
自愿转轮运动对抑郁模型大鼠学习记忆及基底外侧杏仁核c-fos表达的影响
2013-09-27 08:07崔建梅药宏慧李中华庞立杰贺继平45
体育学刊 2013年5期
崔建梅,药宏慧,李中华,庞立杰,贺继平45
(1.中北大学 体育学院,山西 太原 030051;2.太原理工大学 体育学院,山西 太原 030024;2.山西体育职业学院,山西 太原 030006;4.山西医科大学 汾阳学院,山西 汾阳 030001)
抑郁症(depression)是情感性精神障碍之一,它是一种以情感病态变化主要是情绪低落为显著特征的精神性疾病。抑郁症的发病率呈逐年上升趋势,专家预测2020年抑郁症将成为人类的第二杀手[1]。应激性生活事件与抑郁症的发病存在密切关系,已成为诱发抑郁的重要因素,应激可以引起机体许多功能的改变,诸如认知功能、消化功能、性功能、内分泌功能等。现代生活带给人类的应激因素很多,包括社会竞争压力越来越大、生活环境不稳定性增加等。学习记忆能力降低是抑郁症状的重要内容,许多实验结果表明一定程度的应激可以引起大鼠的学习记忆能力减退[2],而研究证实,运动锻炼具有改善人们不良情绪的作用,对于抑郁症的发病有一定的预防作用[3]。
对动物和人类的研究表明应激激素皮质醇可以对记忆功能起到重要调节作用[4]。杏仁核属于边缘系统的皮质下结构,杏仁核在不同层次调控海马处理信息的过程,参与情感发生和记忆的保存[5],是严重心理应激时中枢神经系统发生变化的主要区域之一,进一步的研究还显示杏仁核的不同核团在情绪记忆巩固过程中有着不同功能。其中,杏仁核基底外侧核是感觉信息与行为输出的情感意义之间联合学习的关键部位[6],在情绪记忆的巩固中具有关键性作用[7]。c-fos基因是原癌基因家族成员,它编码的核蛋白 fos调节靶基因转录,是细胞内信号传递通路中非特异转录调节因子,人们发现c-fos基因表达能反映神经细胞的功能状态,是抗抑郁药在中枢极为快捷有效的指标[8],与神经元的信号转导与认知功能和记忆有关,而且 c-fos基因的表达对长期记忆的保持是必需的[9]。慢性不可预知性应激(Chronic Unpredictable Stress,CNS)是近年来国内外应用最为广泛的抑郁动物模型之一,它可模拟抑郁的核心症状——快感缺乏,目前被广泛运用于抑郁症的病理生理机制、抗抑郁药物研发及作用机制的研究。自愿转轮运动是动物在正常的活动时间和已经熟悉的环境中,没有外力逼迫而自愿进行运动,观察到的结果更接近动物的实际情况。因此,本实验通过复制CNS大鼠模型,观察长期慢性自愿转轮运动对CNS模型大鼠血清皮质醇的影响,测试大鼠空间学习记忆能力及杏仁基底外侧核 c-fos的变化,探讨自愿转轮运动提高抑郁模型大鼠学习记忆能力的可能机制。
1 材料与方法
1.1 动物及分组
雄性健康成年SD大鼠40只(3月龄),体重200~220 g,室温(20±2) ℃,湿度(55±10)%,12 h昼夜循环光照。大鼠购回后适应环境1周,将40只大鼠随机分为4组,即正常对照组、运动组、应激模型组及应激运动组,每组10只。应激模型组及应激运动组单独喂养。
1.2 慢性不可预知性应激模型[3]制备
制备慢性中等不可预知应激抑郁动物模型。应激刺激为24 h禁水、5 min悬尾、2 h束缚应激、5 min冷水强迫游泳(水温 4 ℃)、24 h 禁食、5 min 热应激(45 ℃)、昼夜颠倒、20 min电击足底(0.8 A,每次持续4 s、间隔5 min),以上刺激顺序随机安排,每天1次,连续28 d。非应激组大鼠除了必要的例行鼠笼清洁,不做任何干扰。
1.3 试剂
皮质醇放射免疫试剂(天津德普生物技术医学产品有限公司);c-fos测定试剂盒(武汉博士德生物工程有限公司)。
1.4 训练方案
1)运动组从第 2周接受自愿转轮锻练,笼内有一可自由转动的跑轮(直径33 cm,宽度10 cm),动物可自由进跑轮进行锻炼,红外线检测装置和数据分析系统自动采集和分析转轮情况,运动组及应激运动组大鼠允许适应自愿转轮1周。2)应激模型组从第2周开始接受为期4周的CNS 程序。3)应激运动组从第2周接受自由转轮锻炼,同时每天接受1次CNS程序,为期 4周。4)正常对照组除了必要的例行鼠笼清洁,不做任何干扰。
1.5 血清皮质醇测试
CNS及运动训练结束后第2天上午(09:00—11:00),所有大鼠经尾静脉取血,低温离心分离血清,置于4 ℃冰箱保存,用放射免疫分析法测大鼠血清皮质醇(Cort)。
1.6 旷场试验
尾静脉取血后第2天进行旷场实验以评估大鼠的一般运动能力及探索行为。测试旷场实验箱大小为l00 cm×l00 cm×50 cm,周壁为黑色,底面分为面积相等的25个方格(20 cm×20 cm)。旷场反应箱正上方装有一个摄像头监视大鼠在箱内的活动,并将图像信息传入计算机,再通过回放录像,观察大鼠5 min内穿格数目(crossing squares,以3只脚爪进入1个新格为准)、直立次数(the times of rearing,以2只前肢离开底面为准)、修饰次数及粪便颗粒,每只测试结束后进行粪便的清理,防止遗留气味影响下一只测试。
1.7 八臂迷宫实验程序
所有大鼠在第6周周一进行八臂迷宫实验。实验前大鼠先在迷宫中适应2 d,每天2次。适应时3至4只大鼠同时置于迷宫中,自由活动和摄取食物10 min,适应后进行每天2次的训练。每次训练中,八臂中只有4臂放置了食物(分别为1、3、5、7号臂),整个实验过程均维持此顺序。大鼠置于迷宫中央区,此时中央区四周用鼠门关住,15 s后将门打开并开启分析测试软件,测试定时10 min,大鼠可选择进入任意一臂,以摄取食物。大鼠进入有食物的臂且摄取了食物为 1次正确选择;分析记忆的记录参数为错误选择的次数,重新进入放食物臂或第 1次进入放食物臂而不摄取食物为工作记忆错误(working memory error,WME),进入不放食物臂称为参考记忆错误(reference memory error,RME),两者之和为总记忆错误(total error,TE)当大鼠将4臂食物吃完或10 min后,实验结束,记录所需时间(TT)。
1.8 脑组织的取材及免疫组织化学染色
八臂迷宫测试后即刻,大鼠经常规灌注固定,脑冠状位连续冰冻切片,片厚40 μm,隔4张取1张。切片经0.5% Triton X-100 PBS液孵育(37 ℃)1 h,然后加入10%山羊血清工作液孵育(37 ℃)1 h,加入c-fos血清第一抗体(兔抗鼠),37 ℃孵育5 h后,4 ℃再孵育36 h;随后加入生物素羊抗兔第二抗体,37 ℃孵育1 h;接着放入卵白素生物素复合物中,37 ℃孵育40 min,以上各步骤均用0.01 mol/L PBS (pH=7.2~7.4) 5 min×3次充分漂洗3遍(每次5 min);DAB呈色,缓冲液中止显色,用蒸馏水反复漂洗。0.2%明胶裱片、梯度酒精脱水、二甲苯透明、中性树胶封片。
1.9 图像分析和统计学处理
用数码生物显微镜和形态学显微图像分析系统,参照大鼠脑立体定位图谱测定大鼠BLA c-fos免疫阳性产物。每组大鼠各随机选取10张BLA c-fos的片子在10×40倍下进行测量,计数阳性细胞个数、阳性产物面积和灰度。采用SPSSl3.0统计软件包进行统计分析,描述性资料采用平均数±标准差表示,采用单因素方差分析对组间差异进行统计,P<0.05为显著性差异,P<0.01为极显著性差异。
2 实验结果及分析
2.1 大鼠旷场实验测试结果
表1结果显示:与对照组比较,应激模型组及应激运动组大鼠穿越格数、直立次数均显著减少(P<0.01、P<0.05),应激模型组穿越格数平均减少18.79格,直立次数减少 19.49次;运动组及应激运动组大鼠穿越格数、直立次数均显著高于应激模型组(P<0.01);与对照组比较,应激模型组大鼠修饰次数显著减少(P<0.01),平均减少4.6次,粪便颗粒显著增多(P<0.01);经过4周自愿转轮运动,应激运动组及运动组大鼠修饰次数均显著高于应激模型组(P<0.01、P<0.05),粪便颗粒均低于应激模型组(P<0.05、P<0.01),提示慢性应激可以导致大鼠抑郁样行为,而长期自愿转轮运动可以改善大鼠慢性应激导致的抑郁样行为。
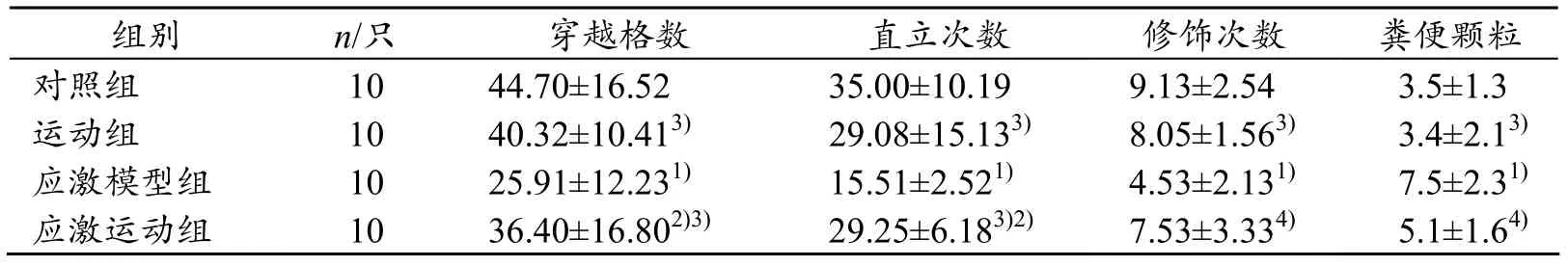
表1 各组大鼠旷场实验测试结果(x±s)
2.2 大鼠血清皮质醇测试结果
结果显示:对照组、运动组、应激模型组、应激运动组血清皮质醇分别为(11.25±2.56)、(13.12±3.45)、(21.46±3.57)、(15.87±2.63)ng/mL。与对照组比较,应激模型组及应激运动组大鼠血清皮质醇水平均明显升高(P<0.01、P<0.05),应激模型组升高10.21 ng/mL;运动组、应激运动组与应激模型组比较皮质醇水平显著下降(P<0.01、P<0.05),应激运动组下降5.59 ng/mL,提示长期自愿转轮运动可能通过拮抗 HPA轴功能亢进从而改善抑郁症状。
2.3 大鼠八臂迷宫行为学测试结果
表2结果显示:与对照组比较,应激模型组及应激运动组完成八臂迷宫时间显著延长(P<0.01、P<0.05),应激模型组增加115.20 s,应激运动组增加42.83 s;与应激模型组比较,经过长期自愿转轮运动,运动组及应激运动组大鼠完成八臂迷宫时间显著缩短(P<0.01),应激运动组平均缩短75.37 s。
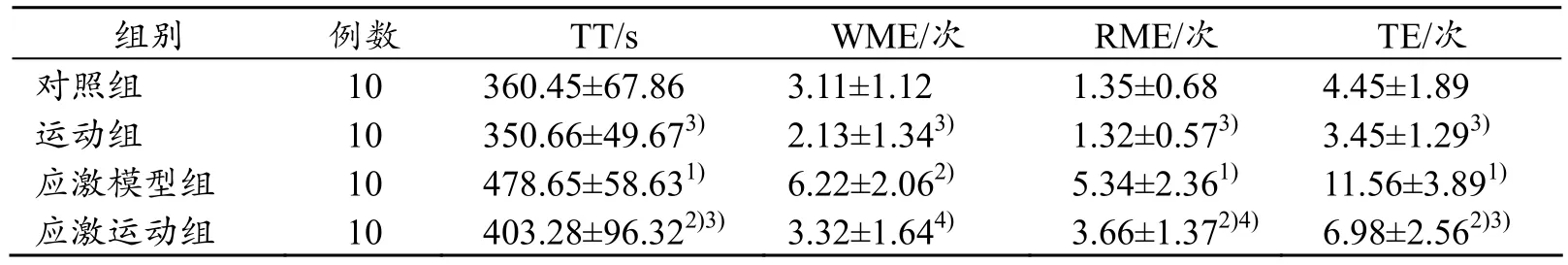
表2 各组大鼠八臂迷宫测试结果(x±s)
与对照组比较,应激模型组重新进入放食物臂或第1次进入放食物臂而不摄取食物的次数(WME)显著增多(P<0.05),平均增多 3.11次;进入不放食物臂的次数(RME)显著增多(P<0.01),总错误次数(TE)远远高于对照组(P<0.01),平均增加7.11次;与对照组比较,应激运动组WME增多,但无统计学差异(P>0.05),RME及TT均显著增多(P<0.05)。
与应激模型组比较,运动组及应激运动组WME、RME均显著减少(P<0.01、P<0.05),总错误次数(TE)远远低于应激模型组(P<0.01),应激运动组平均减少4.58次。
2.4 各组大鼠基底外侧杏仁核(BLA) c-fos免疫阳性细胞数量、面积及灰度
在c-fos光镜下可见BLA内分布有大量的c-fos,免疫反应产物呈棕褐色,低倍镜下为褐色小圆点,染色均匀,高倍镜下为圆形和椭圆形的褐色神经元,主要位于神经元胞体和突起的胞浆内,细胞核着色。
测量结果(见表3)显示:与对照组比较,应激模型组、应激运动组BLA c-fos免疫阳性细胞数量及面积均显著减少(P<0.01),应激模型组降低幅度分别为66%、58%,灰度差异没有显著性(P>0.05);与应激模型组比较,运动组及应激运动组BLA c-fos免疫阳性细胞数量和阳性产物面积表达均增加(P<0.01、P<0.05),应激运动组增加幅度分别为77%、72%,阳性细胞平均目标灰度值虽降低,但差异没有显著性(P>0.05)。

表3 各组大鼠基底外侧杏仁核(BLA)c-fos免疫阳性细胞数量、面积和灰度(x±s)
3 讨论
3.1 自愿转轮运动对抑郁模型大鼠旷场实验的影响
抑郁模型目前可供选择的很多,有应激模型、药物模型、脑损伤模型、遗传选择性模型、基因敲除模型等。其中由Katz等率先提出,后经Willner[10]改进的慢性轻度不可预见性应激模型相对较为可靠,其理论依据与人类抑郁症中慢性、低水平的应激源促进疾病发生、加速疾病发展的机理更接近。研究提示,它可模拟人类抑郁症的核心症状,包括活动减少、好奇探究行为能力下降等[11]。旷场实验是一个用动物行为指标检测类似于人的复杂情绪如焦虑、抑郁等的经典实验,可检测动物的自发行为和探究行为,可用来测试动物中枢神经系统的“兴奋”或“抑郁”状态[12]。穿格数及直立次数表示大鼠的探究行为,修饰(理毛)时间反映大鼠对环境警觉性的高低,排便量反映了紧张恐惧状态。本实验结果显示,应激模型组大鼠穿越格数、直立次数及修饰次数均显著减少、粪便颗粒显著增多,这些行为改变表明长期慢性应激使大鼠活动及探究能力降低、对新鲜环境的好奇程度下降、兴趣丧失、紧张程度增加、对自身的关注下降及运动迟滞,表现为抑郁样。经过4周自愿转轮运动,应激运动组大鼠穿越格数、直立次数及修饰次数均显著增多,粪便颗粒显著减少,说明长期自愿转轮运动可以在一定程度上改善应激大鼠的抑郁样行为,与Salmon等[13]研究结果一致。
3.2 自愿转轮运动对抑郁模型大鼠血清皮质醇及学习记忆的影响
近来的研究显示,抑郁患者血浆皮质醇浓度分泌增加,失去了正常人夜间自发性分泌抑制的节律,整天处于肾上腺皮质功能亢进状态[14]。Rubin等[15]研究还发现,与健康对照相比,抑郁患者的肾上腺皮质增生约38%,增生的程度与皮质醇的浓度有关,且随着抑郁的恢复,这种增生似乎也随着皮质醇的正常化逐渐消失。在本研究中,与对照组相比,CNS显著增加了应激模型组大鼠血清皮质醇水平,平均值增加 11.21 ng/mL,增加幅度为 90%,而经过 4周自愿转轮运动运动应激组大鼠血清皮质醇水平显著下降,尽管远高于对照组,但与应激模型组相比显著降低,平均值减少5.59 ng/mL,说明自愿转轮运动很可能是通过拮抗HPA轴功能亢进,从而改善抑郁症状,在一定程度上发挥抗抑郁作用。运动组经过4周自愿转轮运动血清皮质醇水平有增高的趋势,但与对照组比较无显著差异。
认知功能包括感知觉、记忆、注意、语言、思维、意识、情感、结构运用及高级执行能力、定向力和自知力等。认知功能与情绪之间有着密切联系,最近研究结果表明一定程度的应激可以引起大鼠的学习记忆能力减退,同时血中皮质醇水平显著升高,推测皮质醇分泌增加可能是损伤大鼠空间学习记忆能力以及记忆保持的原因之一。Bosch等[16]发现,皮质醇可以显著损害长时记忆的提取,和服用安慰剂的被试者相比,皮质醇组被试24 h后的单词自由回忆量显著碱少,但直接回忆和再认记忆不受影响。
研究发现,学习记忆能力降低是抑郁症状的重要表现[17-18]。八臂迷宫已被公认为一种比较理想的学习记忆模型,动物通过观察周围一些固定参照物,经过反复识认,从而在其脑内将参照物同空间方位联系起来,来学习和记忆自身与饵(置于迷宫臂末端的白色小食丸)的相对位置,获得辨别该空间方位的能力,是一种评估啮齿动物空间学习记忆能力的工具,已被广泛接受和应用[19]。本实验中,大鼠经过4周慢性不可预知的应激,应激模型组大鼠出现记忆力减退的表现。研究发现,运动锻炼有益于大鼠的学习记忆,具有改善消除人们不良情绪的作用,对于抑郁症有一定的改善作用[20],本研究结果显示经过4周自愿转轮运动运动组大鼠八臂迷宫完成时间,WME、RME及TE均减少,尽管与对照组无显著差异,但仍反映出自愿转轮运动有增强学习记忆的趋势,可能与4周自愿转轮运动对皮质醇的释放有良性的调节作用,能抵抗糖皮质激素的过量分泌。研究表明,认知功能与糖皮质激素的关系呈倒U型,损伤效应与糖皮质激素的不足和过多有关,正常水平的糖皮质激素参与学习记忆的过程[21]。应激运动组大鼠完成八臂迷宫时间显著缩短,平均减少75.37 s,重新进入放食物臂或第一次进入放食物臂而不摄取食物的次数、进入不放食物臂的次数及总错误次数显著减少,总错误次数减少4.58次,说明长期自愿转轮运动可能通过拮抗 HPA轴功能亢进及改善抑郁大鼠脑组织形态和结构的变化,进而改善抑郁大鼠的学习记忆能力,具体机制有待于进一步研究。
3.3 自愿转轮运动对抑郁模型大鼠基底外侧杏仁核c-fos表达的影响
通过大量动物实验和临床实践观察,杏仁核的功能已逐渐被了解,其与学习记忆、情绪调控和神经系统疾病如抑郁症等情绪情感等密切相关[22]。杏仁核内的多种神经递质参与这种调节过程,研究发现,糖皮质激素在参与记忆巩固和提取的过程中具有双向作用且互相关联。BLA 是这一记忆调节系统的关键部位,该部位与海马及其它脑区协同促成糖皮质激素对记忆巩固和提取阶段产生不同的影响,同样在糖皮质激素对记忆提取的损害中起着关键的调节作用[23]。研究发现,选择性的损毁BLA可以抑制糖皮质激素对记忆提取的损害[24]。c-fos基因通常被看作神经元激活的标志物,c-fos是原癌基因家族中一类可以被第二信使诱导的原癌基因,它们能对神经递质、激素等刺激作出反应进行表达,在体内相当于第三信使的作用,它与学习、记忆及运动调控密切相关。研究表明,即刻早期基因c-fos的表达和蛋白质合成是LTP能够维持数周乃至数月的物质基础[25]。He J等[26]研究发现,即刻早期基因c-fos、c-jun及其基因产物的异二聚体调节其他基因的转录,是短暂刺激引起突触连接长期变化的中介,说明原癌基因c-fos、c-jun对于编码空间记忆是必要的。国内宋倩等采用足底电击大鼠,通过Morris水迷宫测试大鼠空间学习记忆能力,然后应用免疫细胞化学方法检测 c-fos在海马和前脑皮层的表达,结果表明,急性应激所致小鼠空间学习记忆功能的改变可能与c-fos蛋白表达的上调有关[27]。
本研究结果显示:长期慢性应激模型组大鼠BLA c-fos 表达显著减少,与Ostrander等[28]研究结果一致,可能是由于长期应激导致基底外侧杏仁核神经元功能不同程度的损害,引起 c-fos在神经元受损后表达降低,同时说明 c-fos可以作为神经元活性的一个敏感指标。而运动组及应激运动组大鼠自愿转轮运动4周后BLA c-fos阳性细胞及面积明显增多,提示应激大鼠及正常大鼠经过长期自愿转轮运动提高学习记忆能力可能与BLA c-fos表达增强有关,但是BLA c-fos增强的机制仍不清楚。目前的研究已检测到,参与c-fos基因激活的第二信使通路至少有3种,它们是cAMP、Ca2+和甘油二酯依赖的蛋白激酶C (Protein kinase C,PKC)。长期自愿转轮运动使兴奋性氨基酸大量释放,导致 Ca2+内流[29],胞内钙离子浓度升高时,与钙调蛋白(CaM)结合形成Ca/CaM复合物,对转录因子进行磷酸化修饰,从而启动c-fos即刻早期基因的表达[30];运动可以引起脑内突触前膜多巴胺、5-羟色胺等神经递质浓度改变,这些递质作为细胞间传递信息的第一信使,作用于突触后膜的相关受体,BLA内含大量多巴胺能受体、5-羟胺能受体都是与核苷酸环化酶耦联的受体,对细胞内环腺嘌呤核苷酸(cAMP)有上调作用,进而激活c-fos基因的表达[31];上调的cAMP进一步促进谷氨酸合成及递质释放,谷氨酸再作用于突触后天门冬氨酸(NMDA)及非 NMDA受体,使突触后神经元兴奋产生长时程增强效应,继而激活即刻早期基因c-fos[32],Ca2+/钙调蛋白依赖性蛋白激酶信号通路是脑内是脑内参与学习记忆的重要通路之一,CaMK可被NMDA受体的活化所激活,参与突触可塑性的调节[33]。研究认为由NMDA受体启动和维持的LTP是学习记忆过程中主要的神经机制之一[34]。shimizu等[35]认为NMDA受体的激活是LTP诱导环节上最为关键的一步。
[1] Imai H,Wada T,Sakamoto R,et al. Activities of daily living rather than depressive symptoms increase the risk of mortality in Japanese community-dwelling elderly people:a 4-year longitudinal survey[J]. J Am Geriatr Soc,2012,60(6):1191-1193.
[2] Wolf O T,Bauser D S,Daum I. Eyeblink conditional discrimination learning in healthy young men is impaired after stress exposure[J]. Psychophysiology,2012,49(2):164-171.
[3] Zheng Hang,Liu Yanyou,Li Wei. Beneficial effects of exercise and its molecular mechanisms on depression in rats[J]. Behavioural Brain Research,2006,168:47-55.
[4] Tse Y C,Bagot R C,Hutter J A,et al. Modulation of synaptic plasticity by stress hormone associates with plastic alteration of synaptic NMDA receptor in the adult hippocampus[J]. PLoS One,2011,6(11):e27215.
[5] Gavin C F,Rubio M D,Young E,et al. Myosin II motor activity in the lateral amygdala is required for fear memory consolidation[J]. Learn Mem,2011,19(1):9-14.
[6] Cooke S F,Bliss T V. Plasticity in the human central nervous system[J]. Brain,2006,129:1659-1673.
[7] Wells A M,Lasseter H C,Xie X,et al. Interaction between the basolateral amygdala and dorsal hippocampus is critical for cocaine memory reconsolidation and subsequent drug context-induced cocaine-seeking behavior in rats[J]. Learn Mem,2011,18(11):693-702.
[8] 韩济生. 神经科学纲要[M]. 北京:北京医科大学-中国协和医科大学联合出版社,1993:23-24.
[9] Countryman R A,Kaban N L,Colombo P J.Hippocampal c-fos is necessary for long-term memory of a socially transmitted food preference[J]. Neurobiology of Learning and Memory,2005,84(3):175-183.
[10] Willner P. The validity of animal models of depression[J]. Psychopharmacology,1984,83(1):1-16.
[11] Konkle A T,Baker S L,Kentner A C,et al.Evaluation of the effects of chronic mild stressors on hedonic and physiological responses:sex and strain compared[J]. Brain Res,2003,992:227-238.
[12] Chkhartishvili E,Maglakelidze N,Babilodze M,et al.Changes of open field behavior in animal model of depression[J]. Georgian Med News,2011,11(200):107-112.
[13] Liu W,Xu Y,Lu J,et al. Swimming exercise ameliorates depression-like behaviors induced by prenatal exposure to glucocorticoids in rats[J]. Neurosci Lett,2012,524(2):119-123.
[14] Takebayashi M,Kagaya A,Uchitomi Y,et a1.Plasma dehydroepiandrosterone sulfate in unipolar major depression short communication[J]. J Neural Transm,1998,105(4-5):537.
[15] Rubin R T,PhilliPs J J,Mccraeken J J,et a1.Adrenal gland volume in major depression relationship to basal and stimulated Pituitary-adrenal cortical axis function[J]. Biological Psychiatry,1996,40(2):89.
[16] Xavier Bosch. Stress hormone impairs long-term retrieval of memorised information[J]. Neuroscience,2000,3:313-314.
[17] Zhang Z H,Wu L N,Song J G,et al. Correlations between cognitive impairment and brain derived neurotrophic factor expression in the hippocampus of post-stroke depression rats[J]. Mol Med Report,2012,6(4):889-893.
[18] Shif O,Gillette K,Damkaoutis C M,et a1. Effects of Ginkgo biloba administered after spatial learning on water maze and radial arm maze performance in young adult rats[J]. Pharmacol Biochem Behav,2006,84(1):17-25.
[19] Valladolid-Acebes I,Stucchi P,Cano V,et al. High-fat diets impair spatial learning in the radial-arm maze in mice[J]. Neurobiol Learn Mem,2011,95(1):80-85.
[20] Marais L,Stein D J,Daniels W M. Exercise increases BDNF levels in the striatum and decreases depressive-like behavior in chronically stressed rats[J].Metab Brain Dis,2009,24(4):587-597.
[21] Abdul Aziz N H,Kendall D A,Pardon M C. Prenatal exposure to chronic mild stress increases corticosterone levels in the amniotic fluid and induces cognitive deficits in female offspring,improved by treatment with the antidepressant drug amitriptyline[J]. Behav Brain Res,2012,231(1):29-39.
[22] Ramot A,Akirav I. Cannabinoid receptors activation and glucocorticoid receptors deactivation in the amygdala prevent the stress-induced enhancement of a negative learning experience[J]. Neurobiol Learn Mem,2012,97(4):393-401.
[23] Roghayeh Pakdel, Ali Rashidy-Pour.Glucocorticoid-induced impairment of long-term memory retrieval in rats:An interaction with dopamine D2 receptors[J]. Neurobiology of Learning and Memory,2006,85(3):300-306.
[24] Dominic M. Dwyer. Lesions of the basolateral,but not central,amygdala impair flavour-taste learning based on fructose or quinine reinforcers[J]. Behavioural Brain Research,2011,220:349-353.
[25] Tanaka K,Weda N,Ogewa N. Chronic cerebral hypopeffusion induces transient eversible rnonoaminegic changes in the rat brain[J]. Neurochern Res,2000,25(3):313-319.
[26] He J,Yamada K,Nabeshima T. A role of Fos expression in the CA3 region of the hippocam- pus in spatial memory formation in rats[J]. Neuropsychopharmacology,2002,26(2):259-268.
[27] 宋倩,孔宏,张亚楠,等. 急性应激对小鼠空间学习记忆功能及海马和前脑皮层c-fos蛋白表达的影响[J].曲阜师范大学学报:自然科学版,2009,35(4):89-92.
[28] Ostrander M M,Ulrich-Lai Y M,Choi D C,et al.Chronic stress produces enduring decreases in novel stress -evoked c-fos mRNA expression in discrete brain regions of the rat[J]. Stress,2009,12(6):469-477.
[29] Schober T,Knollmann B C. Exercise after myocardial infarction improves contractility and decreases myofilament Ca2+sensitivity[J]. Circ Res,2007,100(7):937-939.
[30] Zhao R,Liu L,Rittenhouse A R. Ca2+influx through both L-and N-type Ca2+channels increases c-fos expression by electrical stimulation of sympathetic neurons[J].Eur J Neurosci,2007,25(4):1127-1135.
[31] Zhang W,Tingare A,Ng D C,et al. IP(3)-dependent intracellular Ca(2+) release is required for cAMP-induced c-fos expression in hippocampal neurons[J]. Biochem Biophys Res Commun,2012,425(2):450 -455.
[32] Allen C E,Worsley M A,King A E,et al. Fos expression induced by activation of NMDA and neurokinin-1 receptors in the trigeminal subnucleus caudalis in vitro:role of protein[J]. Brain Kinases Res,2011,1368:19-27.
[33] Moriguchi S,Shioda N,Han F,et al. Galantamine enhancement of long-term potentiation is mediated by calcium/calmodulin-dependent protein kinase II and protein kinase C activation[J]. Hippocampus,2009,19(9):844-854.
[34] Manahan-Vaughan D,Wildfbreter V,Thomsen C.Rescue of hippocampal LTP and learning deficits in a rat model of psychosis by inhibition of glycine transporter-1(GlyT1)[J]. Eur J Neurosci,2008,28(7):1342-1350.
[35] shimizu E,Tang Y P,Rampon C,et a1. NMDA receptor-dependent synaptic reinforcement as a crucial process for memory conso1idation[J]. Science,2000,290(5494):1170-1174.
猜你喜欢
浙江大学学报(医学版)(2023年2期)2023-08-08
中国急救医学(2022年2期)2022-11-15
磁共振成像(2022年6期)2022-07-30
——“AABC”和“无X无X”式词语
小学生学习指导(中年级)(2021年3期)2021-04-06
中国医学影像技术(2021年2期)2021-03-30
少儿美术(快乐历史地理)(2020年8期)2021-01-04
疯狂英语·新读写(2020年3期)2020-06-06
音乐生活(2020年12期)2020-01-05
中国体育教练员(2017年2期)2017-07-31
兰台世界(2017年12期)2017-06-22
