
小兴安岭4种典型阔叶红松林土壤有机碳分解特性
2013-09-11 08:38赵溪竹毛子军侯玲玲
生态学报 2013年2期
宋 媛,赵溪竹,毛子军,孙 涛,侯玲玲
(东北林业大学森林植物生态学教育部重点实验室,哈尔滨 150040)
土壤有机碳SOC(soil organic carbon)是陆地生态系统中最大的有机碳库,全球约有1400—1500 Pg碳以有机质形式储存于土壤中,是陆地植被碳库(500—600 PgC)的2—3倍[1-2]。土壤有机碳不仅为植被生长提供碳源、维持土壤良好的物理结构,同时也以CO2等温室气体的形式向大气释放碳[3]。森林土壤有机碳约占全球土壤有机碳的70%[4],其含量的微小变化都可能导致大气中CO2的浓度发生巨大变化[5]。
土壤有机质具有高度的复杂性和异质性,其组成成分包括易分解的微生物、新鲜的植物材料和在土壤中存在几千年的难分解物质[6],所以,用单一组分或一库来描述整个土壤的分解速率和周转周期是不合理的[7]。Parton等根据有机碳周转时间的不同,将其划分为活性碳库(5%,周转时间0.1—4.5 a),缓效性碳库(60%—85%,周转时间5—50 a)和惰性碳库(10%—40%,周转时间50—3000 a)[6,8],并且土壤有机碳循环动力学模型已经证明用三库一级动力学模型能够合理描述土壤有机碳的分解动态变化[9]。森林土壤有机碳不同碳库的准确评价是理解土壤碳循环机制和碳库动态变化的重要一步[10]。
小兴安岭林区是对全球变化敏感的高纬度地区,但是关于该地区土壤有机碳的绝大多数研究都集中在其碳库储存方面,而且多数是利用土壤普查数据和剖面分布图进行的区域尺度的土壤碳估算[11],基于不同群落类型的土壤有机碳含量的研究很少。此外,土壤有机碳不同组分周转时间各异,传统的研究中往往缺乏对土壤缓效性有机碳和惰性有机碳含量及周转速率的研究。因此,本研究以小兴安岭地区的地带性顶级生态系统(阔叶红松林)为研究对象,选取沿海拔梯度分布的4种不同阔叶红松林,其中云冷杉红松林为针叶林占优势,其他3种林分为针阔混交林,推测不同群落类型在土壤有机质的组成上会有不同,而且不同海拔梯度下每种林分的温度、湿度、以及其他环境条件可能会有不同,从而对土壤有机碳的含量、组成以及分解产生一定的影响。本项研究的目的是比较分析不同林分以及不同土层下的土壤有机碳分解速率的变化情况,并且最终量化表达土壤有机碳不同组分的含量以及周转速率。研究结果对于预测未来气候变化背景下中国温带地区针阔混交林生态系统的碳循环过程具有重要意义,并最终为正确理解陆地生态系统碳循环过程和准确评估碳排放提供科学依据。
1 研究地区与研究方法
1.1 研究地概况
实验样地设于伊春地区的丰林国家级自然保护区(129°06'—129°30'E,47°54'—48°19'N)。海拔 280—683 m,坡度多在10—25°之间,该区属于温带大陆性季风气候,四季分明,冬季寒冷干燥,夏季短而湿热,年平均温度-0.5℃,年平均降水量650 mm,无霜期约为120 d左右,地带性土壤为暗棕壤。该地区为小兴安岭典型地段,主要森林类型有阔叶红松林、云冷杉林、落叶松林、白桦天然次生林和针叶杂木混交次生林(针混林)等,地带性植被是以红松占优势的阔叶红松林。本实验以该区内4种典型阔叶红松林群落类型为研究对象:蒙古栎红松林(QP:Quercus mongolica-Pinus koraiensis forest)、枫桦红松林(BP:Betula costata-Pinus koraiensis forest)、椴树红松林(TP:Tilia amurensis-Pinus koraiensis forest)、云冷杉红松林(PAP:Picea sp.-Abies nephrolepis-Pinus koraiensis forest)。各样地的具体立地情况和植被组成见表1。

表1 不同群落类型样地概况Table 1 General situation of different forest sample plots
1.2 土壤样品的采集与处理
2008年8月上旬,在4个群落类型内布设30 m×30 m样方,在样方外距离四角2—3 m处及样方内的中心位置选定1 m×1 m土壤样方,挖取3个土壤剖面,剖面深度根据土壤发生层次而定(至母质C层为止),每一土层取约1 kg土样装入样品袋,带回室内,风干、研磨、过筛后进行土壤有机碳和化学性质测定分析。
1.3 实验方法
1.3.1 土壤有机碳和全氮含量测定
土壤惰性碳的测定采用酸水解法[12],即称取2 g过2 mm筛的风干土样于消煮管中,然后加入6 mol/L HCl并在115℃下消煮16 h,样品冷却后用蒸馏水洗至中性,再在55℃下烘干,研磨过180 μm筛,用重铬酸钾容量法-外加热法测得的有机碳即为惰性碳。土壤全氮的测定采用传统的半微量开氏法(K2SO4-CuSO4-Se蒸馏法)。
1.3.2 土壤有机碳释放速率的培养实验
采用室内恒温培养、碱液吸收法[10],即称取过0.95 mm风干土样100 g于玻璃广口瓶底部,广口瓶的体积为900 mL左右,调节含水量为田间持水量65%时的需水量(用质量法维持实验期间广口瓶中的土壤含水量),在小烧杯中加入25 ml 0.4 mol/L NaOH(保证在要求时间内能吸收完释放的CO2,并有少量盈余),之后将小烧杯放入广口瓶中,同时进行空白实验,然后密封放置在25℃的恒温培养箱内,黑暗条件下培养90 d。培养期间,定期通气,以保证微生物活动所需氧气。在一定时间间隔之后,将吸收液全部抽出并换上新的吸收液,再按上述方法继续培养。抽出的吸收液加20 mL的1 mol/L BaCl2,摇匀后以酚酞为指示剂,用0.4 mol/L HCl滴定中和未耗尽的NaOH,通过HCl消耗量来计算CO2的数量,进而求出有机碳的分解量。
1.3.3 凋落物现存量、有机碳含量、呼吸速率的测定
在4个样地(30 m×30 m)的四个角和中心设置5个5 m×5 m样方,在样方内随机选取1个0.5 m×0.5 m的小样方,收集该小样方内的地面凋落物,拣出石块和泥粒等,将样品装入样品袋中并编号,带回实验室,在65℃下烘至恒重后称重,计算其生物量。并将采集的植物样品在植物粉碎机中粉碎,通过0.3—0.5 mm的筛子,用分析天平称取植物样品0.02—0.03 g,放在25 cm×200 cm的硬质试管中。采用重铬酸钾容量法-外加热法测得有机碳。
凋落物呼吸的测定:在每个固定样地(30 m×30 m)内随机选择4个1 m×1 m的小样方,用壕沟隔断法去除凋落物与根系,其内部呼吸速率代表土壤微生物的呼吸速率。同时靠近每个小样方边缘再随机设置高6 cm(清除凋落物)和10 cm(有凋落物)的PVC土壤环各3个,形成1个区组。凋落物呼吸速率等于总呼吸速率减去样方外无枯枝落叶的土壤呼吸速率。具体的实验操作步骤参照陆斌等[13]的方法。
1.4 数据分析
模型拟合法[14,9]:本研究应用三库一级动力学模型Csoc=Cae-kat+Cse-kst+Cre-krt拟合土壤有机碳的分解动态,利用统计软件SAS8.2拟合出活性有机碳和缓效性有机碳含量及平均驻留时间MRT。式中Csoc为t时的土壤有机碳含量;Ca和Ka为活性碳有机碳含量和分解速率;Cs和Ks为缓效性有机碳含量和分解速率;Cr和Kr为惰性有机碳含量和分解速率。假设田间土壤惰性碳的平均驻留时间为1000a,MRTlab=MRTfield/Q10,Q10=2[(25-MRT)/],式中,Q10是温度系数,MAT为该地区的年平均温度,通过此方程可将田间平均驻留时间(MRTfield)转换为实验室平均驻留时间(MRTlab)。同时根据Kr=1/MRT可计算出惰性碳库的分解速率。利用90d培养数据,实验测得Csoc、Cr和假定的MRT,根据一级动力学方程,用SAS8.2中的非线性回归,经过循环迭代,迭代收敛,残差平方和最小,拟合出Ca、Ka和Ks3个参数的估计值。最后用Cs=Csoc-Ca-Cr,得到Cs。绘图采用origin8.0软件完成。
2 结果
2.1 土壤有机碳分解动态
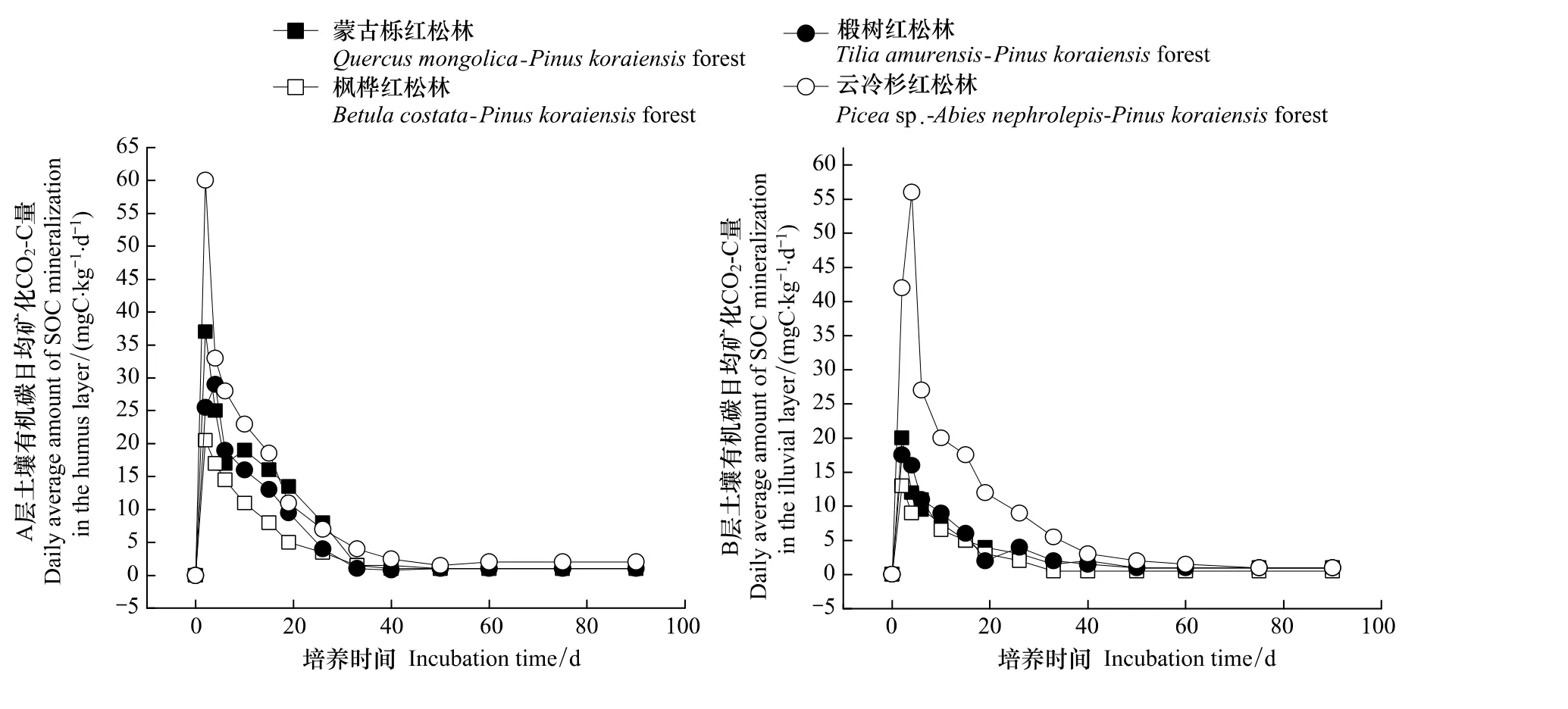
图1 实验室培养条件下小兴安岭地区阔叶红松林土壤有机碳释放速率动态Fig.1 Decomposition of SOC from four korean pine-broadleaved forest under Xiaoxing'an Regions in the laboratory
由图1可以看出,虽然在不同的阔叶红松林中,土壤有机碳的分解速率不同,但是其分解动态均呈现相似的变化规律,即在培养前期,分解迅速,之后缓慢下降并基本趋于稳定状态。A层(腐殖质层)土壤在培养实验进行的第2天,云冷杉红松林、蒙古栎红松林和枫桦红松林土壤有机碳的释放速率均达到最大,而椴树红松林的有机碳释放速率却仍在增加,并在第4天达到最大值。在实验进行至第50天时,各阔叶红松林的有机碳释放速率降至最低,均在1 mgC kg-1d-1左右。B层(淀积层)土壤有机碳的分解情况表现为:云冷杉红松林土壤有机碳释放速率在第4天达到最大,其值为56 mgC kg-1d-1,而其他3种林分在第2天就已经达到最大值,分别为:蒙古栎红松林(20 mgC kg-1d-1)、椴树红松林(17.5 mgC kg-1d-1)和枫桦红松林(13 mgC kg-1d-1)。可见各林分B层土壤的最大有机碳释放速率均小于A层土壤,除此之外,土壤有机碳的分解时间也长于A层,在培养的第75天才降至最小值,稍微滞后于A层。在开始培养的第一周内,A层的土壤有机碳分解量占总分解量的57%—62%,而在B层该值为61%—63%,由此可知土壤有机碳的分解主要集中在培养前期。
4种阔叶红松林不同土壤层有机碳分解规律不同,在A层,不同阔叶红松林土壤有机碳分解速率的大小顺序为:云冷杉红松林>蒙古栎红松林>椴树红松林>枫桦红松林,且蒙古栎红松林和椴树红松林之间没有表现出太大的差异,但是在B层,云冷杉红松林的分解速率明显大于其他3种林分,而这3种林分的土壤有机碳分解速率却没有表现出明显不同。
2.2 土壤有机碳各组分含量及周转情况
2.2.1 土壤有机碳各组分含量
本研究表明(表2),4种林分A层SOC的含量在86.87—166.14 g/kg之间,B层SOC在23.92—148.28 g/kg之间。活性碳的含量分别在0.77—2.95 g/kg之间和0.69—2.75 g/kg之间,其占总有机碳的0.89%—1.78%和1.91%—2.87%。缓效性碳的含量分别在 19.87—37.51 g/kg 和7.76—35.39 g/kg 之间,占总有机碳的22.58%—28.44%和23.87%—42.63%。惰性碳的含量分别在 66.23—125.67 g/kg 和 13.04—110.05 g/kg之间,占总有机碳的69.98%—76.24%和54.50%—74.22%。在4种阔叶红松林中,云冷杉红松林在A层或B层,土壤活性碳、缓效性碳及惰性碳的含量均是最大的,然而位于最高海拔的蒙古栎红松林,其土壤有机碳各组分的含量却是最小的,且与枫桦红松林没有太大差异,而椴树红松林的土壤总有机碳及各组分的含量仅次于云冷杉红松林。虽然A层的活性碳和缓效性碳含量大于B层,但是其各自占总有机碳的比例却是A层小于B层。
2.2.2 土壤有机碳各组分在土壤剖面的垂直分布
不同阔叶红松林中,土壤有机碳各组分的分布情况虽然不同,但有机碳各组分在A层的含量始终大于B层,而且其随土层加深而下降的幅度也有不同,其B层相对于A层下降的幅度分别为,活性碳:蒙古栎红松林(10.4%)、枫桦红松林(8.9%)、椴树红松林(5.0%)、云冷杉红松林(6.8%),缓效性碳:蒙古栎红松林(48.5%)、枫桦红松林(64.0%)、椴树红松林(6.1%)、云冷杉红松林(5.7%),惰性碳:蒙古栎红松林(80.3%)、枫桦红松林(81.0%)、椴树红松林(39.4%)、云冷杉红松林(12.4%)。由土壤有机碳各组分的下降程度可知,云冷杉红松林和椴树红松林的土壤有机碳在土壤剖面中分布较均匀,而蒙古栎红松林和枫桦红松林的土壤有机碳在土壤剖面分布的稳定性较差。此外,在整体情况下,惰性碳下降最明显,缓效性碳次之,活性碳最小。
2.2.3 土壤有机碳各组分周转情况
由表2可知,4种阔叶红松林土壤有机碳各组分的驻留时间的变化情况与其含量的分布情况一致,即云冷杉红松林>枫桦红松林>椴树红松林>蒙古栎红松林,且在A层的驻留时间小于B层,由此进一步说明各阔叶红松林土壤A层有机碳的分解速率大于B层。4种阔叶红松林土壤活性碳在A层和B层的驻留时间分别在12—35 d和27—58 d之间,其中枫桦红松林和椴树红松林的驻留时间差异不大。蒙古栎红松林土壤活性碳的驻留时间在两个土层之间相差37 d,而枫桦红松林却只相差8 d。缓效性碳库在A层和B层的驻留时间分别为4—19 a和16—37 a。枫桦红松林土壤缓效性碳的驻留时间在A土层和B土层之间相差21 a,相差时间最少的为椴树红松林,只有7 a。根据公式可计算出土壤惰性碳的驻留时间为171 a。
3 讨论
3.1 土壤有机碳分解动态及其影响因素
土壤有机碳的分解主要集中在培养前期,说明土壤有机碳在组成成分上是较复杂的,既有易分解的组分,也有很难分解的组分,所以将土壤有机碳分为易被微生物分解利用的活性碳和理化性质稳定极难分解的惰性碳以及介于两者之间的缓效性碳是合理的。在培养早期,分解的SOC主要来源于由大部分植物残体、相当数量的微生物和周转迅速的微小动物碎片等组成的活性碳[15],这些物质提供给微生物充足的营养物质,使得微生物活动剧烈,分解速率较快。但是在培养后期,难分解的木质素和纤维素等成分的有机碳占优势,释放给微生物的能源物质缓慢,微生物活动也相应缓和[7]。在培养实验进行的早期,由于土壤有机质中的活性部分的分解,导致CO2通量迅速下降,之后稳定性介于活性碳和惰性碳之间的有机碳的分解作用占优势,导致CO2的释放速率逐渐趋于稳定[6]。Yang等[10]也发现同样的规律。Zhang等[16]在中国南方3种水稻土的表层土壤有机碳的矿质化作用和CO2释放的研究中也发现在114 d的培养实验中,有机碳的矿质化作用可以分成明显不同的3个阶段。在不同群落类型中,A层土壤有机碳的释放速率大于B层,这可能是因为森林的枯枝落叶层不但为土壤层提供大量有机碳,而且使表层土壤具有较高的养分浓度和较好的水分条件,从而为植物细根向表土层聚集提供了良好的条件[17],而根系分泌物和死亡的根可以为微生物提供能源物质[18]。夏汉平等[19]的研究发现在同一土壤剖面,表层的养分含量和微生物数量都明显高于中下层。在半干旱地区的土壤酶活性的剖面分布大多随着深度的增加而降低[20]。在生长状况不同的云冷杉林中,其土壤酶活性和土壤养分含量均随土层深度的增加而降低[21]。所以,可能是由于土壤酶以及养分在土壤剖面中的不同分布导致土壤有机碳分解速率发生差异。
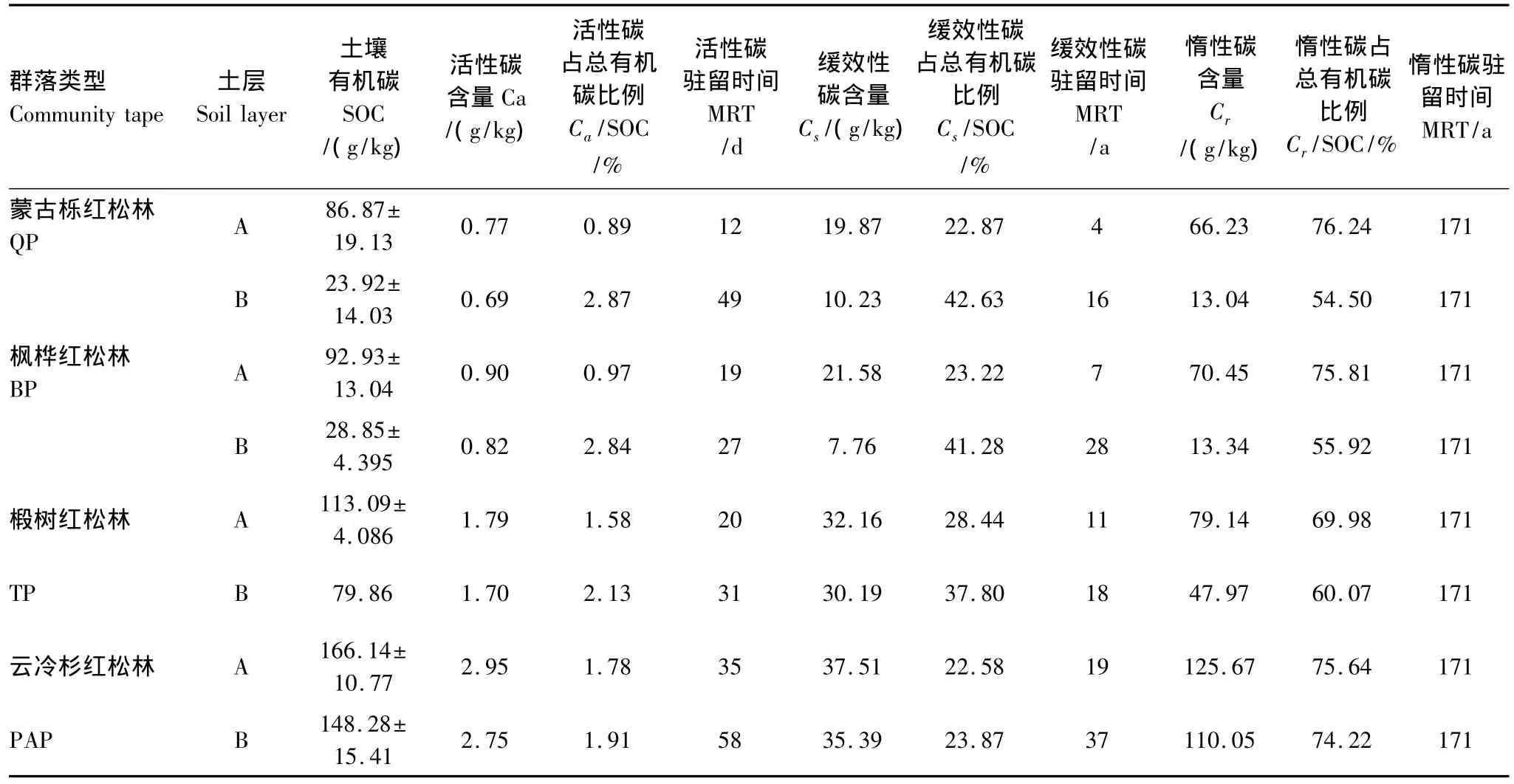
表2 不同阔叶红松林土壤活性、缓效性和惰性有机碳含量及驻留时间Table 2 Pool sizes and laboratory mean residence times(MRT)of active,slow and resistant soil organiccarbon pools under different korean pine-broadleaved forest
研究结果表明,在土壤A层,4种阔叶红松林土壤有机碳分解速率的大小顺序为:云冷杉红松林>蒙古栎红松林>椴树红松林>枫桦红松林,在B层,云冷杉红松林的分解速率明显大于其他3种林分,而这3种林分的土壤有机碳分解速率却没有表现出明显不同。因为土壤孵化实验是在恒定温度和湿度下进行的,温度和湿度不再是影响有机碳分解的主要因素[7]。有研究认为,在实验室培养条件下,不同群落间土壤有机碳分解动态的差异可能与土壤有机碳的初始含量、C/N比和土壤的N含量不同有关[22],而且C/N相对较高的林地土壤,抗分解能力强,有利于土壤有机碳的累积[23]。Côté等[24]认为不同森林类型土壤有机碳矿质化作用速率的不同是由活性碳库引起的,为此,进行了土壤有机碳的最大分解速率与土壤有机碳的初始含量、活性碳含量、C/N比及土壤的N含量的相关性分析,结果表明:土壤有机碳的最大分解速率与土壤活性碳及总有机碳的含量均有一个极显著的相关关系(P<0.01)(图2);在4种阔叶红松林中,无论在土壤A层还是B层,云冷杉红松林的土壤全氮含量是最多的,分别为0.68%和0.56%,而蒙古栎红松林的土壤全氮量却只有0.36%和0.14%(表3);在C/N方面,云冷杉红松林最高,A层的C/N是24.35,除与蒙古栎红松林差异不显著外,与其他两种林分的C/N比差异显著,各林分C/N的大小顺序为:云冷杉红松林>蒙古栎红松林>椴树红松林>枫桦红松林,与A层土壤有机碳的分解速率的变化情况一致。云冷杉红松林B层的C/N也是最大的,与其余3种林分差异显著,而后3种红松林的C/N之间没有显著差异,这与B层土壤有机碳分解速率的动态变化规律也是相一致的。土壤有机碳最大释放速率与C/N的相关分析结果表明,两者之间有一个极其显著的相关关系(P=0.009)(图2),说明土壤较高的C/N值可能是影响土壤中有机碳滞留的重要因素[25],而且其矿质化作用的强度是受基质的性质决定的[26]。徐侠等[27]在武夷山不同海拔高度土壤活性有机碳变化的研究中发现土壤有效碳与土壤全氮之间有一个极显著的相关关系,而且不同海拔高度群落类型的土壤有机碳释放速率的变化规律与其土壤中的C/N分布情况一致。该结论与本实验的研究结果一致,此外,在姜培坤[28]的研究中也发现土壤活性碳与土壤全氮之间表现出很好的相关性,由此进一步说明土壤有机质中氮的含量会影响微生物对其分解和利用的速率。
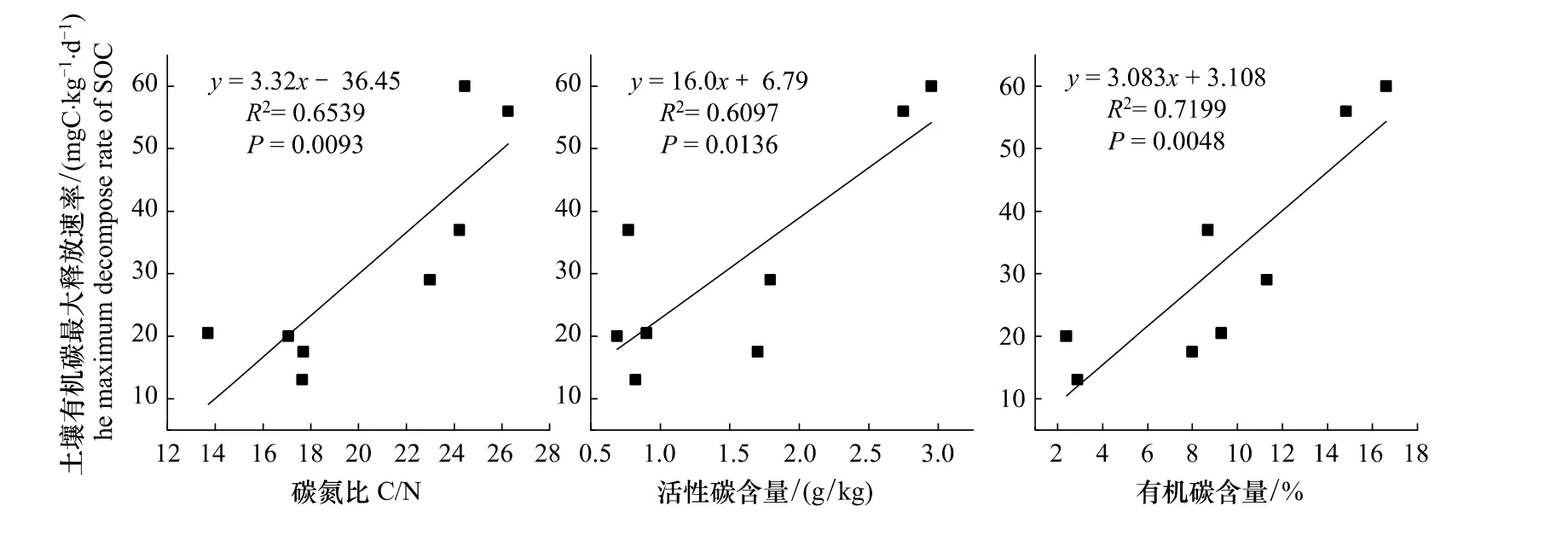
图2 土壤有机碳释放速率与碳氮比、有机碳含量、活性碳含量的相关关系Fig.2 Correlation between decomposition rate of SOC with C/N,active carbon,soil carbon organic
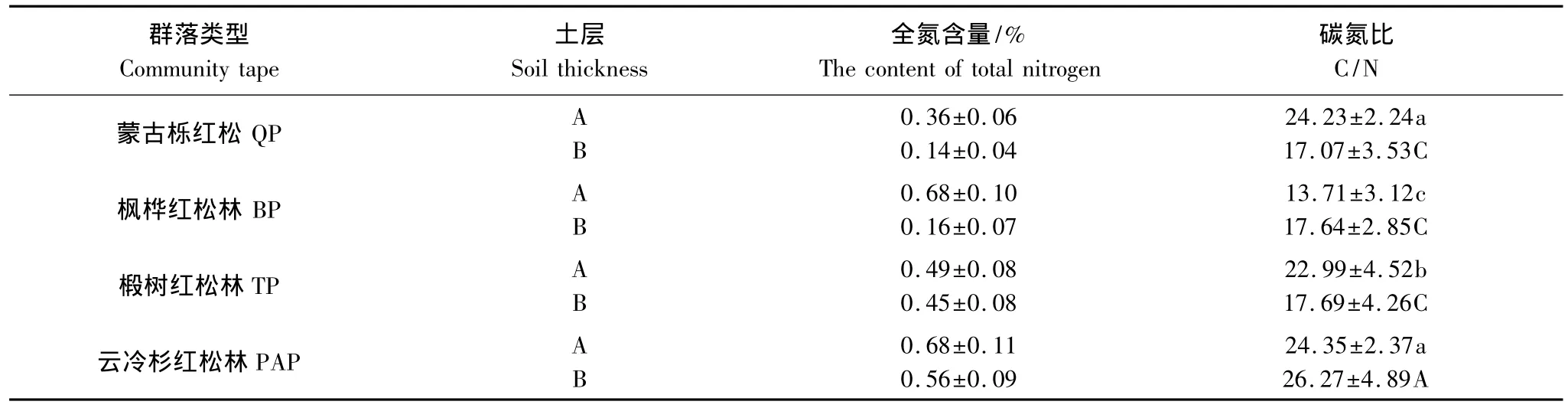
表3 不同红松林土壤全氮含量及碳氮比Table 3 Total nitrogen and SOC/TN under different Pinus koraiensis forests
植物群落间树种组成的差异决定了各群落凋落物的化学属性,如木质素、氮元素、C/N、木质素/N等不同,而凋落物的上述特性决定了供分解者分解的土壤有机质的性质[29-31]。凋落物作为外源有机物质,由易分解成分(如糖类、淀粉、脂肪等)和难分解成分(木质素、多酚等)组成,进入土壤后其分解不但会增加土壤中的碳源,改变土壤理化性质,还会增加土壤中各种微生物的数量,改变土壤微生物群落性状,从而对土壤有机碳矿化产生重要影响。凋落物自身分解特征与土壤有机碳的分解特征相同,均是前期迅速,后期缓慢[32]。并且有研究表明在凋落物分解的初期,凋落物分解速率的差异是由凋落物中易分解有机物的含量决定的,而与凋落物中的总有机碳含量、全氮含量及C/N比等没有直接的关系[33]。王清奎等[34]的研究发现添加凋落物显著增加了土壤中可溶性有机碳含量。虽然在本研究中,4种阔叶红松林的凋落物生物量及有机碳含量均没有表现出显著的差异性(P>0.05)(表4),但是仍然可以看出其大致的变化趋势,即与土壤有机碳含量的变化情况一致(表3),由此说明其与有机碳的分解有一定的联系。对4种林分的凋落物呼吸测定结果(表4)进一步说明了凋落物的化学性质对土壤有机碳的分解具有非常重要的作用。史学军等[35]也发现麻栎、马尾松和青冈栎3种凋落物对土壤有机碳矿化的贡献率都在80%以上。

表4 不同红松林凋落物有机碳含量、现存量和呼吸作用Table 4 Biomass and organic and respiration of litter under different Pinus koraiensis forests
3.2 土壤有机碳各组分含量
4种阔叶红松林土壤A层和B层的活性碳分别占总有机碳的0.89%—1.78%和1.91%—2.87%,平均驻留时间为12—35 d和27—58 d。缓效性碳占总有机碳的22.58%—28.44%和23.87%—42.63%,平均驻留时间为4—19 a 和18—37 a。惰性碳占总有机碳的69.98%—76.24%和54.50%—74.22%。Iqbal等[25]发现,林地中的土壤活性有机碳占总有机碳的0.84%,平均驻留时间为49 d,缓效性碳占总有机碳的25.27%,平均驻留时间为27 a,惰性碳占总有机碳的73.88%,与本研究的结论相符。此外,Yang等[10]在中国温带和亚热带森林的土壤有机碳分解和碳库研究中得出如下结论,即在长白山样地的0—12 cm土层中的土壤活性碳占总有机碳的0.93%,平均驻留时间为11 d,缓效性碳占总有机碳的25.20%,平均驻留时间为4 a,惰性碳占总有机碳的73.87%,平均驻留时间为250 a。邵月红等[7]也对长白山地区不同森林植被下土壤有机碳各组分的含量及周转速率进行研究,发现针阔混交林下的暗棕壤的土壤有机碳含量在11.90—103.37 g/kg之间,活性碳含量在0.22—0.55 g/kg之间,平均驻留时间为17—60 d,缓效性碳含量在8.23—31.90 g/kg之间,平均驻留时间为9—49 a,惰性碳含量在3.44—70.91 g/kg之间。
无论在土壤A层,还是土壤B层,4种阔叶红松林土壤总有机碳和活性有机碳含量的大小顺序均为:云冷杉红松林>椴树红松林>枫桦红松林>蒙古栎红松林,B层的Ca/Ct和Cs/Ct大于A层。一般情况下,B层的土壤有机碳含量较少,主要是由于其来源于少量植物细根、根毛及根系分泌物,或者是从土壤表层下移的较精细的有机质(可能是腐殖质),较难分解。而在本研究中,椴树红松林和云冷杉红松林的土壤B层有机碳含量偏高,但是A层和B层土壤之间的有机碳含量表现出不同,所以本文中所谈及的B层很可能是位于土壤A层下缘和B层上缘之间的土层。在B层,云冷杉红松林的活性碳及缓效性碳占总有机碳的比例均比其他林分小,而且,活性碳和缓效性碳的驻留时间也是最长的,除此之外,在4种林分中,云冷杉红松林的惰性碳占总有机碳的比例是最大的。这与云冷杉红松林分布地的树种组成及地形特点有关,在组成上,该林分主要组成树种均为常绿针叶树种,凋落叶中难分解的木质素等成分较阔叶树种多[36],在生境上,该林分位于半阴坡下部,地形平缓,土壤温度较其他3种红松林稍低,土壤含水量较高,形成了相对冷湿的环境,所以有机物质容易积累,而且分解较慢。因此,云冷杉红松林的土壤有机碳较其他林分稳定。周玉荣等[37]的研究发现云冷杉林下的土壤呼吸较弱,形成巨大的土壤碳贮库,是我国森林主要的碳库。然而位于最高海拔的蒙古栎红松林,其土壤总有机碳以及有机碳各组分的含量均是最少的。首先,这可能与蒙古栎的生态习性有关,在自然景观里,因为蒙古栎对湿度、土壤条件有着比其他大多数森林树种更大的生态耐性或生态幅度,所以其只能在极端的或贫瘠的土壤上才能保持地位[38]。栎树比较耐旱,在次生林区的冈脊部位,尤其经反复破坏和火烧后,是诸立地类型中最为干旱瘠薄的一类,在这样的立地条件下,惟有栎树最能适应,在短时间内其他树种不可能更替它[39]。在样地的基本情况调查中,也发现蒙古栎红松林的土层厚度是最薄的只有30—40 cm。其次,蒙古栎红松林样地位于阳坡上部陡坡,其他3种红松林样地均位于半阴坡的缓坡和平地。在坡度较大的情况下,凋落物以及土壤中的有机质容易被雨水带走,不利于有机碳的累积。何志斌等[40]研究发现阴坡的土壤有机碳含量高于半阴坡和阳坡。此外,渠开跃等[41]在辽东山区不同林型土壤有机碳剖面分布特征及碳储量研究中也发现,坡度较大的蒙古栎林地的土壤有机碳含量是最少的。
不同阔叶红松林土壤活性碳、缓效性碳和惰性碳的含量都是A层>B层,但是其各自占总有机碳的比例却是B层>A层,这说明森林植被的凋落物主要影响了A层土壤,使A层土壤的总有机碳含量明显提高,虽然也使活性碳、缓效性碳的含量有所提高,但它主要是增加了总有机碳中的惰性碳含量,从而使得A层土壤的活性碳、缓效性碳分别占总有机碳的比例小于B层土壤。这与邵月红等[7]在长白山森林土壤有机碳库大小及周转的研究中得到的结果一致。渠开跃等[41]也认为由于土壤表层地表枯落物较为丰富,土壤有机质较为丰富,使得表层有机碳含量较高,而在亚表层、底层受地表枯落物的影响较小,有机质相对表层低,因此导致亚表层、底层有机碳含量低于表层。
活性碳、缓效性碳和惰性碳均与土壤总有机碳有一个显著的相关关系(P<0.01),各自相关程度的大小依次为:Cr(R2=0.990,P=0.000)>Cs(R2=0866,P=0.01)>Ca(R2=0.771,P=0.004)。其中活性碳的相关性最小,这是因为在一定的空间范围内,土壤活性碳极易受环境条件变化的影响,易氧化分解,而且由于土壤较高的背景值和较大的空间异质性,总土壤有机碳的微小变化很难被发现,所以活性碳是用来指示土壤有机碳动态变化的一个较有用的指标[42]。在本研究中,土壤惰性碳随土层深度加大而下降的最明显,而活性碳的下降程度最小。土壤有机碳在土壤剖面表现出下降规律,这与土壤有机碳主要来源于凋落物有关,然而,活性碳含量的下降不仅与土壤总有机碳随土层深度的增加而下降有关,还与B层较高的粘粒含量密切相关[43]。然而Elzein等[44]却发现虽然土壤总有机碳含量随土层深度增加呈现快速的指数下降趋势,但是,性质稳定的土壤有机碳在土壤剖面中却是均匀分布的。这可能与土壤类型和土壤剖面的具体特点有关。
4 结论
实验室培养条件下,土壤有机碳的分解主要集中在培养前期,分解的土壤有机碳的57%—63%是在开始培养的第一周内完成的;4种阔叶红松林腐殖质层土壤有机碳的分解速率均大于淀积层,但是不同林分之间却有差异:在腐殖质层,云冷杉红松林>蒙古栎红松林>椴树红松林>枫桦红松林,但蒙古栎红松林和椴树红松林之间差异不明显,淀积层,云冷杉红松林的分解速率明显大于其他3种林分,但是其他3种林分间差异不明显。土壤有机碳分解速率与土壤总有机碳、活性碳、及土壤的C/N呈正相关。
不同林分土壤腐殖质层的总有机碳、活性碳、缓效性碳和惰性碳的含量明显大于淀积层,但是3种土壤有机碳占总有机碳的比例却是腐殖质层小于淀积层,在4种红松林中,云冷杉红松林的土壤总有机碳、活性碳、缓效性碳及惰性碳含量最大,相比之下位于高海拔地带的蒙古栎红松林却是最小的,即基于群落类型的土壤有机碳含量的多少与其各自所处的地理位置有很大的关系。凋落物主要增加了土壤表层的有机碳含量,而且主要是增加了惰性碳的含量。
各群落土壤腐殖质层活性碳和缓效性碳的平均驻留时间短于淀积层的平均驻留时间。
[1] Su Y Z,Zhao H L.Advances in researches on soil organic carbon storages,affecting factors and its environmental effects.Journal of Desert Research,2002,22(3):220-228.
[2] Schlesinger W H.Evidence from chronosequence studies for a low carbon-storage potential of soils.Nature,1990,348(6298):232-234.
[3] Zhou L,Li B G,Zhou G S.Advances in controlling factors of soil organic carbon.Advances in Earth Science,2005,20(1):99-105.
[4] Pan G X,Li L Q,Zhang X H.Perspectives on issues of soil carbon pools and global change—With suggestions for studying organic carbon sequestration in paddy soils of China.Journal of Nanjing Agricultural University,2002,25(3):100-109.
[5] Raich J W,Schlesinger W H.The global carbon dioxide flux insoil respiration and its relationship to vegetation and climate.Tellus,1992,44B(2):81-99.
[6] Townsend A R,Votousek P M,Trumbore S E.Soil organic matter dynamics along gradients in temperature and land use on the island of Hawaii.Ecology,1995,36(3):721-723.
[7] Shao Y H,Pan J J,Xu X W,Yang L X.Determination of forest soil organic carbon pool sizes and turnover rates in Changbaishan.Journal of Soil and Water Conservation,2006,20(6):99-102.
[8] Parton W J,Schimel D S,Cole C V,Ojima D S.Analysis of factors controlling soil organic matter levels in great plains grasslands.Soil Science Society of America Journal,51(5):1173-1179.
[9] Collins H P,Christenson D R,Blevens R L,Bundy L G,Dick W A,Hunggisand D R,Paul E A.Soil carbon dynamics in corn-based agroecosystems:Results from13C natural abundance.Soil Science Society of America Journal,1999,63(3):584-591.
[10] Yang L,Pan J,Shao Y,Chen J M,Ju W M,Shi X,Yuan S.Soil organic carbon decomposition and carbon pools in temperate and sub-tropical forests in China.Journal of Environmental Management,2007,85(3):690-695.
[11] Yang J Y,Wang C K.Soil carbon storage and flux of temperate forest ecosystems in northeastern China.Acta Ecologica Sinica,2005,25(11):2875-2882.
[12] Leavitt S W,Paul E A,Pendall E,Pinter P J Jr,Kimball B A.Field variability of carbon isotopes in soil organic carbon.Nuclear Instruments and Methods in Physics Research,1997,123(1/4):451-454.
[13] Lu B,Wang S H,Mao Z J,Sun T,Jia G M,Jin S B,Sun P F,Cheng C X.Soil respiration characteristics of four primary Korean pine communities in growing season at Xiaoxing'an mountain,China.Acta Ecologica Sinica,2010,30(15):4065-4074.
[14] Leavitt S W,Foollett R F,Paul E A.Estimation of slow-and fast-cycling soil organic carbon pools from 6N HCl hydrolysis:14C and soil dynamics:special section=Estimation des cycles lents et rapides des zones de carbone d'origine organique d'hydrolyse de 6N HCl.Radiocarbon,1996,38(2):231-239.
[15] Gody S,Phillip S,Sharon R.Carbon and nitrogen in the light fraction of a forest soil:vertical distribution and seasonal patterns.Soil Science,1983,135(2):79-87.
[16] Zhang X H,Li L Q,Pan G X.Topsoil organic carbon mineralization and CO2evolution of three paddy soils from South China and the temperature dependence.Journal of Environmental Sciences,2007,19(3):319-326.
[17] Hendrick R L,Pregitzer K S.Temporal and depth-related patterns of fine root dynamics in northern hardwood forests.Journal of Ecology,1996,84(2):167-176.
[18] Smith J L,Paul E A.The significance of soil microbial biomass estimations//Bollag J M,Stotzky G,eds.Soil Biochemistry.New York:Marcel Dekker,1991:359-396.
[19] Xia H P,Yu Q F,Zhang D Q.The soil acidity and nutrient contents and their characteristics of seasonal dynamic changes under 3 different forests of dinghushan nature reserve.Acta Ecologica Sinica,1997,17(6):645-653.
[20] Ding H,Hu H B,Wang R C.The relationships between soil enzyme activity and soil physical-chemical properties or microbial biomass in semi-arid area.Journal of Nanjing Forestry University:Natural Sciences Edition,2007,31(2):13-18.
[21] Chen L M,Man X L.Soil enzymes activities and soil nutrient of Picea and Abies forest.Science of Soil and Water Conservation,2009,7(4):94-99.
[22] Wang Q K,Wang S M,Yu X J,Zhang J,Liu Y X.Soil carbon mineralization potential and its effect on soil active organic carbon in evergreen broadleaved forest and Chinese firplantation.Chinese Journal of Ecology,2007,26(12):1918-1923.
[23] Jiang Y,Zhang Y G,Liang W J,Wen D Z.Profile distribution and storage of soil organic carbon in an aquic brown soil as affected by land use.Scientia Agricultura Sinica,2005,38(3):544-550.
[24] Côté L,Brown S,Paré,D,Fyles J,Bauhus J.Dynamics of carbon and nitrogen mineralization in relation to stand type,stand age and soil texture in the boreal mixedwood.Soil Biology and Biochemistry,2000,32(8/9):1079-1090.
[25] Iqbal J,Hu R G,Lin S,Ahamadou B,Feng M L.Carbon dioxide emissions from Ultisol under different land uses in mid-subtropical China.Geoderma,2009,152(1/2):63-73.
[26] Diaz-Raviña M,Carballas T,Acea M J.Microbial biomass and metabolic activity in four acid soils.Soil Biology and Biochemistry,1988,20(6):817-823.
[27] Xu X,Chen Y Q,Wang J S,Fang Y H,Quan W,Ruan H H,Xu Z K.Variations of soil labile organic carbon along an altitude gradient in WuyiMountain.Chinese Journal of Applied Ecology,2008,19(3):539-544.
[28] Jiang P K.Soil active carbon pool under different types of vegetation.Scientia Silvae Sinicae,2005,41(1):10-13.
[29] Berg B,Mcclaugherty C.Nitrogen release from litter in relation to the disappearance of lignin.Biogeochemistry,1987,4(3):219-224.
[30] Taylor B R,Parkinson D,Parsons W F J.Nitrogen and lignin content as predictors of litter decay rates:a microcosm test.Ecology,1989,70(1):97-104.
[31] Riffaldi R,Saviozzi A,Levi-Minzi R.Carbon mineralization kinetics as influenced by soil properties.Biology and Fertility of Soils,1996,22(4):293-298.
[32] Li Y Q,Xu M,Sun O J,Cui W C.Effects of root and litter exclusion on soil CO2efflux and microbial biomass in wet tropical forests.Soil Biology and Biochemistry,2004,36(12):2111-2114.
[33] Wu Q B,Wang X K,Ouyang Z Y.Effects of labile organic carbon on the litters decomposition process.Ecology and Environment,2006,15(6):1295-1299.
[34] Wang Q K,Wang S L,Yu X J,Zhang J,Liu Y X.Effects of Cunninghamia lanceolata-broadleaved tree species mixed leaf litters on active soil organic matter.Chinese Journal of Applied Ecology,2007,18(6):1203-1207.
[35] Shi X J,Pan J J,Chen J S,Yang Z Q,Zhang L M,Su B,Li P Z.Effects of different types of litters on soil organic carbon mineralization.Chinese Journal of Environmental Science,2009,30(6):1832-1837.
[36] Li X F,Han S J,Guo Z L,Zheng X B,Song G Z,Li K X.Decomposition of pine needles and twigs on and under the litter layer in the natural Korean pine broadleaved forests.Journal of Beijing Forestry University,2006,28(3):8-13.
[37] Zhou Y R,Yu Z L,Zhao S D.Carbon storage and budget of major Chinese forest types.Acta Phytoecologica Sinica,2000,24(5):518-522.
[38] Gao Z T,Wu X C.Discussion on regulation on geographic distribution in Mengguli.Protection Forest Science and Technology,2005,(2):83-84.
[39] Chen D K,Zhou X F,Zhao H X,Wang Y H,Jin Y Y.Study on the structure,function and succession of the four types in natural secondary forest.Journal of Northeast Forestry University,1982,(2):1-20.
[40] He Z B,Zhao W Z,Liu H,Su Y Z.Characteristic of picea crassifolia forest soil organic carbon and relationship with environment factors in the Qilian Mountain.Acta Ecologica Sinica,2006,26(8):2572-2577.
[41] Qu K Y,Feng H M,Dai L M,Zhou L.Profile distribution and storage of soil organic carbon of main forest types in eastern mountainous region of Liaoning.Chinese Journal of Soil Science,2009,40(6):1316-1320.
[42] Powlson D S,Jenkinson D S.A comparison of the organic matter,biomass,adenosine triphosphate and mineralizable nitrogen contents of ploughed and direct-drilled soils.The Journal of Agricultural Science,1981,97(3):713-721.
[43] Geng Y Q,Yu X X,Yue Y J,Li J H,Zhang G Z,Liu S.Soil active organic carbon pool of coniferous and broadleaved forest in the mountainous area of Beijing.Journal of Beijing Forestry University,2009,31(5):19-24.
[44] Elzein A,Balesdent J.Mechanistic simulation of vertical distribution of carbon concentrations and residence times in soils.Soil Science Society of America Journal,1994,59(5):1328-1335.
参考文献:
[1] 苏永中,赵哈林.土壤有机碳储量、影响因素及其环境效应的研究进展.中国沙漠,2002,22(3):220-228.
[3] 周莉,李保国,周广胜.土壤有机碳的主导影响因子及其研究进展.地球科学进展,2005,20(1):99-105.
[4] 潘根兴,李恋卿,张旭辉.土壤有机碳库与全球变化研究的若干前沿问题——兼开展中国水稻土有机碳固定研究的建议.南京农业大学学报,2002,25(3):100-109.
[7] 邵月红,潘剑君,许信旺,杨丽霞.长白山森林土壤有机碳库大小及周转研究.水土保持学报,2006,20(6):99-102.
[11] 杨金艳,王传宽.东北东部森林生态系统土壤碳贮量和碳通量.生态学报,2005,25(11):2875-2882.
[13] 陆斌,王淑华,毛子军,孙涛,贾桂梅,靳世波,孙鹏飞,程春香.小兴安岭4种原始红松林群落类型生长季土壤呼吸特征.生态学报,2010,30(15):4065-4074.
[19] 夏汉平,余清发,张德强.鼎湖山3种不同林型下的土壤酸度和养分含量差异及其季节动态变化特性.生态学报,1997,17(6):645-653.
[20] 丁菡,胡海波,王人潮.半干旱区土壤酶活性与其理化及微生物的关系.南京林业大学学报:自然科学版,2007,31(2):13-18.
[21] 陈立明,满秀玲.云冷杉林土壤酶活性与土壤养分的研究.中国水土保持科学,2009,7(4):94-99.
[22] 王清奎,汪思龙,于小军,张剑,刘燕新.常绿阔叶林与杉木林的土壤碳矿化潜力及其对土壤活性有机碳的影响.生态学杂志,2007,26(12):1918-1923.
[23] 姜勇,张玉革,梁文举,闻大中.潮棕壤不同利用方式有机碳剖面分布及碳储量.中国农业科学,2005,38(3):544-550.
[27] 徐侠,陈月琴,汪家社,方燕鸿,权伟,阮宏华,徐自坤.武夷山不同海拔高度土壤活性有机碳变化.应用生态学报,2008,19(3):537-544.
[28] 姜培坤.不同林分下土壤活性有机碳库研究.林业科学,2005,41(1):10-13.
[33] 吴庆标,王效科,欧阳志云.活性有机碳含量在凋落物分解过程中的作用.生态环境,2006,15(6):1295-1299.
[34] 王清奎,汪思龙,于小军,张剑,刘燕新.杉木与阔叶树叶凋落物混合分解对土壤活性有机质的影响.应用生态学报,2007,18(6):1203-1207.
[35] 史学军,潘剑君,陈锦盈,杨志强,张黎明,孙波,李佩忠.不同类型凋落物对土壤有机碳矿化的影响.环境科学,2009,30(6):1832-1837.
[36] 李雪峰,韩士杰,郭忠玲,郑兴波,宋国正,李考学.红松阔叶林内凋落物表层与底层红松枝叶的分解动态.北京林业大学学报,2006,28(3):8-13.
[37] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡.植物生态学报,2000,24(5):518-522.
[38] 高志涛,吴晓春.蒙古栎地理分布规律的探讨.防护林科技,2005,(2):83-84.
[39] 陈大珂,周晓峰,赵惠勋,王义弘,金永岩.天然次生林四个类型的结构、功能及演替.东北林业大学学报,1982,(2):1-20.
[40] 何志斌,赵文智,刘鹄,苏永中.祁连山青海云杉林斑表层土壤有机碳特征及其影响因素.生态学报,2006,26(8):2572-2577.
[41] 渠开跃,冯慧敏,代力民,周莉.辽东山区不同林型土壤有机碳剖面分布特征及碳储量研究.土壤通报,2009,40(6):1316-1320.
[43] 耿玉清,余新晓,岳永杰,李金海,张国桢,刘松.北京山地针叶林与阔叶林土壤活性有机碳库的研究.北京林业大学学报,2009,31(5):19-24.
猜你喜欢
北京林业大学学报(社会科学版)(2022年4期)2023-01-04
建筑与预算(2022年6期)2022-07-08
小哥白尼(野生动物)(2021年9期)2022-01-17
纺织学报(2020年10期)2021-01-06
中国林副特产(2020年5期)2020-10-09
防护林科技(2017年7期)2017-09-01
绿色环保建材(2017年2期)2017-03-08
天津造纸(2016年1期)2017-01-15
前卫文学(2016年3期)2016-07-01
农家顾问(2016年5期)2016-05-14
