
若尔盖湿地公路建设生态恢复中植物群落多样性与相似性研究
2013-09-14 09:19秦纪洪
水土保持研究 2013年4期
陈 蓓,马 睿,秦纪洪,孙 辉
(1.四川大学 锦城学院 土木与建筑工程系,成都611731;2.泰安市水土保持生态环境监测站,山东泰安271000;3.成都大学 城乡建设学院,成都610106;4.四川大学 环境与科学工程系,成都610065)
国道213线郎川段(郎木寺—川主寺)改扩建工程穿越国家级自然保护区若尔盖湿地,鉴于若尔盖高寒草甸草原和高原湿地地处青藏高原东南缘的生态脆弱区的环境敏感性,以及作为长江与黄河上游重要水源地、高原湿地生物多样性重点区域的生态重要性,在设计与建设时充分考虑了有效的生态与工程措施以降低工程对草原与湿地生境的破坏。高寒草甸草毡层形成时间漫长,一旦破坏不但难以恢复,而且草毡层丧失会直接导致高寒草甸土壤侵蚀与沙化,因此在工程建设时采用了工程前将施工地表的草皮切块堆放贮藏,主体施工完成后将切块堆放的草皮覆盖到工程中破坏的道路两侧地表,从而实现地表植物群落快速恢复和有效保持水土、构建生态公路的目的。这些措施的实施及其效果评价,对于高寒区域工程施工后的生态恢复重建与水土保持具有重要意义。
目前若尔盖生态环境逐渐向沙化草地演变,沙化速度已达平均每年816 h m2[1],且土壤有机质含量低,土壤肥力低,草毡层形成过程很漫长,草产量少[2]。前期的初步试验中表明,通过收集开挖地面的草皮,回复移植加强湿地和草原创面的生态恢复是可行的[3-4],在川西北高原湿地的工程建设中采取草皮移植快速恢复工程破坏生境的措施在一定程度上取得了成效,前期的研究表明,草皮恢复后的植物群落结构稳定且成效与割取草皮厚度、大小、面积相关[5]。对于评估这种生态恢复技术,还需要对恢复群落和原生背景群落开展对比研究[6]。本文对郎川公路工程生态恢复中利用的切割草皮恢复后的植物群落与原生草甸群落的多样性与相似性开展对比研究,以评价在高寒地带工程建设中通过利用原生草皮剥离后再覆盖进行生态恢复技术的可行性与有效性。
1 实验材料与方法
1.1 研究地点概况
实验地点位于郎川公路中北段,属若尔盖县,位于32°11′—33°56′N,102°35′—103°03′E,海拔3 500 m。具青藏高原大陆季风气候特征,冬季寒冷干燥、日照多而强、日温差大,夏季降水明显增多而进入湿季。多年平均气温1.1℃,最热月(7月)多年平均气温10.7℃,最冷月(1月)多年平均气温-10.7℃。年平均降雨量656.8 mm,最大年降水量为862.9 mm,历年单日最大雨强65.3 mm/d,历年平均蒸发量1 233.2 mm,历年平均相对湿度为78%。草地年均日照实际时数为2 417 h,年平均风速2.4 m/s。
1.2 样地调查方法
野外调查地点为郎川公路若尔盖花湖地区草皮移栽试验样地,距公路50 m,样方规格见表1。选取2组不同人工切割规格(面积和厚度)的草皮回覆道路建设破坏的地表群落恢复情况的样方6个,1个背景原生草甸样方,分别进行物种调查,密度、高度、盖度、频度、地上生物量(鲜重和干重)的测定,同时采环刀土样(100 c m3)调查地下部分生物量状况,重复3次。采用SPSS软件Base模块进行统计分析。

表1 调查样方的基本情况
1.3 群落β多样性与相似性测度指标
本研究主要调查群落中的物种数、各物种的个体数、高度、盖度、密度、频度和生物量等能够反映群落结构特征的值,采用通用的5种β多样性指数。β多样性表示物种或物种多度沿着群落内或群落间从一个生境到另一个生境变化的速率和范围[7],采用β多样性指数对群落的测度意义在于可以揭示生境被分割的程度或不同地段的生境多样性[8],且β多样性与α多样性一起构成了群落或生态系统总体多样性或一定地段的生物异质性[9-10],因而可提示群落的结构、功能及动态机制。
1.3.1 Whittaker指数βW:
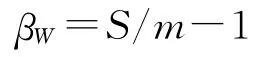
式中:S——系统中记录的物种总数;m——各样方物种的平均数。
1.3.2 Cody指数βc:

式中:g(H)——沿生境梯度 H 增加的物种数目;l(H)——沿生境梯度H 失去的物种数目,即在上一个梯度中存在,而在下一个梯度中没有的物种数目。
1.3.3 Routledge指数(βR,βI,βE)

式中:S——系统中的物种总数;r——分布重叠的物种对数。

式中:ei——种i出现的样方数;αj——样方j的物种数目;T——所有调查样方的物种总数。其中,T=∑ei=∑αj。

1.3.4 群落相似性测度方法 利用Jaccard指数Cj和Sorenson指数Cs测度群落的相似性。

式中:j——两个群落或样方的共有种数;a,b——样方A和样方B的物种数。
1.4 物种重要值指标
重要值计算公式为:IV=(相对高度%+相对盖度%+相对密度%+相对频度%+相对生物量%)/5。
2 结果与分析
2.1 样地物种
经调查,研究区进行道路建设破坏创面生态恢复的回覆草皮样方的物种有32种,隶属8科28属,表2列出了各样方中16种主要物种的重要值,人工移栽草皮植物群落主要为耐寒多年生禾本科和莎草科,优势种和建群种以禾本科(Gr a mineae)的羊茅属(Festuca L.)、早熟禾属(Poa L.)、披碱草(El y mus dahu-ricu),莎 草 科 (Cyper aceae)的 嵩 草 属 (Kobresia willd.),老鹳草(Ger anium orientali-tibeticum)等为主。除禾本科外,菊科(Com-positae)的风毛菊属(Saussurea DC.)、毛茛科(Ranuncul aceae)的银莲花属(Anemone L.)、豆科等若干属的一些种等类群的物种增加,有毒有害物种增加[5]。一些偶见种出现在试验样方中,如鹅绒委陵菜(Potentill a.anserin)等,但背景样方中的偶见种却变为各个草皮试验样方内的常见种。这可能是由于光照的改变使部分喜光植物萌发所致,其次由于切割使草皮具有边缘效应[11],边缘的植物长势好于大片的原生植被。原生样方中还调查到以下偶见种:棘豆(Oxytropis spp.)、米口袋属(Guel denstaedtia spp.)、龙胆(Gentiana spp.)、马先蒿属(Pedicul aris L.)、黄帚橐吾(Ligul aria vir gaurea)、狼毒(Euphorbia f ischeriana)等,在群落中出现的频率很低,多半是由于种群本身数量稀少的缘故,偶见种可能是偶然的入侵种,也可能是群落中的衰退残遗种,因此表2未列出。

表2 各样方主要物种的重要值
2.2 不同样地群落的β多样性
选取不同的测度方法比较原生样方7与其它6个草皮样方,以及每个相邻样方之间的β多样性的变化速率,结果如图1、图2所示。从图1可以看出,五种β多样性具有相似的变化趋势,表明这几种指数具有较好的可加性。原生样方与草皮样方之间的变化起伏说明各样方群落间物种替代速率的变化不同,反映出由于人为切割使得原有生境变得破碎而导致群落的β多样性的变化。随着样方面积从30 c m×30 c m增加到50 c m×50 c m,5种指数均呈上升趋势,且变化趋势十分相似。Routledge指数(βR,βI,βE)变化较为稳定,表明草皮样方受到切割干扰后的生境破碎并不十分明显,作为人工恢复样方具有较好的实践性。

图1 原生样地与各个草皮样地之间β多样性指数的二元数据属性测度结果
βW指数反映β多样性与物种丰富度S之间的关系[12],通过对原生样方与草皮群落βw的计算,发现物种代替程度与取样面积密切相关,βW随着取样面积的增加而增加,这是由于样方内微环境类型或资源异质性增加,而样方间的异质性反而逐渐降低,物种的周转速率也随之降低。郭正刚等[13]的研究表明,在青藏高原北部高海拔地区,当样方面积为8 m2和16 m2时,βW随取样面积的变化差异不再显著,趋于稳定,而本研究中草皮切割的面积均小于该值,这说明如果增加取样面积,草地群落物种发生变异的尺度会更大,物种代替程度就会降低,更有利于群落的恢复。
Cody指数βC[14]可用来测度群落间交错区的物种在环境梯度上的变化或周转速率,表达的是沿环境梯度增加或减少的物种在数量上的概念。通过对新增加和失去的物种数目进行比较,只考虑物种的存在与否,能获得十分直观的物种更替概念。
对比原生样方对应其它各样方(图1)的β多样性和各个切割草皮两两样方间(图2)的β多样性这两类曲线,发现这5种曲线不仅具有一致的变化趋势,而且有很大的相似性,说明β多样性测度方法生态学意义明显,实用价值较高。从实际操作方便、计算简捷等方面考虑,应当首选βW和βC指数进行群落β多样性的测度。βW指数和βC指数均受取样面积的影响,βW指数随取样面积的增加而增加,而βC指数则随取样面积的增加而减少。

图2 各个草皮样地两两之间β多样性指数的二元数据属性测度结果
2.3 物种相似性测定结果
β多样性的另一方面,即不同群落间的β多样性的测度也不容忽视。目前,比较成熟的群落分类和排序技术能较好地解决这一问题,最简便的方法是运用相似性系数测度群落或生境间的β多样性。在众多的指数中,应用最广、效果最好的是Jaccar d指数(Cj)和Sorenson指数(Cs),表3—4列出了不同样方两两之间的相似性对比结果。

表3 各样方间相似性指数Cj计算结果

表4 各样方间相似性指数Cs计算结果
表3—4表明,与原生样方7相似性指数较高的是切割草皮样方2、5、6;切割草皮样方间相似性指数最低的是3—4和1—4。由于样方2,5,6的体积分别为30 c m×30 c m×20 c m、50 c m×50 c m×20 c m和50 c m×50 c m×10 c m,物种丰富度较高,与原生样方相似性较高,样方1,3物种丰富度指数较低,与其他样方的相似性较小。由此可得出,取样面积越大,与背景越相似的群落,恢复效果越好。
3 结论
随着样方面积的增加,五种β多样性指数均呈上升趋势,且变化趋势相似,并较为稳定,说明草皮样方受到切割干扰后的生境破碎不十分明显。增加取样面积,草地群落物种发生变异的尺度会更大,物种代替程度就会降低,更有利于群落的恢复。从实际操作方便、计算简捷等方面考虑,应首选βW和βC指数。对相似性指数的计算结果表明,取样面积越大、与背景越相似的群落,恢复效果越好。因此在进行人工生态恢复时,利用当地自然植物群落的结构,保持群落多层次和多样性结构[15],有利于形成较稳定的植物群落,提高边坡稳定性。
研究区草地生态系统的恢复和重建结合了草皮切割技术和重新铺设技术,利用了高寒草甸地带土壤有机质含量较高、原生植被较好的特点,虽然生物体矮小,但根系密集呈毡状[16],施工时对其分割划块铲起,移植到适宜地方保存,待工程结束后回铺到路基边坡,消减了道路建设造成的生物总量损失。虽然人为切割干扰使草地原有的物种分布被打乱,但整个梯度上的系统结构仍比较有序,各样方群落多样性与相似性分析表明,生态系统功能下降不大,恢复效果良好,对当地湿地景观保护有一定的作用,在种质资源保护区进行草地生态系统保育与提高种质资源方面具有较好的实践意义。
致谢:本研究野外工作得到了国道213郎川公路建设指挥部刘东、四川省草原研究所张昌兵和道里刚的大力帮助,在此谨表谢忱。
[1] 雍国玮,石承苍,邱鹏飞.川西北高原若尔盖草地沙化及湿地萎缩动态遥感监测[J].山地学报,2003,21(6):758-762.
[2] 赵仁昌.若尔盖草地沙化及防治对策[J].四川环境,1995,14(2):15-20.
[3] 陈蓓,孙辉,秦纪洪,等.朗川公路若尔盖湿地段人工移栽草皮的群落生态学研究[J].水土保持研究,2007,14(2):68-71.
[4] 陈蓓,孙辉,秦纪洪,等.高寒地区道路工程的生态效应及其生态恢复研究进展[J].四川环境,2009,28(2):102-105.
[5] 马睿,孙辉,陈蓓,蹇依,等.若尔盖郎川公路建设区有毒有害植物扩散及生态安全性评价[J].世界科技研究与发展,2006,28(3):70-75.
[6] 苗保河,郑延海,伏芳,等.北京市门头沟区石灰窑遗址及公路边坡裸露地表植被修复模式的生态评价[J].水土保持研究,2011,18(6):125-128.
[7] 赵志模,郭依泉.群落生态学原理与方法[M].重庆:科学技术文献出版社,1989:147-148.
[8] 马克平.生物群落多样性的基本原理和方法.[M].北京:中国科学技术出版社,1994:141-165.
[9] Wilson M V,Sch mida A.Measuring beta diversity with presence-absence data[J].J.Ecol.,1984,72:1055-1064.
[10] 马克平,刘灿然,刘玉明.生物群落多样性的测度方法:Ⅱ.β多样性的测度方法[J].生物多样性,1995,3(1):38-43.
[11] 关卓今,裴铁璠.生态边缘效应与生态平衡变化方向[J].生态学杂志,2001,20(2):52-55.
[12] Whittaker R H.Evolution and measurement of species diversity[J].Taxon,1972,21:213-251.
[13] 郭正刚,刘慧霞,王根绪,等.人类工程对青藏高原北部草地群落β多样性的影响[J].生态学报,2004,24(2):384-388.
[14] Cody M L.Towards a theory of continental species diversity bird distributions over Mediterranean habitat gradients[M]∥Cody M L,Diamond M J.Ecology and Evolution of Communities.Cambridge:Harvard University Press,1975.
[15] 罗双,孙海龙,刘冲.四川道路边坡自然恢复的植被多样性研究[J].水土保持研究,2011,18(6):51-57.
[16] 刘兰华,李耀增,康锋锋.高原地区铁路建设生态恢复技术初探:以青藏铁路格唐段为例[J].水土保持研究,2007,14(1):310-312.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
考试与评价·高二版(2021年5期)2021-09-10
落叶果树(2021年6期)2021-02-12
科学技术创新(2021年2期)2021-01-21
林业调查规划(2020年3期)2020-06-03
今日重庆(2018年3期)2018-05-30
文体用品与科技(2018年9期)2018-05-04
体育科技(2016年2期)2016-02-28
