
拟穴青蟹(Scylla paramamosain)NOS蛋白结构分析及同源建模
2013-09-24 07:57王世佳林浩鹏李升康
汕头大学学报(自然科学版) 2013年2期
王世佳,林浩鹏,李升康
(1.汕头大学广东省海洋生物技术重点实验室,广东 汕头 515063;2.汕头大学生物系,广东 汕头 515063)
0 引言
拟穴青蟹(S.paramamosain),属于十足目(Decapoda),梭子蟹科(Portunidae),青蟹属(Syclla de Hann),广泛分布于我国东南沿海,包括浙江、福建、台湾、广东、广西和海南沿海水域[1].拟穴青蟹属于暖水,广盐性蟹类,肉质鲜美,营养丰富,个体大,生长快,适应性强,具有较高的经济价值,在我国传统水产养殖中占有重要地位,也是汕头牛田洋围垦区养殖的主要品种之一.近年来,汕头沿海青蟹养殖基地屡遭病害,导致了青蟹产量及经济效益的严重损失[2].研究青蟹免疫相关基因,探索青蟹的先天性免疫机制,从而更好的为青蟹病害的免疫防治服务就显得十分必要.在拟穴青蟹中,诱导型NOS基因(inducible NOS,iNOS)能被一些细胞因子或免疫刺激物诱导,被认为是一种典型的免疫相关基因.通过一氧化氮合成酶合成的NO,主要参与信号转导及免疫调控[3],在青蟹的先天性免疫防御中起非常重要的作用.本研究旨在通过分析拟穴青蟹NOS蛋白的理化性质、二级结构及三级结构,并在三级结构的基础上进行空间结构的同源建模,为进一步研究SpNOS空间结构和免疫功能的关系奠定基础.
1 材料与方法
1.1 NOSs基因与生物信息学分析软件
拟穴青蟹NOS蛋白的基因序列来自NCBI数据库(http://www.ncbi.nlm.nih.gov/),其登录号为FR875156.1.其他物种NOS蛋白登录号为:南美白对虾(Litopenaeus vannamei)(ADD63793.1),囊对虾(Marsupenaeus japonicus)(BAI67609.1),斑节对虾(Penaeus monodon)(ACJ54486.1),眼斑龙虾(Panulirus argus)(ACZ60615.1),滨蟹(Carcinus maenas)(ACY56317.1),黑背陆栖蟹(Gecarcinus lateralis)(AAT46681.1);三级结构可视化软件Cn3D,RasWin及AntheProt 3D Viewer;序列分析软件DNAStar;分析结果显示软件Antheprot Graphic Viewer.建模软件EASR-Modeller,评估网站为http://nihserver.mbi.ucla.edu.
1.2 SpNOS一级结构分析
利用Expert Protein Analysis System(ExPASy)的蛋白分析工具对SpNOS蛋白的氨基酸组成、相对分子质量、等电点等理化性质进行分析.用ScanProsite分析蛋白的功能域.采用ProtScale工具(http:∥www.expasy.ch/tools/protscale.html)中的Kyte and Doolittle算法对SpNOS蛋白进行疏水性分析,通过Expasy的2ZIP数据库预测亮氨酸拉链结构[4].利用HMMTOP在线服务器http://www.enzim.hu/hmmtop/html/submit.html分析SpNOS蛋白的跨膜结构[5].
1.3 SpNOS二级结构分析
利用CFSSP在线服务器(http://www.biogem.org/tool/chou-fasman/)[6],Sopma(http://npsapbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)[7]在线服务器,Psipred在线服务器(http://bioinf.cs.ucl.ac.uk/psipred/)等对SpNOS蛋白的二级结构进行分析,包括α-螺旋、β-转角、无规则卷曲及延伸链等[8].
1.4 SpNOS三级结构分析
利用Swiss-Prot在线分子模建服务器及Phyre2服务器对SpNOS蛋白序列进行三级结构分析.通过BLAST分析得到模板链,利用(Procheck)将对照模板构建的三维结构进行评估.利用EASY-Modeller建模并进行能量的优化,优化后利用Procheck对模建结果进行检测,计算得出Ramachandran图[9].

图1 SpNOS蛋白各种氨基酸组成比例图
2 结果
2.1 SpNOS氨基酸组成特性分析
分析结果(表1)表明,拟穴青蟹NOS蛋白由1214个氨基酸组成,原子总数为
19 137个,分子式为C6100H9469N1711O1807S50,分子量为137 290.6 Da,理论等电点为6.56.SpNOS蛋白中各种氨基酸的比例见图1.在所有组成氨基酸中,亮氨酸含量
最高为9.1%,色氨酸含量最低为1.6%. 拟穴青蟹NOS蛋白氨基酸组成与其他物种比较结果表明,拟穴青蟹SpNOS蛋白与眼斑龙虾(Panulirus argus)NOS蛋白氨基酸组成最为相似.
2.2 SpNOS结构域分析
将SpNOS蛋白的氨基酸序列提交到ScanProsite服务器,对SpNOS蛋白进行翻译后修饰位点结构分析.结果显示,该蛋白中有1个黄素氧还原蛋白类似物域,一个铁氧还蛋白型黄素腺嘌呤二核苷酸结合域,19个酪氨酸激酶II磷酸化位点,5个酪氨酸激酶磷酸化位点,11个蛋白激酶C磷酸化位点,16个豆蔻酰化位点,2个酰胺化位点,2个N-糖基化位点,1个cAMP和cGMP依赖型蛋白激酶磷酸化位点,并在其序列中发现一段双向核定位域(422-437)和一段30AA长的脯氨酸富集域(1171-1201).
利用HMMTOP在线服务器http://www.enzim.hu/hmmtop/html/submit.html分析SpNOS的跨膜结构.结果表明,其为胞外蛋白,没有跨膜结构.
采用ProtScale工具(http://www.expasy.ch/tools/protscale.html)中的Kyte和Doolittle算法对SpNOS蛋白进行疏水性分析.结果表明,该蛋白中含有较多亲水性氨基酸,亲水性较强.
将SpNOS蛋白序列提交至Expasy的2ZIP数据库预测亮氨酸拉链结构.结果显示,SpNOS不含亮氨酸拉链结构.
2.3 SpNOS二级结构分析
通过CFSSP服务器(http://www.biogem.org/tool/chou-fasman/)预测SpNOS的二级结构成分.结果表明,SpNOS中可能形成α-螺旋的氨基酸残基有790个,占氨基酸总数的65.1%;可能形成延伸链的残基有764个,占氨基酸总数的62.9%;可能形成不规则卷曲的氨基酸残基有152个,占氨基酸总数的12.5%.
通过SOPMA服务器(http://npsa-pbil.ibcp.fr/cgi-bin/secpred_sopma.pl)预测SpNOS的二级结构成分.结果表明,α-螺旋和不规则卷曲交替存在,是SpNOS整体结构的主要结构元件.蛋白的两段都由α-螺旋占据,一端有一段不规则卷曲聚集区,仅有少量α-螺旋存在.延伸链均匀分布在整条链中.这种分布有利于蛋白质结构的稳定.
2.4 SpNOS蛋白的三级结构分析及同源建模
由于SpNOS的氨基酸数目达到1000以上,整条链建模并不十分可靠.因此,通过对三段进行分割分别建模,三段分别为1-422个氨基酸(模型一),423-919个氨基酸(模型二),920-1214个氨基酸(模型三).三段蛋白采用的模板分别为2G60(小鼠抗FLAG抗原结合片段),3HR4(人类一氧化氮合成酶及钙调蛋白复合体),1QGY(念珠藻铁氧还蛋白),得到三个结构,到PROCHECK中检查,得到了可靠的结果(图2ABC).其拉氏图的可靠性分别达到98.3%,98.2%和96.8%(图2BCD).
3 讨论
一氧化氮合酶(nitric oxide synthase,NOS)是NO合成过程中的关键酶.目前,已经确定的NOS有神经元型(neuronal NOS,nNOS)、内皮型(endothelial NOS,eNOS)和诱导型(inducible NOS,iNOS)三种,它们作用于精氨酸生成NO而发挥生物学效应.NO除具有神经传导和松弛平滑肌等功能外,还具有抗菌、抗病毒、抗寄生虫等作用[10],它可以通过作用于病原体的核酸、蛋白质和脂类等杀灭病原体[11].
已经证实,SpNOS是一种诱导性的NOS(iNOS)[3].诱导型一氧化氮合成酶iNOS能被一些细胞因子或免疫刺激物诱导.近几年iNOS作为抗病力指标在哺乳动物、鱼类、以及昆虫和贝类等无脊椎动物中已有比较深入的研究[4].研究发现,白斑综合症病毒(WSSV)在感染中国明对虾初期可以诱导血细胞产生iNOS,随着WSSV在中国明对虾体内的大量增殖及其对血细胞的破坏,使得iNOS活性显著降低,对虾也趋于死亡.由此,iNOS能够作为反映对虾在病毒感染过程中健康状况的有效指标,在水产免疫方面具有重大意义[12].
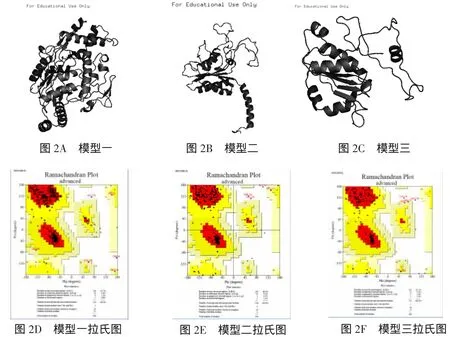
图2 三维结构模型及拉氏图分析
同源建模已经广泛应用于各种生物体的基因功能预测[13-16].本研究以SpNOS为对象,首先在理化性质方面与眼斑龙虾,南美白对虾,滨蟹,囊对虾,斑节对虾,黑背陆栖蟹等六种甲壳类动物的NOS蛋白做了比较,发现在氨基酸残基总数,原子总数,碱性氨基酸总数,酸性氨基酸总数,理论等电点方面,拟穴青蟹NOS蛋白与眼斑龙虾NOS蛋白更为接近.
跨膜结构的预测结果显示NOS蛋白没有跨膜结构.疏水性分析的结果显示拟穴青蟹NOS蛋白中含有较多的亲水性蛋白,亲水性较强,也与NOS是非跨膜蛋白的结果相符.亮氨酸拉链预测结果表明拟穴青蟹NOS蛋白序列中没有亮氨酸拉链结构,可以推测NOS蛋白不是DNA结合蛋白.在功能域分析中,发现SpNOS的N-端有一段长达30个氨基酸(1171-1201)的脯氨酸富集域,推测该区域可能与调节蛋白结合,引发级联反应,作用于免疫通路.
二级结构对蛋白质的稳定性有着很大影响.本研究所采用的chou-fasman方法是比较经典的预测方法.SOPMA服务器和Pispred服务器是目前应用较广也较为成熟的二级结构预测服务器,两者都是通过对氨基酸评分来预测可能的二级结构的.SOPMA服务器相比chou-fasman方法,筛选更加严格,避免了一个氨基酸形成多种二级结构的可能.Pispred服务器的评分系统比SOPMA服务器更为严格,结果展示也更为直观.三种方法的预测结果都显示NOS蛋白的主要结构元件是α-螺旋和不规则卷曲,散在的延伸链散布在整条链中.
本实验通过同源建模的方法,得到SpNOS三部分的模型,其拉氏图结果表明结构比较合理.但是,整个SpNOS分子的三维结构还需进一步研究.由于SpNOS长达1214个氨基酸,PDB数据库中还无法找到对应整条链的模板蛋白.于是,我们通过对其三个不同的保守域进行分别建模,得到三个模型.PROCHECK检测发现,模型质量不太好.因此,本次研究利用了EASY-Modeller进行建模.通过Blast搜索PDB中同源性最高的蛋白,以同源性最高的蛋白质为模板建模,建立其模型后,通过软件附属功能可以对其实现能量最小化(共轭梯度法)和动力学模拟,进行简单的人为修改,而且所得模型的能量能通过表格自动展现出来,本实验得出来的结果通过PROCHECK分析,符合质量要求.
甲壳类动物不存在特异性免疫,研究具体的免疫相关基因可以丰富我们对其先天性免疫系统的认识.拟穴青蟹的NOS蛋白空间结构的模拟研究对于探究拟穴青蟹NOS在其非特异性免疫系统中的作用意义重大.本研究利用生物信息学的方法对拟穴青蟹的NOS蛋白进行了一级、二级结构预测和同源建模分析.但由于蛋白质结构性质和作用途径及其复杂,预测结果需进一步实验验证才能得出确切的结论.
[1]林琪,李少菁,黎中宝,等.中国东南沿海青蟹属(Scylla)的种类组成[J].水产学报,2007,31(2):211-219.
[2]李升康,朱烁,张昭,等.利用cDNA-AFLP技术研究副溶血弧菌感染下拟穴青蟹的基因差异表达[J].水产学报,2012,36(4):503-508.
[3]Li S K,Zhang Z,Li C B,et al.Molecular cloning and expression profiles of nitric oxide synthase(NOS)in mud crab Scylla paramamosain[J].Fish Shellfish Immunol,2012,32(4):503-512.
[4]王鹤,张高华,王旭达,等.獐茅耐盐基因SOS1的克隆及序列分析[J].中国农业大学学报,2012,3:28-33.
[5]李任峰,田香勤,何启盖,等.大肠杆菌P4 FliC蛋白结构分析及同源建模[J].生物技术通报,2010,10:143-148.
[6]Chou P Y,Fasman G D.Conformational parameters for amino acids in helical,β-sheet,and random coil regions calculated from proteins[J].Biochemistry,1974,13(2):211-222.
[7]Geourjon C,Deléage G.SOPMA:significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments[J].Comput Appl Biosci.1995,11(6):681-684.
[8]Gómez-Anduro G A,Sotelo-Mundo R R,Muhlia-Almazán A.et al.Tissue-specific expression and molecular modeling of cytosolic manganese superoxide dismutases from the white shrimp Litopenaeus vannamei[J].Dev Comp Immunol.2007,31:783-789.
[9]Kuntal B K,Aparoy P,Reddanna P.EasyModeller:A graphical interface to MODELLER[J].BMC Res Notes.2010,3:226.
[10]Chakravortty D,Hensel M.Inducible nitric oxide synthase and control of intracelular bacterial pathogens[J].Microbes and Infection,2003,5:621-627.
[11]Kro¨ncke K D,Fehsel K,Kolb-Bachofen V.Nitric oxide:cytotoxicity versus cytoprotection-how,why,when,and where?[J].Nitric Oxide,1997,l(2):107-120.
[12]姜国建,于仁诚,王云峰,等.中国明对虾(Fenneropenaeus chinensis)血细胞中一氧化氮合成酶的鉴定及其在白斑综合症病毒感染过程中的变化[J].海洋与湖沼,2004,5(34):342-350.
[13]孟珍,尤晓颜,姜成英,等.系列硫氧化还原酶(SOR)的同源建模[J].应用与环境生物学报,2010,16(3):424~428.
[14]张海瑞,翟军军,窦永喜,等.小反刍兽疫病毒非结构蛋白V基因的克隆及其编码蛋白的结构与功能[J].中国兽医科学,2011,41(03):235-239.
[15]柳广飞,周集体,王竞,等.Phodobacter shoaeroides偶氮还原酶三级结构同源建模与分析[J].大连理工大学学报,2010,50(2):171-175.
[16]陈锋菊,刘元东,高健,等.嗜酸氧化亚铁流感菌的核糖-5-磷酸异构酶的同源建模研究[J].中山大学学报,2010,49(4):93-98.
猜你喜欢
华人时刊(2023年1期)2023-03-14
汉字汉语研究(2021年2期)2021-08-30
当代水产(2019年7期)2019-09-03
汉字汉语研究(2019年2期)2019-08-27
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
学生天地(2017年24期)2017-11-07
河北书画研究(2016年3期)2016-04-28
动物医学进展(2015年10期)2015-12-07
食品工业科技(2014年7期)2014-03-11
