
云南与印度蝗虫区系的比较
2013-09-19 09:28毛本勇杨国辉
大理大学学报 2013年10期
毛本勇,李 超,李 淼,杨国辉
(大理学院农学与生物科学学院,云南大理 671003)
云南地处中国西南边陲,北纬21°9′~29°15′,东经97°39′~106°12′,东部与贵州、广西为邻,北部同四川相连,西北隅紧倚西藏,西部同缅甸接壤,南部和缅甸、老挝、越南毗连,总面积3.94×105 km2。云南气候深受印度洋西南季风和西风环流季节交替的影响,具有浓郁的低纬度高原气候特点。云南干、湿季分明,年降水量大部分地区800~1600 mm;年温差较小,年均气温5~24℃;由于地形极为复杂,西北高,东南低,境内高山林立,峡谷深嵌,立体气候明显,南北温差达19℃左右。全省除西北端属湿润高原气候、滇南低热河谷区属北热带湿润季风气候外,大部分属亚热带高原型季风气候等7个气候类型〔1〕。
印度位于亚洲南部,北纬 8°04′~37°06′,东经68°07′~97°25′,与巴基斯坦、中国、尼泊尔、不丹、缅甸和孟加拉国为邻,濒临孟加拉湾和阿拉伯海,总面积29.8×105 km2。印度北部为高山区,中部为平原,南部大部分属热带草原,西南部为热带雨林,平原、高原、缓丘占国土面积的60%左右;印度北部属高原气候,年平均气温12~14℃,中东部属热带季风气候,年均气温26~29℃,干、湿季分明,年降水量大部分地区500~1500 mm。
两地理区位置毗邻,纬度重叠,同属东洋界,但其地质历史悬殊,云南地形、地貌和局域气候更为复杂多样。生物多样性的大尺度分布格局是现代环境与地史过程共同作用的结果〔2〕。基于对云南和印度两地现代环境与地史过程的认识,比较两地区蝗虫区系的异同,能够为揭示其区系渊源、推测物类的演化过程提供参考,反之也可为推测地史演化过程提供借鉴。
1 材料与方法
1.1 材料 数据和地理分布信息来源于文献〔3-6〕。
1.2 方法 采用D.Otte(1995)直翅目分类系统〔7〕,以蝗虫属级单元和种级单元为研究对象,比较其多样性和丰富度,揭示两地区蝗虫物类的区系特点;根据共布单元的分布状况,揭示两地区蝗虫物类的相关性。遵循分布越广的单元起源越早、多样性大的高级单元起源较早、含特有单元越多的高级单元起源越早、地区特有单元起源较晚、单元数多的地区可能为其起源中心、化石产地可能为其起源中心的原则〔8-9〕,推测物类起源中心;根据物类的分布模式、扩散能力强弱(长翅种类>短翅种类>鳞翅种类)和生态适应性来推断扩散方向。
2 结果
2.1 多样性 云南现有蝗总科和锥头蝗总科昆虫3科14亚科92属225种(亚种),含云南特有属20属(占21.74%),特有种138种(占61.33%)。印度现有同类昆虫4科20亚科158属337种(亚种),含印度特有属37属(占23.42%),特有种127种(占37.69%)。可见,两地蝗虫既包含大量的本地特有成分,又混合了一些与其他地区共同分布的成分,反映了两地蝗虫区系性质的特殊性和复杂性;云南特有种成分比重较大的结果反映了其更为明显的物种分化事实,表明其区系成分较印度更加特殊;两地均为蝗虫属级阶元的重要起源地。
2.2 丰富度 单位面积蝗虫属级阶元丰富度云南为23.35×10-5属/km2,印度为5.30×10-5属/km2,前者为后者的4.41倍;单位面积蝗虫种级阶元丰富度云南为57.11×10-5种/km2,印度为11.31×10-5种/km2,前者为后者的5.15倍。王志恒等〔10〕认为,随地形复杂度的增加,物种丰富度和物种密度呈现先增大后减小的趋势。从地形、地貌和气候等环境因子上看,云南生境异质性较大。异质性大的生境能够为生物提供特殊的庇护场所,避免突发性事件(如第三纪中后期的全球气候变冷)造成的物种灭绝〔2〕。此结果可能预示着云南蝗虫物类丰富度与其生境异质性程度有正相关关系。
2.3 共布成分相似性 两地区蝗虫共布成分共有49属47种。采用相似性系数(similarity coefficient,Sc)计算公式Sc=〔2C/(A+B)〕×100%(A=甲地区物类数,B=乙地区物类数,C=甲、乙两地区共有物类数)〔11〕计算其相似性。云南和印度蝗虫共布属相似性系数=2×49/(92+158)=39.20%,为中等不相似水平;共布种相似性系数=2×47/(225+337)=16.73%,为极不相似水平。可见,两地蝗虫在属级阶元和种级阶元的相似性都不高,说明它们在起源上的同源性较小。
2.4 共布种分布模式 两地蝗虫共布种47种,共有4种分布型:即东洋种27种,占共布种总数的57.45%;东洋+古北种13种,占27.66%;东洋+非洲种2种,占4.26%;东洋+古北+非洲种5种,占10.64%。其中东洋+古北种均主要分布在东洋界,仅在两界交界处呈过渡分布,加上纯粹的东洋种,比例达85.11%,表明两地蝗虫物种交流主要体现在东洋种内部。这些蝗虫大多为扩散能力较强的长翅种类,扩散方向大约为由南向北,由于生态适应性所限,其分布边界止于古北界东缘。
2.5 共布属分布模式 两地蝗虫共布属有49属,分3类:单区共布属、双区共布属和多区共布属。属的分布范围是所含物种的各分布范围的总和。
2.5.1 单区属 共12属,占共布属的24.49%。单区共布属只分布在东洋界,原则上起源于东洋界,可用于更精细地推断属的起源及物种扩散方向。各属在东洋界各亚界分布的物种数及分布模式见表1。
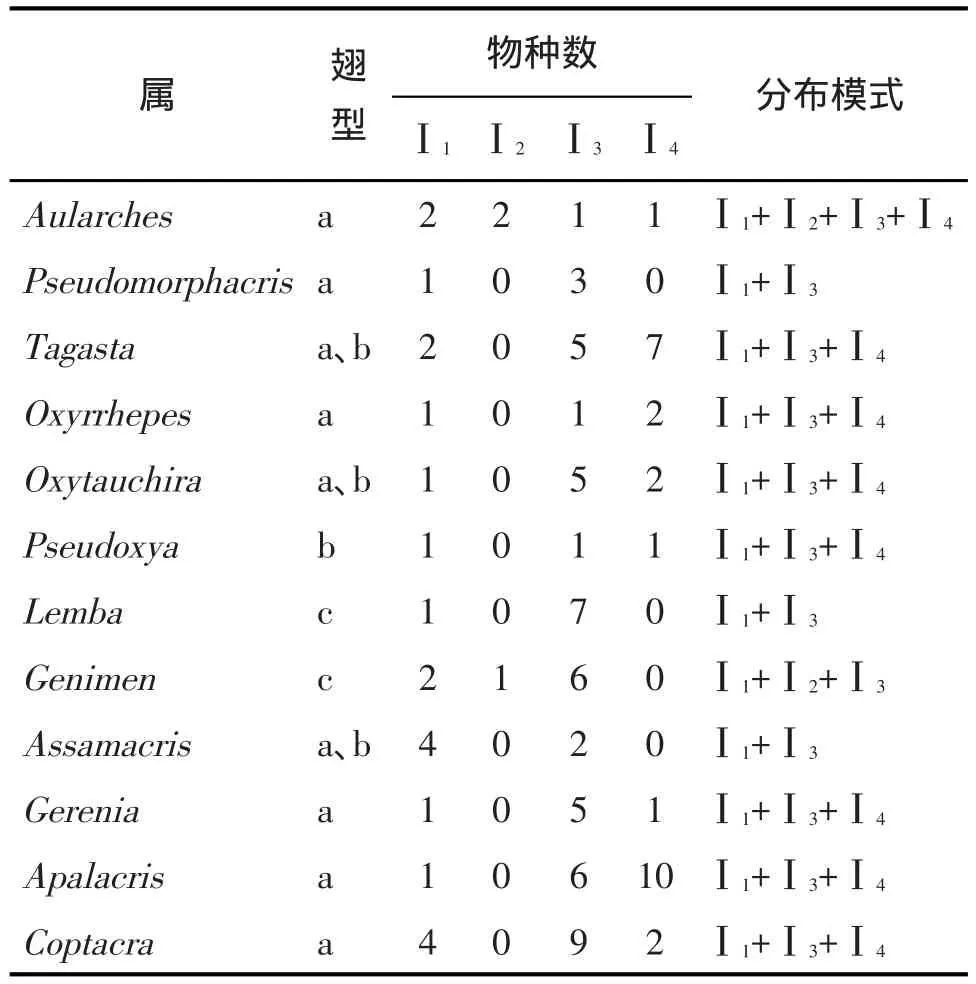
表1 云南与印度蝗虫单区共布属分布物种数及分布模式
从表1可知,单区属有4种分布模式:印度亚界+印中亚界+印马亚界(7属:橄蝗属Tagasta、大头蝗属Oxyrrhepes、板角蝗属Oxytauchira、伪稻蝗属Pseudoxya、点翅蝗属Gerenia、胸斑蝗属Apalacris和切翅蝗属Coptacra)、印度亚界+印中亚界(3属:似橄蝗属Pseudomorphacris、舟形蝗属Lemba和阿萨姆蝗属Assamacris)、印度亚界+锡兰亚界+印中亚界(1属:庚蝗属Genimen)及印度亚界+锡兰亚界+印中亚界+印马亚界(1属:黄星蝗属Aularches)。
2.5.2 双区属 共19属,占共布属的38.78%,在各大区分布的物种数及分布模式见表2。从表2可知,双区属有3种分布模式:东洋+古北属(9属:凸额蝗属Traulia、竹蝗属Ceracris、黄脊蝗属Patanga、踵蝗属Pternoscirta、厚蝗属Pachyacris、素木蝗属Shirakiacris、棉蝗属Chondracris、缺背蝗属 Anaptygus和雏蝗属Chorthippus)、东洋+非洲属(8属:梭蝗属Tristria、卵翅蝗属Caryanda、斜翅蝗属Eucoptacra、十字蝗属Epistaurus、板胸蝗属Spathosternum、蔗蝗属Hieroglyphus、棒腿蝗属Tylotropidius和坳蝗属Aulacobothrus)、东洋+澳洲属(2属:芋蝗属Gesonula和直斑腿蝗属Stenocatantops)。

表2 云南与印度蝗虫共布双区属分布数量及模式
2.5.3 多区属 共18属,占共布属的36.73%,其在各大区分布的物种数及分布模式见表3。从表3可知,多区属有5种分布模式:东洋+古北+非洲属(7属:斑腿蝗属Diabolocatantops、外斑腿蝗属Xenocatantops、黑背蝗属Eyprepocnemis、佛蝗属Phlaeoba、戛蝗属Gonista、疣蝗属Trilophidia和飞蝗属Locusta)、东洋+非洲+澳洲属(2属:刺胸蝗属Cyrtacanthacris和长夹蝗属Choroedocus)、东洋+古北+非洲+澳洲属(7属:负蝗属Atractomorpha、稻蝗属Oxya、车蝗属Gastrimargus、绿纹蝗属Aiolopus、异距蝗属Heteropternis、小车蝗属Oedaleus和剑角蝗属Acrida)、东洋+古北+非洲+澳洲+新热带属(1属:束颈蝗属Sphingonotus)、东洋+古北+非洲+新北+新热带属(1属:沙漠蝗属Schistocerca)。
2.6 共布属的起源与进化 根据分布物种数推断起源中心,根据分布模式和扩散能力并结合生态适应性推断扩散方向,结果大致可以归类为以下4种情况。
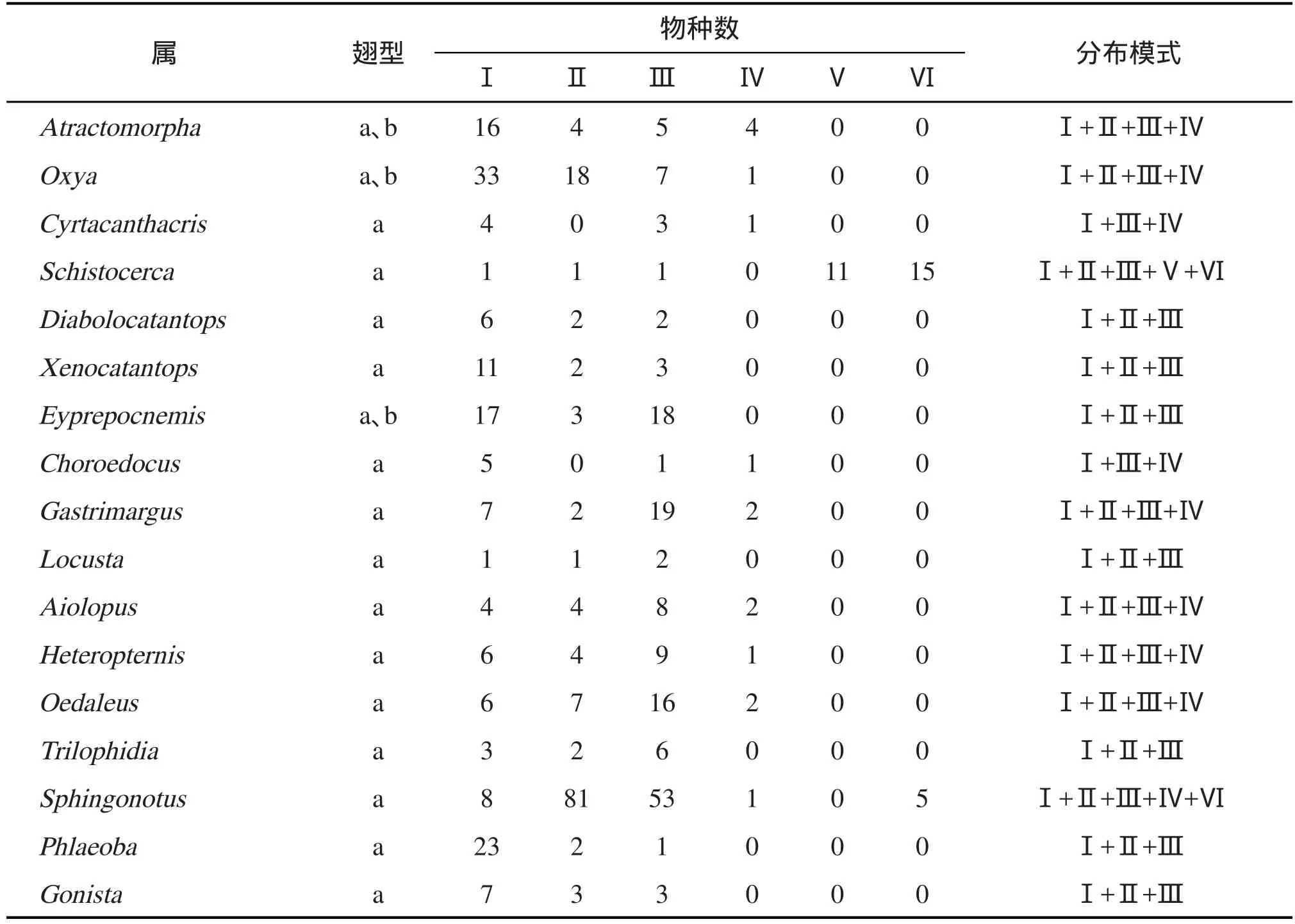
表3 云南与印度蝗虫共布多区属分布数量及模式
2.6.1 东洋界起源向周围扩散 原则上所有单区属均为东洋界起源,其中印中和印马亚界为主要的策源地,例如橄蝗属、板角蝗属、点翅蝗属、胸斑蝗属和切翅蝗属。它们普遍具有较低的生态价,仅适应东洋界生境,在向周围扩散过程中仅向西进入生境较为相似的印度亚界,而未进入温带大陆性气候的古北界。双区属中东洋界起源扩散到古北界的属有凸额蝗属、竹蝗属、黄脊蝗属、踵蝗属和厚蝗属5属;东洋起源扩散到非洲界的属有卵翅蝗属、蔗蝗属、棒腿蝗属、坳蝗属4属;东洋起源扩散到澳洲界的属有芋蝗属和直斑腿蝗属2属。多区属中东洋界起源,扩散到其他地理区的属有负蝗属、稻蝗属、斑腿蝗属、外斑腿蝗属、刺胸蝗属、长夹蝗属、佛蝗属和戛蝗属8属。以上合计31属,占共布属的63.3%,表明东洋区系成分在两地蝗虫交流中占首要位置。
2.6.2 非洲界起源向周围扩散 双区属中非洲界起源扩散到东洋界的属有梭蝗属、板胸蝗属、斜翅蝗属和十字蝗属4属;多区属中非洲界起源向周围扩散的属有黑背蝗属、飞蝗属、车蝗属、绿纹蝗属、异距蝗属、小车蝗属、疣蝗属和剑角蝗属8属。二者合计12属,占共布属的24.5%,表明非洲区系成分在两地蝗虫交流中占次要地位。
2.6.3 古北界起源向周围扩散 双区属中古北界起源扩散到东洋界的属有棉蝗属、缺背蝗属和雏蝗属3属;多区属中古北界起源,扩散到其他地理区的属有束颈蝗属1属。二者合计4属,占共布属的8.2%,表明古北区系成分在两地蝗虫交流中的影响不大。
2.6.4 新热带界起源经新北界扩散 仅沙漠蝗属1属,占共布属的2.0%。
3 讨论
生物现有分布格局的形成是生物类群在古地理、气候演变等多种因素的综合作用下长期发展演化的结果。云南地质构造主要包括3个单元——华南板块、思茅板块(含临沧地块和昌宁—孟连缝合带)和保山板块(含腾冲地块)〔12-14〕。在早泥盆世,华南板块并中南半岛板块和东马来亚板块从组成冈瓦纳古陆的印度—澳大利亚板块边缘断裂、分离,同时西缅甸板块也从冈瓦那古陆分离〔14-15〕。华南板块的西缘大约沿梅里雪山—碧罗雪山—打洛一线延伸入中南半岛〔12〕。这些陆块的动植物区系在古生代后期为温润的赤道特提斯区系成分〔14〕。川滇古陆为华南板块西侧部分,呈狭长纵条,北起北纬32°52′的丹巴之南,向南延伸至北纬22°云南南部,其陆地范围虽经多次变化,但从未被海水完全淹没〔16〕,成为中国西南地区本土昆虫起源的主要策源地〔3〕。晚石炭世至早二叠世,以昌宁—临沧—孟连为代表的思茅块体,通过弧后分离从华南板块分离出来〔14-15〕。早二叠世,包括保山地块、滇缅泰马(Sibumasu)陆块在内的Cimmerian块体从冈瓦那古陆东部分离〔13〕,带着南缘生物的后裔向北漂移,至晚三叠世〔16〕或晚二叠世〔17〕,拼合于华南板块。印度板块和澳大利亚板块自白垩纪从冈瓦纳古陆分离并向北移动,前者于始新世与亚洲大陆相撞,并继续向北漂移导致了喜马拉雅造山运动〔18〕。第四纪青藏高原隆起,使云南地貌格局发生深刻变化。第四纪冰期导致昆明—大理一线以北地区的自然环境由原来的中亚热带常绿阔叶林变成为温带偏湿润针阔叶混交林〔19〕,冰期和间冰期的交替往复引发了海陆变迁及昆虫的南北迁移。这些不同时期的地质历史事件对云南与印度地区蝗虫分布格局的形成产生了巨大影响。
原直翅目(Protorthoptera)昆虫发生在古生代石炭纪的联合古陆时期〔3,20〕,在晚三叠世的地层中仍发现其化石〔21〕,至少在晚二叠世分化出直翅目〔22〕,到晚第三纪有大量属种是现生属种〔23〕。
云南大部地区在原直翅目起源前就与冈瓦那古陆分离,而印度迟至白垩纪才脱离冈瓦那古陆,因此,二者直翅目在起源上的同源性较小。然而,保山陆块在直翅目出现后仍可能与印度板块短暂相连,随即便率先脱离冈瓦那古陆。庚蝗属现今的分布格局很能与上述地质历史相互印证。庚蝗属已知9种,分布于盈江、瑞丽、西双版纳、缅甸西部、斯里兰卡以及印度南部和阿萨姆。庚蝗属无翅,扩散能力极弱,现今的分布格局可能源于二叠纪冈瓦那古陆东部的Cimmerian块体及印度陆块(含斯里兰卡),之后随这些陆块漂移形成现今分布格局。至于共布属中主要为印中亚界和印马亚界起源的区系成分扩散入印度的时间,应该在印度板块与欧亚大陆相撞的始新世以后,扩散的类群主要是翅发达的长翅类群。为何扩散方向多为单向而非双向呢?这可能与印度板块由南向北快速漂移导致环境剧烈改变有关。环境改变后,印度的原生物种因不能适应这种快速变化而灭绝,从而空出了一些生态位。与古北成分比较,来自印中亚界和印马亚界的东洋成分更容易占领这些生态位。非洲起源扩散到东洋界的共布属的扩散途径可归纳为以下两个途径:其一是经印度板块(或许还有Cimmerian块体)扩散到东洋界;其二是经欧亚大陆迂回扩散至印度和云南。共布成分中古北成分少的原因可能与第四纪冰川对两地的影响较小有关。
依靠物类的现生数量推测发生中心也许并不一定符合历史事实,但在寻找到更多化石证据之前,或许这是最客观的推断方法。随着蝗虫区系资料的完善及地史研究的深入,或许会有更多引人入胜的发现。
致谢:大理学院农学与生物科学学院徐吉山博士提供部分文献。
〔1〕吴征镒,朱彦丞.云南植被〔M〕.北京:科学出版社,1987:3-10.
〔2〕唐志尧,王志恒,方精云.生物多样性分布格局的地史成因假说〔J〕.生物多样性,2009,17(6):635-643.
〔3〕毛本勇,任国栋,欧晓红.云南蝗总科区系、分布格局及适应特性〔M〕.北京:中国林业出版社,2011:1-336.
〔4〕Chandra K,Gupta S K,Shishodia M S.A Checklist of Orthoptera(Insecta)of India〔J/OL〕.〔2013-05-03〕.http://zsi.gov.in/checklist/orthoptera.pdf.
〔5〕Nayeem R,Usmani K.Taxonomy and field observations of grasshopper and locust fauna(Orthoptera:Acridoidea)of Jharkhand,India〔J〕.Mun.Ent.Zool.,2012,7(1):391-417.
〔6〕Eades D C,Otte D.Orthoptera Species File Online.Version 2.0/4.1.〔DB/OL〕.〔2013-05-03〕.http://Orthoptera.Species-File.org.
〔7〕张宏杰,霍科科,陈锐.中国夏氏与Daniel Otte蝗虫分类系统比较〔J/OL〕.〔2008-07-10〕.http://jonline.nwu.edu.cn/wenzhang/208053.pdf.
〔8〕袁忠林,沈林,尚素琴,等.缘脊叶蝉亚科生物地理学研究(同翅目,叶蝉科)〔J〕.动物分类学报,2006,31(1):1-10.
〔9〕汉弗莱斯,帕伦特.分支生物地理学:植物和动物分布的解释性格局〔M〕.张明理,译.北京:高等教育出版社,2004:1-166.
〔10〕王志恒,陈安平,方精云.湖南省种子植物物种丰富度与地形的关系〔J〕.地学学报,2004,59(6):889-894.
〔11〕张镱锂.植物区系地理研究中的重要参数:相似性系数〔J〕.地理研究,1998,17(4):429-434.
〔12〕曹仁关.云南省板块构造与古生物地理区的划分〔J〕.中国区域地质,1994(1):52-64.
〔13〕Metcalfe I.Stratigraphy,palaeontology and palaeogeography of the Carboniferous of Southeast Asia〔J〕.Mem.Soc.Geol.France,1984,147:107–118.
〔14〕李朋武,高锐,崔军文,等.西藏和云南三江地区特提斯洋盆演化历史的古地磁分析〔J〕.地球学报,2005,26(5):387-404.
〔15〕Buwett C,Duhig N,Bewy R,et al.Asian and South-western Pacific Continental Terranes Derived from Gondwana,and their Biogeographic Significance〔J〕.Aust.Syst.Bot.,1991,4,13-24.
〔16〕钟章成,秦万成,徐茂其.四川植被地理历史演变的探讨〔J〕.西南师范学院学报,1979(1):1-13.
〔17〕殷鸿福.三叠纪古生物地理与大陆漂移〔J〕.地质科学,1980(3):265-278.
〔18〕Hall R.Cenozoic reconstrutions of SE Asia and the SW Pacific:changing patterns of land and sea〔C〕//Metcalfe I,Smith J,Morwood M,et al.Faunal and Floral Migrations and Evolution in SE Asia-Australasia.Lisse:Swets and Zeitlinger Publishers,2001:35-56.
〔19〕周杰,沈吉.中国西部环境演变过程研究〔M〕.北京:科学出版社,2007:1-221.
〔20〕Carpenter F A.Structure and relationships of the Carboniferous insects Eucanus valis(Protorthoptera:Eucanidae)〔J〕.Psyche,1976,83:223-242.
〔21〕Hong Y C,Li Z Y.The discovery of a new family Sunopteridae fam.nov.(Insecta:Protorthoptera)from Shaanxi Province,China〔J〕.Geological Bulletin of China,2012,35(5):647-652.
〔22〕Carpenter F A.The Lowe Permian Insects of Kansas.Part 11.The Orders Protorthoptera and Orthoptera〔J〕.Psyche,1966,73:46-88.
〔23〕张雅林,袁忠林,高志方.地球发展历史概论及大陆漂移在生物地理分布研究中的应用〔J〕.西北农林科技大学学报:自然科学版,2004,32(6):69-78.
猜你喜欢
农业工程学报(2022年14期)2022-10-19
热带作物学报(2022年9期)2022-10-17
许昌学院学报(2022年3期)2022-07-19
少儿美术(快乐历史地理)(2020年10期)2020-03-18
基层中医药(2018年5期)2018-08-31
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
分析化学(2017年9期)2017-10-16
地球科学与环境学报(2016年4期)2016-08-23
安徽农业科学(2015年20期)2015-10-21
橡塑技术与装备(2015年7期)2015-07-03
