用速率(μmol·m-2·s-1),I为光合有效辐射强度(PAR,μmol·m-2·s-1),a为表观量子效率,Amax为光饱和点时的最大同化速率(μmol·m-2·s-1),Rd为暗呼吸速率(μmol·m-2·s-1)。
1.2.2 非直角双曲线模型 直角双曲线模型中假定光合作用是单一的酶催化反应,这显然与实际情况不相符。Herrick等[23]和 Thornley[7]根据酶促反应动力学原理,提出非直角双曲线模型:
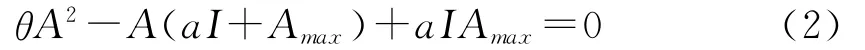
式中,A为总光合作用速率(μmol·m-2·s-1);θ为非直角双曲线的凸度;且当θ=0时,公式(2)即转换为公式(1),即为直角双曲线模型;当θ≠0时,公式(2)转换为:
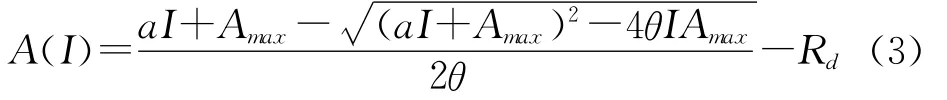
式中,θ为反映光响应曲线弯曲程度的曲角参数,取值0≤θ≤1。
由于直角和非直角双曲线在有限光强下不会达到饱和,本研究采用最大光合速率时的光合有效辐射强度作为饱和点(Lsp)[18,24]。
1.3 数据分析 本研究数据应用Excel软件进行初步处理,用SPSS 13.0软件拟合光响应曲线以及对数据进行差异性分析。
2 结果与分析
2.1 两种拟合方法的比较 本研究分别用两种模型对4种蒿属植物的光合-光响应曲线进行拟合,结果表明,两种模型拟合度相关系数(R2)均在0.925~0.998。通过两种拟合方式得到的4种蒿属植物光响应曲线参数,仅有表观量子效率(a)在两种模型间均差异显著(P<0.05),而光饱和点和补偿点等拟合指标差异不显著(P>0.05)(表1)。表观量子效率(a)反映植物光合作用对光的利用效率,是利用4种蒿属植物的净光合速率在PAR小于200 μmol·m-2·s-1的条件下进行拟合得到的结果(图1-2)。除灰苞蒿外,其他3种蒿属植物的直角双曲线拟合得到的表观量子效率(a)显著高于非直角双曲线拟合值(P<0.05),且大于0.07(表1)。前人研究认为,在理论条件下,植物的最大表观量子效率应该为0.080~0.125[25-26],但在试验条件下光反射、折射以及种间差异等原因,造成其表观量子效率(a)低于其理论值(0.04~0.07)[27]。因此,本研究中非直角双曲线拟合的a较符合理论值,而直角双曲线拟合的表观量子效率(a)则偏大。
从4种蒿属植物的两种模型拟合结果的比较中可以看出,直角双曲线模型拟合得到的4种蒿属植物的暗呼吸速率(Rd)高于非直角双曲线模型拟合值,而非直角双曲线模型拟合得到的Rd更接近于实测值;直角双曲线模型拟合得到的最大净光合速度(Amax)在毛莲蒿和蒙古蒿中高于非直角双曲线模型拟合值,而在牡蒿和灰苞高中则低于非直角双曲线模型的拟合值;同时,在毛莲蒿和牡蒿中非直角双曲线模型拟合得到的Amax更接近实测值,而在蒙古蒿和灰苞蒿中则是以直角双曲线模型拟合得到的Amax更接近实测值(表2)。

表1 4种蒿属植物光合光响应曲线拟合参数Table 1 Photosynthetic parameters of four Artemisia species from two fitting models

图1 毛莲蒿和蒙古蒿的表观量子效率Fig.1 Quantum use efficiency of Artemisia vestita and A.mongolica

表2 4种蒿属植物光合光响应曲线测量数据及其拟合值Table 2 The measured and simulated values of photosynthesis light-response curve of four Artemisia species
2.2不同蒿属植物的光响应参数比较 两种模型拟合得到的牡蒿的表观量子效率均为最高,其次是蒙古蒿和毛莲蒿,灰苞蒿的表观量子效率(α)最小(表1),说明牡蒿对光响应最为敏感,其光能利用效率最高。光饱合时的最大净光合速率(Amax)也是对叶片光合能力的一个度量,在一定程度上反映了植物的遗传特性和对生境的适应能力[18]。两种模型拟合的毛莲蒿和牡蒿的Amax高于拟合的蒙古蒿和灰苞蒿,表明毛莲蒿和牡蒿的光合能力最强。而蒙古蒿和灰苞蒿的光补偿点(Lcp)和光饱合点(Lsp)相比毛莲蒿和牡蒿较低,说明蒙古蒿和灰苞蒿对弱光的适应性较强,但对强光的忍受能力较低,光合能力较弱。
3 讨论
用两种模型对4种蒿属植物叶片光合-光响应曲线进行拟合,结果表明,两种模型都能较好地反映净光合速率(Amax)与光合有效辐射强度(PAR)之间的关系,其相关系数(R2)均在0.925~0.998。但相关系数(R2)只是对模型的一个整体评价,主要反映模型与数据间的整体接近程度[23]。因此,从模型拟合的各种参数角度来评价光合模型更具有实验意义。直角双曲线拟合的α、Rd和Lsp数值均高于非直角双曲线拟合结果,而Amax在毛莲蒿和蒙古蒿的直角双曲线拟合值略高于非直角双曲线拟合值,Lcp却呈现出相反的结果。同一组数据利用不同的拟合模型导致拟合参数存在一定的差异,这样的情况增加了比较研究的难度[18]。以往的研究认为,产生这样的结果,是因为非直角双曲线模型引入了曲线弯曲程度,可将光响应曲线中光合作用随光强增加而增加的线性响应阶段与光合作用的光饱合阶段区分开[28]。因此,在光合有效辐射较弱时,直角双曲线的计算公式没有考虑曲线的弯曲程度,使得初始斜率稍微大于非直角双曲线[29];而当光合有效辐射达到光饱合点后,非直角双曲线显示值趋于稳定,而直角双曲线模型拟合值仍存在上升趋势,不符合植物光合作用对光辐射强度的生理变化特点。所以直角双曲线拟合出的参数大小和曲线的变化趋势不符合实际情况,只有在初始斜率较小时,直角双曲线模型拟合得到的Amax才更符合实测值[26]。总体而言,两种光合响应曲线的拟合方式基本上能反映出4种蒿属植物对于光强变化的响应趋势,而α和Lsp在两种拟合方式间表现出显著的差异性,更是直接表现出两种拟合方式在对植物光利用策略上预测准确度的差异。综上所述,本研究认为非直角双曲线模型在4种蒿属植物的光合光响应曲线拟合中表现较好。
光响应曲线反映了在一定光照辐射强度下植物潜在的光合能力以及植物对生长环境的适应性[30]。本研究所测的4种蒿属植物的光合能力存在一定的差异。毛莲蒿和牡蒿表现出较大的净光合能力和对强光的忍受能力,同时牡蒿具有很高的表观量子效率,表现出对弱光较强的利用能力,而毛莲蒿对弱光的利用率低于牡蒿。对蒙古蒿和灰苞蒿而言,它们对强光的忍受能力较低,因而具有较小的最大净光合能力。4种蒿属植物的光合作用表现出对不同光资源利用能力差异,是长期应对各自不同的生境所造成的[31]。牡蒿为多年生草本,广泛分布于我国湿润和半湿润地区[21],对光资源的利用范围较广;而灰苞蒿为半灌木状草本,多分布于我国西部干旱地区,受到水分亏缺以及白天日照辐射量强的影响,因而灰苞蒿具有较高的光能利用效率(即在较低光照辐射强度下光合速率已达到饱合状态)和低光合能力来减少水分散失。因此,在引种过程中,应考虑当地的实际情况,选取不同的植物。特别是引种具有较高药用价值的牡蒿时,需选择较为湿润的地区种植。
[1] Awada T,Radoglou K,Fotelli M N,etal.Ecophysiology of seedlings of three Mediterranean pine species in contrasting light regimes[J].Tree Physiology,2003,23:33-41.
[2] 张小全,徐德应.18年生杉木不同部位和叶龄针叶光响应研究[J].生态学报,2001,21(3):409-414.
[3] 陈根云,俞冠路,陈悦,等.光合作用对光和二氧化碳响应的观测方法探讨[J].植物生理与分子生物学学报,2006,32(6):691-696.
[4] 张继澍.植物生理学[M].北京:高等教育出版社,2006:104-148.
[5] Elfadl M A,Luukkanen O.Field studies on the ecological strategies ofProsopisjuliflorain a dry land ecosystem:A leaf gas exchange approach[J].Journal of Arid Environments,2006,66:1-15.
[6] Baly E C C.The kinetics of photosynthesis[J].Proceedings of the Royal Society of London.Series-B,Biological Sciences,1935,117:218-239.
[7] Thornley J H M.Mathematical Models in Plants Physiology[M].New York:Academic Press,1976.
[8] Farquhar G D,Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in laves of C3species[J].Planta,1980,149:78-90.
[9] Bassman J,Zwier J C.Gas exchange characteristics ofPopulustrichocarpa,PopulusdeltroidsandPopulus trichocarpa×P.deltroidsclone[J].Tree Physiology,1991,8:145-149.
[10] Prado C H B A,Moraes J De.Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field condition[J].Photosynthetica,1997,33:103-112.
[11] Evans J R,Jakonbsen I,Ögren E.Photosynthetic light-response curves.2:Gradients of light absorption and photosynthetic capacity[J].Planta,1993,189:191-200.
[12] Kyei-Boahen S,Lada R,Astatkie T,etal.Photosynthetic response of carrots to varying irradiances[J].Photosynthetica,2003,41:1-5.
[13] Yu Q,Zhang Y Q,Liu Y F,etal.Simulation of the stomatal conductance of winter wheat in response to light,temperature and CO2changes[J].Annuals of Botany,2004,93:435-441.
[14] Leakey A D B,Uribelarrea M,Ainsworth E A,etal.Photosynthesis,productivity,and yield of maize are not affected by open-air elevation of CO2concentration in the absence of drought[J].Plant Physiology,2006,140:779-790.
[15] Blackman F F.Optima and limiting factors[J].Annals of Botany,1905,19:281-296.
[16] 张中峰,黄玉清,莫凌,等.岩溶植物光合-光响应曲线的两种拟合模型比较[J].武汉植物学研究,2009,27(3):340-344.
[17] 张进忠,林桂珠,林植芳,等.几种南亚热带木本植物光合作用对生长光强的响应[J].热带亚热带植物学报,2005,13(5):413-418.
[18] 张中峰,黄玉清,莫凌,等.岩溶区4种石山植物光合作用的光响应[J].西北林学院学报,2009,24(1):44-48.
[19] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,121(8):76-79.
[20] 周利娟,桑晓清,孙永艳,等.蒿属植物的农药活性及其有效成分[J].江西农业大学学报,2012,34(4):699-705.
[21] 林有润.中国植物志(第七十六卷第二分册)[M].北京:科学出版社,1991:253-255.
[22] 蒋林,林有润.中国篙属植物比较形态和解剖学研究(I)叶表皮结构[J].植物研究,1993,13(4):353-369.
[23] Herrick J D,Thomas R B.Effects of CO2enrichment on the photosynthetic light response of sun and shade leaves of canopy sweetgum trees(Liquidambarstyraciflua)in a forest ecosystem[J].Tree Physiology,1999,19:779-786.
[24] 吴家兵,关德新,孙晓敏,等.长白山阔叶红松林主要树种及群落冠层光合特征[J].中国科学D辑,2006,36(A01):83-90.
[25] 钱莲文,张新时,杨智杰,等.几种光合作用光响应典型模型的比较研究[J].植物科学学报,2009,27(2):197-200.
[26] 于贵瑞,王秋凤.植物光合、蒸腾与水分利用的生理生态学[M].北京:科学出版社,2010.
[27] 许大全.光合作用效率[J].上海:上海科学技术出版社,2002.
[28] 张弥,吴家兵,关德新,等.长白山阔叶红松林主要树种光合作用的光响应曲线[J].应用生态学报,2006,17(9):1575-1578.
[29] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[30] 韦兰英,曾丹娟,张建亮,等.岩溶石漠化区四种牧草植物光合生理适应性特征[J].草业学报,2010,19(3):212-219.
[31] 李熙萌,卢之遥,马帅,等.沙生植物差巴嘎蒿光合特性及 其 模 拟 研 究 [J].草 业 学 报,2011,20(6):293-298.
猜你喜欢
中学生数理化·七年级数学人教版(2023年9期)2023-11-30 03:09:24
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:28
中学生数理化·七年级数学人教版(2019年4期)2019-05-20 10:03:44
价值工程(2017年31期)2018-01-17 00:34:27
电子测试(2017年12期)2017-12-18 06:35:46
中学生数理化·七年级数学人教版(2017年9期)2017-08-15 00:53:19
中学生数理化(高中版.高二数学)(2017年1期)2017-04-16 05:33:43
高中生·天天向上(2016年8期)2016-11-22 09:22:46
水利科技与经济(2016年7期)2016-04-25 13:03:12
水利科技与经济(2016年8期)2016-04-22 03:41:22
