
施氮时期对高产夏玉米光合特性的影响
2013-09-11 08:38张吉旺杨今胜董树亭李登海
生态学报 2013年2期
吕 鹏,张吉旺 ,刘 伟,杨今胜,董树亭,刘 鹏,李登海
(1.作物生物学国家重点实验室/山东农业大学农学院,泰安 271018;2.德州市农业科学研究院,德州 253015;3.山东省玉米育种与栽培技术企业重点实验室,莱州 261448)
氮作为夏玉米必须营养元素,对器官建成、根冠发育、光合作用和源库关系等有重要影响[1]。前人关于氮素对玉米光合特性的影响已有大量报道[2-3]。随施氮量增加,功能叶片的Pn、叶绿素含量、可溶性蛋白含量、Hill反应活性、Ca2+-ATPase活性、Mg2+-ATPase活性和PEPCase活性均不断增大[4]。在一定范围内,玉米叶片的叶绿素含量和光合速率与叶片含氮量呈正相关,氮素供应失调导致光合能力下降[5],同时有研究指出,施氮后光系统Ⅱ(PSⅡ)和光系统Ⅰ(PSⅠ)两个光系统性能的改善及二者协调性的提高增强了光合电子传递性能是灌浆期Pn升高与产量增加的主要原因[6]。
目前,我国玉米生产中存在氮肥施用过量,施氮方式不合理的问题[7]。黄淮海区域典型玉米高产区施氮量为433.5—556.9 kg/hm2,平均用量457.9 kg/hm2[8]。一般生产大田中,农民通常在拔节期一次性施氮,这种方式极易造成“烧苗”,且易造成灌浆期氮素亏缺[9]。生育后期缺氮可导致叶肉细胞叶绿体结构变差,营养器官氮素再分配率增大,引起叶片早衰,同时伤害PSⅡ与PSⅠ之间的电子传递链以及PSⅡ反应中心供体侧和受体侧,从而造成产量下降[10]。因此对于高产夏玉米进行科学的氮肥管理已势在必行[11]。陈国庆等[12]研究了不同施氮量对高产夏玉米冠层的高光谱特征,但目前关于氮素运筹方式对高产夏玉米光合特性的影响鲜见报道。本试验旨在探明施氮时期对高产夏玉米光合特性的影响,为高产夏玉米的氮素管理和夏玉米专用缓控释肥的研发提供科学依据。
1 材料与方法
1.1 试验设计
本试验于2008—2009年在山东农业大学试验农场进行。2008和2009年降水量分别为477.1和459.1 mm,平均气温分别为22.4℃和23.8℃,日照时数分别为770.5 h和813.0 h。土壤为棕壤,其0—20 cm土层基础肥力分别为:有机质 8.81 和 9.21 g/kg,全氮 0.87 和 0.94 g/kg,碱解氮 86.9 和 86.9 mg/kg,速效磷52.65 和 57.43 mg/kg,速效钾129.42 和113.45 mg/kg。试验材料为郑单 958(ZD958)和登海 661(DH661),郑单958是我国目前第一大主推品种,登海661为2005年李登海创造21043 kg/hm2夏玉米高产记录的品种[13]。前茬作物为冬小麦,6月12日播种,10月5日收获,足墒播种,全生育期无灌溉。种植密度75000株/hm2,等行距种植,行距60 cm,小区面积100 m2,3次重复,随机排列。设置6个试验处理,T1为不施氮处理,T2为一次性施氮处理,T4、T5为3次施氮处理,T6为4次施氮处理,施氮量为纯氮360 kg/hm2,肥料为含氮量46%尿素,具体施氮时期及施氮比例见表1。各处理均施P2O5120 kg/hm2(过磷酸钙857 kg/hm2)、K2O 240 kg/hm2(氯化钾400 kg/hm2),分别于播种前和拔节期各施入50%,按高产田进行田间管理。

表1 施氮时期及施氮比例Table 1 Nitrogen application stages and nitrogen application ratio
1.2 测定项目及方法
1.2.1 产量测定
成熟期,每小区收获3行玉米,共30个果穗,调查穗部性状,测定出籽率和含水率,计算产量(按14%折算含水率)。
1.2.2 植株氮素积累测定
成熟期取长势均匀一致的植株5株分为叶片、茎鞘、籽粒,105℃杀青,75℃烘干至恒重,称重后磨粉、测定分析。植株各部位全N含量采用半微量凯氏定氮法测定[14]。
氮素收获指数(NHI,%)=籽粒吸氮量/植株吸氮量×100[15]氮肥偏生产力(NPFP,kg/kg)=施氮区产量/施氮量[15]
1.2.3 叶面积(LAI)及光合色素含量测定
分别于10 叶期(V10),抽雄期(VT),花后14 d,28 d,42 d,收获期(R6)测定叶面积,参照 Arnon[16]的方法测定叶片光合色素含量,测定样品为2009年试验材料。
1.2.4 净光合速率(Pn)及叶绿素荧光参数测定
采用美国LI-COR公司LI-6400光合测定系统测定净光合速率(Pn),开放式气路,冠层果穗叶附近CO2浓度360—380 μmol/mol,应用系统 LED 光源补光,光量子通量为 1400 μmol·m-2·s-1,选择晴天条件下10:00—14:00测定。2008 年测定时间分别为开花后 10、24、38、52,2009 年测定时间分别为 10、26、40、54 d。
采用英国Hansatech公司FMS-2脉冲调制式荧光仪测定叶绿素荧光参数,包括光适应下的最大荧光(Fm')、稳态荧光(Fs),暗适应30 min后的初始荧光(Fo)、最大荧光(Fm)。可得PSⅡ的实际光化学效率ΦPSⅡ、PSⅡ的最大光化学效率(Fv/Fm)、电子传递速率ETR及非光化学淬灭系数NPQ。ΦPSⅡ=(Fm'-Fs)/Fm',NPQ=Fm/Fm'-1,ETR=ΦPSⅡ×PFD×0.5×0.84。净光合速率与叶绿素荧光参数同时测定。
1.2.5 二磷酸核酮糖羧化酶(RuBPCase)和磷酸烯醇式丙酮酸羧化酶(PEPCase)活力测定
根据Racker[17]的方法提取酶液,准确称取叶片0.5 g,置预冷冻过的研钵中,加少量预冷的酸洗石英砂和2.5 mL 提取液(0.1 mol/L Tris-HCl pH 值 8.4 缓冲液,内含10 mmol/L MgCl2、7 mmol/L β-巯基乙醇、1 mmol/L EDTA、5%甘油和1%PVP),冰浴下研磨至匀浆,倒入离心管,4℃下15000×g离心10 min,上清液即为酶提取液,测定样品为2009年试验材料。
RuBPCase活性测定的反应混合液共3.0 mL包括1 mmol/L Tris-HCl(pH 值 8.0)、0.1 mol/L MgCl2、50 mmol/L ATP、50 mol/L DTT、2 mmol/L NADH 和 1 mmol/L EDTA 各 0.3 mL,200 μmol/L NaHCO3溶液 0.1 mL,3-磷酸甘油酸激酶/3-磷酸甘油醛脱氢酶(15 U/15 U)溶液0.1 mL,蒸馏水0.8 mL 30℃恒温水浴10 min,加9 mmol/L的RuBP溶液0.1 mL,最后加酶提取液0.1 mL启动反应,立刻在340 nm下测定反应混合液吸光度的变化。PEPCase活性测定根据施教耐等[18]的方法,反应混合液共3.0 mL包括100 mmol/L Tris-HCl缓冲液(pH 值 9.2)1 mL、10 mmol/L MgCl2溶液 0.1 mL、10 mmol/L NaHCO3溶液 0.1 mL、1 g/L NADH 溶液 0.3 mL、50 U/mL苹果酸脱氢酶0.3 mL、蒸馏水和酶提取液各0.5 mL于28℃下水浴10 min,最后加40 mmol/L PEP溶液200 μL启动反应,立刻在340 nm下测定反应混合液吸光度的变化。
采用Microsoft Excel 2003和DPS 7.05软件进行数据处理和统计分析。
2 结果与分析
2.1 施氮时期对夏玉米叶面积指数(LAI)的影响
施氮显著提高夏玉米LAI,分次施氮较T2显著提高LAI。各处理VT时期LAI达到最大,开花后T5、T6 LAI显著高于T1、T2。DH661成熟期T6的LAI较T1、T2分别提高了12.6%和7.6%。ZD958成熟期T5的LAI较T1、T2分别提高了21.9%、8.1%。可见分次施氮延长了LAI高值持续期,延缓了叶片衰老(表2)。

表2 施氮时期对夏玉米叶面积指数的影响(2009)Table 2 Effects of nitrogen application stages on leaf area index of summer maize
2.2 施氮时期对功能叶光合色素含量的影响
由表3可以看出,施氮处理较T1显著提高叶绿素a和叶绿素(a+b)含量,开花后施氮处理间花后叶绿素b和类胡萝卜素含量的影响不显著。DH661和ZD958叶绿素a和叶绿素(a+b)含量均在花后14 d T4处理下达到最大,T4叶绿素a较T2分别提高了12.8%和15.4%,叶绿素(a+b)较T2分别提高了11.6%和16.4%。花后28 d两品种叶绿素(a+b)含量有所下降,在不同的施氮方式下功能叶的叶绿素(a+b)含量差异不显著,两品种变化趋势一致。
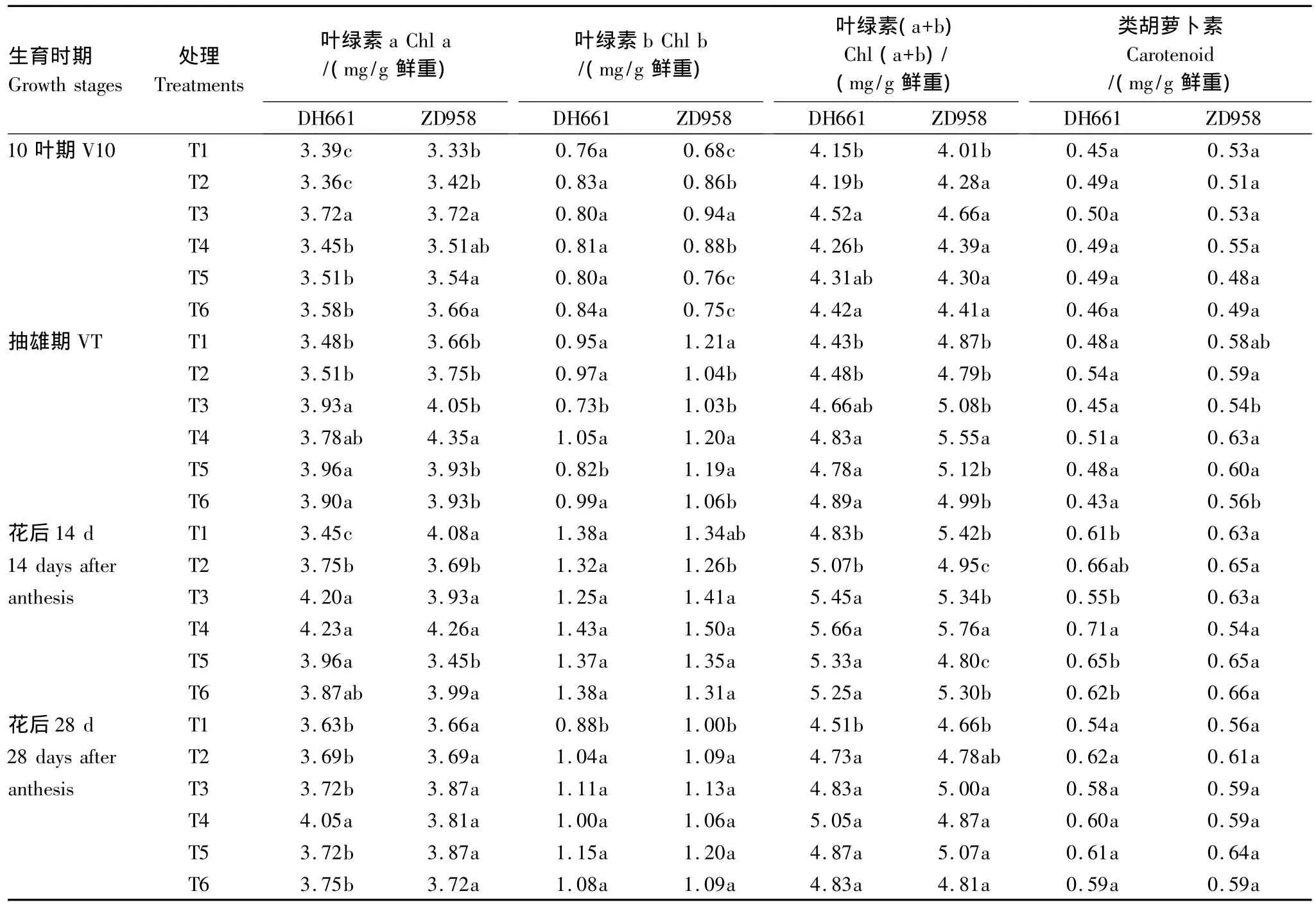
表3 施氮时期对功能叶光合色素的影响(2009)Table 3 Effects of nitrogen application stages on photosynthetic pigment content of ear leaves
2.3 施氮时期对夏玉米叶片净光合速率的影响
施氮显著提高了叶片净光合速率(Pn)。花后10 d T3和T6的Pn显著高于T1、T2;施氮后,花后24 d左右,DH661 的 T4 较 T1、T2 提高34.2%、16.0%;ZD958 的 T6 较 T1、T2 提高27.6%、27.5%。花后52 d 左右两品种T5和T6处理Pn显著高于T1、T2,DH661的T5处理较T1、T2平均提高16.8%、13.4%;ZD958的T5处理较 T1、T2 平均提高41.7%、33.2%(表4)。

表4 施氮时期对叶片净光合速率的影响/(μmol·m-2·s-1)Table 4 Effects of nitrogen application stages on net photosynthetic rate of ear leaves
2.4 施氮时期对夏玉米叶片叶绿素荧光参数的影响
分次施氮PSⅡ实际光化学效率(ΦPSⅡ)、非光化学猝灭系数(NPQ)及电子传递速率(ETR)较T1、T2显著提高,PSⅡ最大光化学效率(Fv/Fm)差异不显著,两品种变化趋势一致。T4、T5、T6处理ΦPSⅡ显著高于T1、T2。两品种T4、T5、T6处理NPQ和ETR显著高于T1、T2。NPQ在花后38 d左右,DH661的T4处理较T1、T2平均提高了75.0%、54.8%,ZD958的 T5处理较 T1、T2提高了70.6%、28.3%。ETR 在花后 38 d左右,DH661 和 ZD958 的 T5 较 T1、T2 平均提高了 49.5%、26.6%和19.9%、16.0%(表5)。

表5 施氮时期对叶片叶绿素荧光参数的影响Table 5 Effects of nitrogen application stages on chlorophyll fluorescence parameters of ear leaves
2.5 施氮时期对夏玉米叶片PEPCase和RuBPCase活性的影响
开花后PEPCase和RuBPCase活性呈先升高后降低的趋势。开花前分次施氮显著提高了开花期PEPCase和RuBPCase活性,开花期ZD958的T3处理PEPCase和RuBPCase活性显著高于T1、T2。花后施氮后PEPCase和RuBPCase活性显著提高。DH661在T5处理下,PEPCase和RuBPCase活性分别在花后28、14 d达到峰值,且显著高于其他处理,ZD958与 DH661的变化趋势一致。花后42 d,DH661在 T4、T5、T6下PEPCase活性显著高于 T1、T2,平均提高了83.8%和20.6%;T4、T5、T6下RuBPCase活性较 T1、T2平均提高了 26.6%和15.5%(表6)。
2.6 施氮时期对夏玉米产量和氮素积累利用的影响
分次施氮可以显著提高夏玉米籽粒产量及氮素在植株和籽粒中的积累。T2较T1增产不显著,平均2a试验结果DH661在T5处理产量最高,较T1、T2和T3分别平均增产15.7%、14.5%和8.3%;ZD958在T6处理产量最高,较T1、T2和T3分别平均增产21.6%、17.4%和6.4%。分次施氮同时提高了植株总氮素积累量和籽粒吸氮量,其中对于籽粒吸氮量的提高更加显著,从而提高了氮素收获指数,DH661和ZD958分别在T5和T4处理下籽粒吸氮量达到最大,较 T1、T2、T3分别平均提高61.8%、51.4%、36.4%和 54.9%、21.9%、12.2%。分次施氮较一次性施氮显著提高了氮肥偏生产力,DH661的T3、T4、T5较T2平均提高10.9%、14.4%、13.6%,ZD958 的 T3、T4、T5 较 T2 平均提高 15.6%、14.3%、17.5%(表7)。

表6 施氮时期对叶片PEPCase和RuBPCase活性的影响Table 6 Effects of nitrogen application stages on activities of PEPCase and RuBPCase in ear leaves

表7 施氮时期对夏玉米产量和氮素积累利用的影响Table 7 Effects of nitrogen application stages on grain yield and nitrogen use efficiency of summer maize
3 讨论
禾谷类作物生育后期的光合性能直接影响到籽粒产量[19]。施用氮素是调控作物生长和光合生产率的重要手段之一[4]。本试验在不改变总施氮量的前提下,通过调节不同的施氮时期及追施比例,研究了不同施氮方式对籽粒产量、氮素积累与利用、光合特性的影响。本研究结果表明,氮素分3—4追施较一次性施入可提高籽粒产量9.3%—18.2%,植株氮素积累量提高16.0%—28.6%,氮肥偏生产力提高10.9%—17.5%。
玉米冠层通过影响其内部的水、热、气等微环境影响着群体的光合效率和生物产量[20]。吕丽华等[2]研究指出,适当的施氮量可构建玉米高效合理的冠层结构。本研究表明,随花前施氮次数及施氮量的增加可提高抽雄期叶面积指数最大值,同时花后施氮延缓了花后植株下部叶片的衰老和脱落,延长了LAI高值持续期,在籽粒灌浆期保持了较高的光合面积。
前人研究指出[21-22],增施氮肥能提高小麦叶片叶绿素含量,增强PSⅠ和PSⅡ的电子传递能力,延长叶片的叶绿素含量缓降期和光合速率高值持续期,改善光合性能;也有研究指出施氮后灌浆期叶片叶绿素含量的变化不是Pn提高的主要原因,而两个光系统性能的改善及二者间协调性的提高增强了光合电子传递链的性能是灌浆期Pn升高与产量增加的主要原因[6]。本研究表明,施氮可提高叶片叶绿素含量,但不同施氮方式对叶绿素含量影响不显著,而分次施氮可显著提高 ΦPSⅡ;3∶5∶2,2∶4∶4 和 1∶2∶5∶2 的分次施氮方式较拔节期一次性施氮都可显著提高非光化学猝灭系数及电子传递速率,从而有效提高了叶片对光能的利用,同时增强了叶片对于光破坏的防御机制,有效调节了光合性能。
植物叶片吸收的光能被天线色素激发为电能后,主要以光化学反应、热耗散和荧光3种形式耗散;分配在光化学反应方向的光合电子流主要直接用于4个方面:碳同化、光呼吸、氧的还原反应(Mehler反应)和氮代谢[23-24]。本研究表明,两品种2∶4∶4分次施肥处理下,花后26 d净光合速率及花后14 d光合羧化酶活力均表现为低于3∶5∶2分次施肥处理,其原因在于花后施氮在促进光合电子羧化速率的同时提高了氧化速率,但供氮过多造成叶片氮代谢旺盛,光合产物的输出率降低,造成光合产物对光合器官的反馈抑制[25]。提高花后施氮比例后较多的呼吸耗能参与了氮代谢,但花后施氮20%—40%较拔节期一次性施氮可显著延长了光合速率高值持续期,同时PEPCase和RuBPCase活性分别提高了5.8%—12.8%和6.9%—17.8%。
本研究发现,登海661作为创造夏玉米高产纪录的新品种较目前第一大主推品种郑单958在开花后具有更好的叶片保绿性。开花后登海661较郑单958具有更高的LAI、Pn、ΦPSⅡ和光合羧化酶活力,在花后40 d和54 d登海661的Pn较郑单958平均提高了22.7%和17.4%;同时登海661较郑单958具有更强的叶片保护机制,花后40 d登海661的ΦPSⅡ较郑单958平均提高了11.3%。由此可见,在相同的施肥方式下登海661较郑单958具有更高的光合性能,这可能是其产量潜力更高的原因之一。
分次施氮有利于构建夏玉米合理冠层,提高了PSⅡ实际光化学效率及电子传递速率,增强了叶片保护机制,有效调节了光合性能。适当增加花后施氮比例有利于促进植株氮代谢的同时,延长了光合速率高值持续期,并使光合关键酶活性保持较高水平。因此,分次施氮显著提高了叶片的光合性能,进而提高了籽粒和氮素利用率。在实际生产中,高产田较一般生产田需要更多的肥料投入来获得高产,建议在高产地块控制施氮量在240—360 kg/hm2[26],根据两品种产量及花后光合特性登海661可采用2:4:4的施氮方式,郑单958可采用3∶5∶2 或 1∶2∶5∶2 的施氮方式。
[1] Mi G H,Chen F J,Chun L,Guo Y F,Tian Q Y,Zhang F S.Biological characteristics of nitrogen efficient maize genotypes.Plant Nutrition and Fertilizer Science,2007,13(1):155-159.
[2] Lü L H,Zhao M,Zhao J R,Tao H B,Wang P.Canopy structure and photosynthesis of summer maize under different nitrogen fertilizer application rates.Scientia Agricultura Sinica,2008,41(9):2624-2632.
[3] Wang J Z,Zhang C N,Zhao H J,Li F L,Yang Y J,Ma P F.Effects of different fertilization methods on chlorophyll fluorescence parameters and yield of summer maize.Plant Nutrition and Fertilizer Science,2008,14(3):479-483.
[4] Duan W W,Zhao H M,Guo C J,Xiao K,Li Y M.Responses of photosynthesis characteristics to nitrogen application rates in summer maize(Zea mays L.).Acta Agronomica Sinica,2007,33(6):949-954.
[5] Guan Y X,Lin B,Ling B Y.The interaction effects of growth light condition and nitrogen supply on maize(Zea mays L.)seedling photosynthetic traits and metabolism of carbon and nitrogan.Acta Agronomica Sinica,2000,26(6):152-158.
[6] Li G,Gao H Y,Liu P,Yang J S,Dong S T,Zhang J W,Wang J F.Effects of nitrogen fertilization on photosynthetic performance in maize leaf at grain filling stage.Plant Nutrition and Fertilizer Science,2010,16(3):536-542.
[7] Ma W Q,Li J H,Ma L,Wang F H,Sisák I,Cushman G,Zhang F S.Nitrogen flow and use efficiency in production and utilization of wheat,rice,and maize in China.Agricultural Systems,2009,99(1):53-63.
[8] Li H H,Ye Y L,Wang G L,Huang Y F.Wheat and corn production,fertilizer application and soil fertility status of typical high-yield areas.Henan Science,2009,27(1):59-63.
[9] Zhao B,Dong S T,Zhang J W,Liu P.Effects of controlled-release fertilizer on yield and nitrogen accumulation and distribution in summer maize.Acta Agronomica Sinica,2010,36(10):1760-1768.
[10] Müller P,Li X P,Niyogi K K.Non-photochemical quenching:a response to excess light energy.Plant Physiology,2001,125(4):1558-1566.
[11] Yu H,Yang G H,Wang Z J.Nitrogen rate and timing considerations on yield and physiological parameters of corn canopy.Plant Nutrition and Fertilizer Science,2010,16(2):266-273.
[12] Chen G Q,Qi W Z,Li Z,Wang J H,Dong S T,Zhang J W,Liu P.Analysis of hyperspectral on super high-yielding maize under different nitrogen levels.Acta Ecologica Sinica,2010,30(22):6035-6043.
[13] Wang C T,Li S K,Han B T.Approaches to high-yielding and yield potential exploration in corn.Science and Technology Review,2006,24(4):8-11.
[14] He Z F.Analysis Technique for Grain Quality of Cereals and Oils.Beijing:Agriculture Press,1985:31-41,57-59.
[15] Shi Y,Yu Z W,Wang D,Li Y Q,Wang X.Effects of nitrogen rate and ratio of base fertilizer and topdressing on uptake,translocation of nitrogen and yield in wheat.Acta Agronomica Sinica,2006,32(12):1860-1866.
[16] Arnon D I.Copper enzymes in isolated chloroplasts:polyphenoloxidase in Beta vulgaris.Plant Physiology,1949,24(1):1-15.
[17] Racker E.Ribulose diphophate carboxylase from spinach leaves//Colowick S P,Kaplan N O,eds.Methods in Enzymology.New York:Academic Press,1962,5:266-270.
[18] Shi J N,Wu M X,Cha J J.The study on PEPCase of plant Ⅰ.Comparison of PEPCase separating and aberrance structure characteristics.Journal of Plant Physiology,1979,5(3):225-236.
[19] Khan M N A,Murayama S,Ishimine Y,Tsuzuki E,Nakamura I.Physio-morphological studies of F1 hybrids in rice(Oryza sativa L.).Plant Production Science,1998,1(4):231-239.
[20] Li S K,Wang C T.The methods of obtaining and expressing information of crop plant shape and population structure.Journal of Shihezi University:Natural Science,1997,1(3):250-256.
[21] Lawlor D W.Carbon and nitrogen assimilation in relation to yield:mechanisms are the key to understanding production systems.Journal of Experimental Botany,2002,53(370):773-787.
[22] Yang Q,Li Y M,Xiao K,Du Y H.Effect of different amount of nitrogen on flag leaf senescence and yield components of wheat.Journal of Agricultural University of Hebei,2002,25(4):20-24.
[23] Evans J R,Poorter H.Photosynthetic acclimation of plants to growth irradiance:The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain.Plant,Cell and Environment,2001,24(8):755-767.
[24] Li H D,Gao H Y.Effects of different nitrogen application rate on allocation of photosynthetic electron flux in Rumex K-1 leaves.Journal of Plant Physiology and Molecular Biology,2007,33(5):417-424.
[25] Walker D A.Regulatory mechanism in photosynthetic carbon mechanism//Horecker B L,Stadtman E R.Current Topic in Cellular Regulation.New York:Academic Press,1976:230-241.
[26] Lü P,Zhang J W,Liu W,Yang J S,Su K,Liu P,Dong S T,Li D H.Effects of nitrogen application on yield and nitrogen use efficiency of summer maize under super-high yield conditions.Plant Nutrition and Fertilizer Science,2011,17(4):852-860.
参考文献:
[1] 米国华,陈范骏,春亮,郭亚芬,田秋英,张福锁.玉米氮高效品种的生物学特征.植物营养与肥料学报,2007,13(1):155-159.
[2] 吕丽华,赵明,赵久然,陶洪斌,王璞.不同施氮量下夏玉米冠层结构及光合特性的变化.中国农业科学,2008,41(9):2624-2632.
[3] 王俊忠,张超男,赵会杰,李付立,杨亚军,马培芳.不同施肥方式对超高产夏玉米叶绿素荧光特性及产量性状的影响.植物营养与肥料学报,2008,14(3):479-483.
[4] 段巍巍,赵红梅,郭程瑾,肖凯,李雁鸣.夏玉米光合特性对氮素用量的反应.作物学报,2007,33(6):949-954.
[5] 关义新,林葆,凌碧莹.光、氮及其互作对玉米幼苗叶片光合和碳、氮代谢的影响.作物学报,2000,26(6):152-158.
[6] 李耕,高辉远,刘鹏,杨吉顺,董树亭,张吉旺,王敬峰.氮素对玉米灌浆期叶片光合性能的影响.植物营养与肥料学报,2010,16(3):536-542.
[8] 李欢欢,叶优良,王桂良,黄玉芳.典型高产区小麦玉米产量、肥料施用及土壤肥力状况.河南科学,2009,27(1):59-63.
[9] 赵斌,董树亭,张吉旺,刘鹏.控释肥对夏玉米产量和氮素积累与分配的影响.作物学报,2010,36(10):1760-1768.
[11] 鱼欢,杨改河,王之杰.不同施氮量及基追比例对玉米冠层生理性状和产量的影响.植物营养与肥料学报,2010,16(2):266-273.
[12] 陈国庆,齐文增,李振,王纪华,董树亭,张吉旺,刘鹏.不同氮素水平下超高产夏玉米冠层的高光谱特征.生态学报,2010,30(22):6035-6043.
[13] 王崇桃,李少昆,韩伯棠.玉米高产之路与产量潜力挖掘.科技导报,2006,24(4):8-11.
[14] 何照范.粮油籽粒品质及其分析技术.北京:农业出版社,1985:31-41,57-59.
[15] 石玉,于振文,王东,李延奇,王雪.施氮量和底追比例对小麦氮素吸收转运及产量的影响.作物学报,2006,32(12):1860-1866.
[18] 施教耐,吴敏贤,查静娟.植物磷酸烯醇式丙酮酸羧化酶的研究 Ⅰ.PEP羧化酶同功酶的分离和变构特性的比较.植物生理学报,1979,5(3):225-236.
[20] 李少昆,王崇桃.作物株型和冠层结构信息获取与表述的方法(综述).石河子大学学报:自然科学学报,1997,1(3):250-256.
[22] 杨晴,李雁鸣,肖凯,杜艳华.不同施氮量对小麦旗叶衰老特性和产量性状的影响.河北农业大学学报,2002,25(4):20-24.
[24] 李海东,高辉远.不同施氮量对杂交酸模叶片光合电子流分配的影响.植物生理与分子生物学学报,2007,33(5):417-424.
[26] 吕鹏,张吉旺,刘伟,杨今胜,苏凯,刘鹏,董树亭,李登海.施氮量对超高产夏玉米产量及氮素吸收利用的影响.植物营养与肥料学报,2011,17(4):852-860.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
作物学报(2022年1期)2022-11-05
河北科技师范学院学报(2022年2期)2022-08-26
农业工程学报(2022年7期)2022-07-09
今日农业(2019年11期)2019-08-15
农业环境科学学报(2017年2期)2017-03-20
农家顾问(2014年9期)2014-10-20
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2012年2期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
