
环境因子对木棉种子萌发的影响
2013-09-11 08:37郑艳玲马焕成SchellerRobert
生态学报 2013年2期
郑艳玲,马焕成,* ,Scheller Robert,高 柱,郑 元
(1.西南林业大学国家林业局西南地区生物多样性保育重点实验室,昆明 650204;2.Department of Environmental Science and Management,Portland State University,Portland,Oregon,97207,USA)
木棉(Bombax malabaricum DC.;B.ceiba L.;Gossampinus malabarica Merr.)是木棉科(Bombacaceae)木棉属(Bombax Linn.)的一种落叶大乔木。该种分布于我国的江西、广东、广西、福建、台湾、四川、贵州、云南等多省;同时印度、斯里兰卡、印度尼西亚、菲律宾、澳大利亚均有分布。木棉全身是宝,其花、根、皮、木材和种子等都可以被利用,具有很高的经济、药用和观赏价值[1]。
干热河谷是我国西南地区的特殊生态系统类型,气候表现为热量充足,水分缺乏,干湿分明,气温全年较高,太阳辐射较强等特点[2]。由于生境条件的限制,干热河谷地区植被覆盖率低,水土流失严重,再加上人为的干扰破坏,干热河谷的生态环境不断恶化,因此,这一地区的生态环境恢复和建设势在必行[2]。在以干、热为显著特点的干热河谷,木棉是当地群落的优势种或常见种[3-4],反映其对当地气候具有很好的适应性。但是,河谷内,木棉成年树木生长良好,有大量结实,而其周围却很少有种苗出现。特殊生境的植物能较好地适应当地生境条件,其种子萌发行为在种群持续、种群动态和群落结构中起着关键作用[5],我们推测,一些环境因素影响木棉种子萌发。
对木棉的研究,国内外集中于木棉的传粉生物学、遗传多样性、组织培养、化学成分分析、纤维理化性质等方面[6-10]。但是木棉种子萌发方面的研究尚未见报道。因此,木棉种子萌发和幼苗生长的一些关键问题有待解决:种子萌发和幼苗生长的最适温度是多少?它们对光照、热激和干旱的敏感性如何?此外,大量研究表明,水杨酸(SA)可以诱导植物对多种胁迫产生一定抗性[11],那么SA处理对木棉种子萌发的抗旱性及抗热性又有何影响?本文通过对以上问题的研究,为实现木棉野外种群的建立和室内幼苗的高效生产提供科学依据,为应用该物种于干热河谷地区的生态恢复提供基础。
1 材料与方法
1.1 材料采集
木棉果实于2011年5月采集于云南省保山市隆阳区,将果实带回实验室自然风干,而后去除果皮和棉絮收集种子。
1.2 种子千粒重及含水量测定
随机抽取50粒种子称重,计算千粒重,设10个重复;种子含水量的测定采用烘干法,随机取25粒种子,在105℃烘箱中烘10 h后称重,计算种子水分含量,设5个重复。
1.3 种子吸胀实验
随机抽取25粒种子,称其干重,浸泡于25℃恒温水中,在前12 h内每隔2 h取出1次,用滤纸吸干表面水分后称重,而后每隔12 h取出1次,计算各自的吸水率,吸水率=(种子总重量-干种子重量)/干种子重量,设3个重复。
1.4 种子生活力测定
按照ISTA种子检验规程[12],采用四唑(TTC,1.0%)染色法测定木棉种子活力。将50粒种子置于35℃温水中浸泡2 d后去皮,然后在35℃条件下用四唑溶液染色12 h,染完色后根据种胚的着色程度和部位,按照国际种子检验规程上的标准鉴定种子的生活力。实验共设置4个重复。
1.5 环境因子对种子萌发和幼苗生长的影响
每个处理各选50粒种子均匀置于铺有两层滤纸、直径为10 cm的培养皿中于人工智能培养箱中进行萌发实验,每天补充一定量蒸馏水,以保持滤纸湿润。培养条件为光照时间12 h/d,光照强度2000 lx,除了检测温度和光照对种子萌发的影响外,其余实验都在35℃下进行。不同环境因子的每个处理水平均设4个重复。
种子萌发以胚根突破种皮2 mm为标准,逐日统计发芽数,并于实验的第10天称量幼苗鲜重,计算萌发率、萌发指数和活力指数。其中萌发指数GI=∑(Gt/Dt)(Gt为第t日的萌发数,Dt为相应的萌发日数);活力指数VI=GI×S(S为萌发第10日的幼苗鲜重)。
1.5.1 温度
培养温度分别恒定在10、15、20、25、30、35、40 和45 ℃8 个水平。
1.5.2 光照
25℃和35℃恒温条件下,在持续黑暗(在培养皿外包裹两层锡箔纸)和正常光照(12 h/d,2000 lx)下将种子进行萌发。
1.5.3 干旱处理
聚乙二醇(PEG)是一种高分子渗透剂,被广泛用来模拟干旱试验。采用PEG(6000)进行干旱胁迫处理,并检测水杨酸(SA)处理对抗旱性的影响。
种子在 PEG 中萌发:PEG(6000)水分胁迫溶液,质量浓度分别设 0(蒸馏水)、0.05、0.10、0.15、0.20 g/mL 5 个水平,与之相对应的溶液水势梯度大约为 0、-0.10、-0.20、-0.40、-0.6[13]。
SA处理后再干旱胁迫:室温下分别将种子在0、1、10、100 mg/L的SA溶液中浸泡24 h,然后在0.10 g/mL PEG中进行萌发。
1.5.4 热激处理
热激:种子在室温吸胀24 h后,分别于35、42、47和51℃热激2 h。
SA处理后再热激:室温下分别将种子在0、1、10、100 mg/L的SA溶液中浸泡24 h,然后在47℃热激2 h。
1.6 数据处理与分析
数据统计分析在Microsoft Excel 2003中进行,萌发率、萌发指数和活力指数在不同处理间的差异性分析采用单因子方差分析(One-Way ANOVA)。
2 结果与分析
2.1 种子千粒重、含水量和种子吸胀
实验表明,木棉种子的千粒重约为39.08 g,自然风干的种子含水量为10.62%。由图1可以看出,种子在25℃温水中浸泡24 h后含水量达到47.57%,而且观察发现,48 h后胚根就陆续突破种皮。可见,木棉种皮透水性良好,不影响种子对水分的有效吸收。

图1 木棉种子的吸水Fig.1 Water absorbing capacity of Bombax malabaricum seeds
2.2 种子生活力
四唑染色结果显示,木棉有生活力的种子占64.0%,糜烂粒占14.0%,其余22.0%未染色部分都是饱满种子,但失活原因还有待研究。
2.3 环境因子对种子萌发和幼苗生长的影响
2.3.1 温度对木棉种子萌发和幼苗生长的影响
表1为不同温度下木棉种子的萌发和幼苗生长情况。在15—35℃之间,随着温度升高,种子萌发速度加快(图2),萌发率依次提高,萌发指数和活力指数也依次显著增加(表1)。截止到萌发第10天,15℃和35℃时的萌发率分别达到7.00%和61.50%。40℃条件下少数种子虽然从第2天开始萌发,但在第4天时根尖开始变褐、变软,萌发的全部种子逐渐失活。在10℃和45℃条件下,没有种子萌发。观察发现,在萌发期间随着萌发温度升高,水分越多越不利,如35℃条件下,水分过多时已经萌发的种子会从根尖慢慢死亡,本实验中以保持滤纸湿润为宜。由此可见,高温会影响种子萌发和幼苗生长,而且高温高湿的影响作用更大。

表1 温度对木棉种子萌发率、萌发指数和活力指数的影响Table 1 Effect of temperature on germination percentage,germination index and vigor index of Bombax malabaricum
结果表示为平均值±标准差,同一行内字母不同代表在0.05水平上差异显著;
2.3.2 光照对木棉种子萌发和幼苗生长的影响
表2显示,25℃萌发时,光照对萌发率和活力指数无显著影响;35℃时,光照对萌发率、萌发指数和活力指数都无显著影响。经观察,两种温度下完全黑暗时产生的幼苗均为黄化苗。
2.3.3 干旱胁迫对种子萌发和幼苗生长的影响
(1)PEG胁迫对木棉种子萌发和幼苗生长的影响如表3所示,PEG浓度为0.05 g/mL时,种子萌发率虽与对照无显著差异,但萌发指数和活力指数都显著低于对照。随着胁迫程度的提高,种子的萌发率、萌发指数和活力指数都显著降低,PEG浓度为0.15 g/mL时,种子就不能萌发。表明木棉在种子萌发阶段对干旱比较敏感。

图2 不同温度条件下木棉种子萌发进程Fig.2 Seed germination percentage of Bombax malabaricum in different temperatures
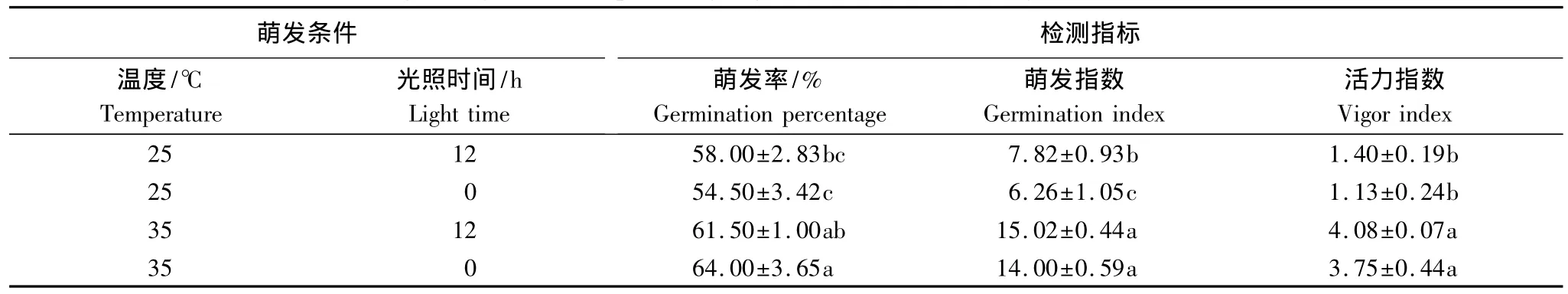
表2 光照对木棉种子萌发率、萌发指数和活力指数的影响Table 2 Effect of light on germination percentage,germination index and vigor index of Bombax malabaricum
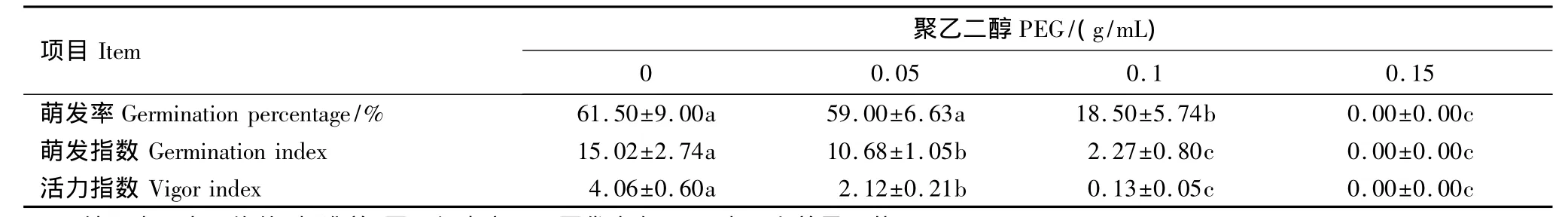
表3 不同浓度聚乙二醇(PEG)对木棉种子萌发率、萌发指数和活力指数的影响Table 3 Effect of Polyethylene glycol(PEG)on germination percentage,germination index and vigor index of Bombax malabaricum
(2)SA处理对种子萌发抗旱性的影响
由表4可以看出,1—100 mg/L SA处理后的萌发率、萌发指数和活力指数中,除10 mg/L SA处理后萌发指数显著高于对照外,其余指标均未比对照有所提高,甚至还显著低于对照。表明,SA处理并不能显著提高木棉种子萌发时的抗旱性,甚至对抗旱性有负面影响。

表4 水杨酸(SA)对木棉在0.1 g/mL PEG中萌发率、萌发指数和活力指数的影响Table 4 Effect of salicylic acid(SA)on germination percentage,germination index and vigor index of Bombax malabaricum in 0.1 g/mL PEG
2.3.4 热激对种子萌发的影响
(1)不同热激温度对种子萌发的影响
将室温下吸胀24 h的种子分别在35、42、47、51℃热激2 h后进行萌发,结果显示,随着温度升高,萌发率依次显著降低(图3)。
(2)SA处理对种子萌发抗热性的影响
表5表明,不同浓度的SA处理后,种子的萌发率、萌发指数和活力指数较对照都显著下降。因此,SA处理不利于提高种子萌发阶段的抗热性。
3 结论与讨论
种子重量是与植物的适应能力有关的一个很重要的性状[14-16],它被认为是在大量小种子和少量大种子之间的进化折中。在一定的能量限度内,种子重量和数量具有一种反向关系[17]。尤其在逆境或有高度竞争压力的环境中,大种子将会有更高的幼苗建植机会,因为它们能为幼苗的早期生长提供更多的储藏物质[18-19]。然而,在相同条件下,小种子有更大的传播、散布和拓殖优势,因此能够占据更多的生态位[20]。经测定,木棉种子比同属的长果木棉(Bombax insigne)及吉贝属的吉贝(Ceiba pentandra)种子都小,可以推测,木棉是通过种子数量和较高的种子散布能力在种间获得竞争优势的。除种间种子大小表型的进化外,由于物种内产生小种子同样有利于占据更多生态位,大种子有利于在竞争中占据优势,因此物种内也趋向于产生大小不等的种子[21]。据观察,不同居群间木棉种子的重量也存在一定差异,具体的遗传变异程度有待进一步研究。

图3 不同温度下热激2 h对木棉种子萌发率的影响Fig.3 Effect of heat shock for 2 h at different temperature on germination percentage of Bombax malabaricum图中直线代表±标准差
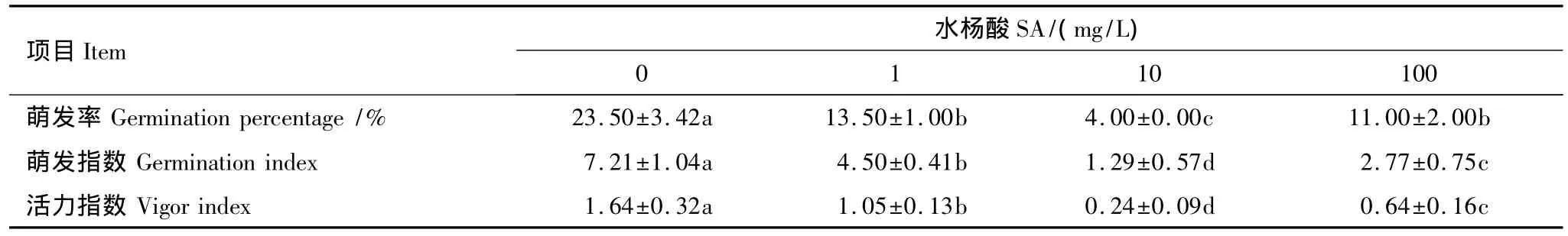
表5 SA处理对木棉种子在47℃热激2 h后萌发的影响Table 5 Effect of SA on germination percentage,germination index and vigor index of Bombax malabaricum after heat shock for 2 h at 47 ℃
干热河谷在热带大陆季风和西南暖湿气流双重影响下,季节性干旱的特征尤为突出。其冬半年为旱季,此期间各干热河谷的降水量一般在50—150 mm,占全年总降水量的8%—18%,且持续时间长达半年以上(11月—翌年5月);而雨季降水量占全年总降水量的82%—92%,降水集中,但持续时间较短(6—10月)[22]。由于蒸发量很高,不少地区不仅旱季干旱,雨季也出现严重的水分亏缺现象[23]。此外,干热河谷的年均气温一般较高,极端最高气温普遍达40℃以上。干热河谷气温高,地温则更高,尤其是在旱季后期,极端最高地面温度一般在70℃以上[24]。因此,“干”、“热”是干热河谷地区气候的显著特点。木棉能够生长在干热河谷这样严酷的环境中,能够正常开花结实,而在野外却发现大树周围很少有幼苗出现。在种子植物的生活史中,种子萌发阶段是植物对环境胁迫抵抗力最弱的时期,任何不利于种子萌发的因素存在都会直接影响到植物种群新个体的产生与补充,影响到种群的稳定性[25-26]。活力检测结果表明64%的木棉种子是有活力的,因此,幼苗的缺失必定是萌发的失败或萌发后幼苗的死亡所导致的,而这与环境因子密切相关。本文研究了木棉种子的基本物理特性,并且针对干热河谷干和热的特点,重点研究了种子萌发和幼苗生长对干旱和热激的响应。
研究结果表明,木棉种子吸水无障碍且光照对其种子萌发无影响,然而,种子萌发对高温和干旱却较敏感而且高温高湿对萌发后的种子也有致死效应。结合木棉种子成熟物候期和环境条件来看,木棉种子于5月左右开始成熟散落,而6—10月便是干热河谷的雨季和高温季节,此时,足够的水分及木棉种子快速吸水的特性保证了种子的充分吸胀。然而,研究结果显示种子在40℃时萌发率会显著下降,萌发后的种子也不能成活,因此,高温(包括地温和气温)是限制木棉种子在雨季萌发及萌发后幼苗存活的关键因子。此外,种子活力会因受到雨季高温高湿的影响而丧失或下降,从而导致种子在随后适宜环境下萌发的失败或幼苗活力的下降,而且到了旱季,干旱又成了限制种子萌发和幼苗生长的关键因子。这也就解释了为什么野外木棉树木结实正常而幼苗少的原因。
SA是一种酚类化合物,作为一种信号分子,在植物体内参与多种代谢调控,可以诱导植物对高温、低温、干旱、重金属、盐害等产生一定抗性[11]。作者对三棱栎(Trigonobalanus doichangensis)的研究中也发现,SA处理可以提高其种子萌发时的抗热性[27];然而本研究表明,不同浓度的SA处理并不能提高木棉种子萌发时的抗旱和抗热性,甚至对其有一定抑制作用,具体作用机理有待研究。有报道:外源SA的效应取决于多种因素,如植物物种和发育时期,施用方式和浓度以及植物内源SA水平[11]。
以上结果表明木棉种子能在较宽的温度范围内(15—35℃)萌发,而且温度越高,萌发越快,幼苗长势也越好。干热河谷地区,干旱和高温是限制木棉种子萌发的关键因子,而且SA处理并不能提高种子萌发时的抗旱和抗热性。
[1] South-Western Forestry College,Forestry Department of Yunnan Province.Iconographia Arbororum Yunnanicorum(Medius).Kunming:Yunnan Science and Technology Press,1990:230-231.
[2] Ou X K.Resource plants and there development ways in Jinsha River dry-hot valley.Journal of Yunnan University,1994,16(3):262-270.
[3] Jin Z Z.The floristic study on seed plants in the dry-hot valleys in Yunnan and Sichuan.Guihaia,1999,19(1):1-14.
[4] Jin Z Z,Yang Y P,Tao G D.The floristic characteristics,nature and origin of seed plants in the dry-hot river valley of SW China.Acta Botanica Yunnanica,1995,17(2):129-143.
[5] Bischoff A,Vonlanthen B,Steinger T,Müller-Schärer H.Seed provenance matters-Effects on germination of four plant species used for ecological restoration.Basic and Applied Ecology,2006,7(4):347-359.
[6] Bhattacharya A,Mandal S.Pollination biology in Bombax ceiba Linn..Current Science,2000,79(12):1706-1712.
[7] Wang S L,Li Q M.Population genetic diversity of Bombax malabaricum(Bombacaceae)in China.Acta Botanica Yunnanica,2007,29(5):529-536.
[8] Atta Alla H K,Moghazy E I,Waly A K.Micropropagation of Bombax malabaricum and Callistemon lanceolatus.Alexandria Journal of Agricultural Research,2003,48(1):103-114.
[9] Zhang X H,Zhu H L,Zhang S W,Yu Q,Xuan L J.Sesquiterpenoids from Bombax malabaricum.Journal of Natural Products,2007,70(9):1526-1528.
[10] Xiao H,Yu W D,Shi M W.Structures and performances of the kapok fiber.Journal of Textile Research,2005,26(4):4-6.
[11] Horváth E,Szalai G,Janda T.Induction of abiotic stress tolerance by salicylic acid signaling.Journal of Plant Growth Regulation,2007,26(3):290-300.
[12] ISTA.International rules for seed testing.Seed Science and Technology,1996,24(Suppl):151-154.
[13] Zhang F J,Li J Q,Xu X Y,Guo A Y,Hu J R,Du S X,Wan F H.Influence of environmental factors on seed germination of Flavera bidentis(L.).Kuntza.Acta Ecologica Sinica,2009,29(4):1947-1953.
[14] Hendrix S D,Nielsen E,Nielsen T,Schutt M.Are seedlings from small seeds always inferior to seedlings from large seeds?Effects of seed biomass on seedling growth in Pastinaca sativa L..New Phytologist,1991,119(2):299-305.
[15] Turnbull L A,Rees M,Crawley M J.Seed mass and the competition/colonization trade-off:a sowing experiment.Journal of Ecology,1999,87(5):899-912.
[16] Geritz S A H.Evolutionarily stable seed polymorphism and small-scale spatial variation in seedling density.The American Naturalist,1995,146(5):685-707.
[17] Yu S L,Chen H W,Li H.Review of advances in ecology of seed mass.Journal of Plant Ecology,2007,31(6):989-997.
[18] Moles A T,Westoby M.Seedling survival and seed size:a synthesis of the literature.Journal of Ecology,2004,92(3):372-383.
[19] Westoby M,Leishman M,Lord J,Poorter H,Schoen D J.Comparative ecology of seed size and dispersal[and discussion].Philosophical Transactions of the Royal Society B:Biological Sciences,1996,351(1345):1309-1318.
[20] Wu G L,Du G Z.Relationships between seed size and seedling growth strategy of herbaceous plant:a review.Chinese Journal of Applied Ecology,2008,19(1):191-197.
[21] Geritz S A H,van der Meijden E,Metz J A J.Evolutionary dynamics of seed size and seedling competitive ability.Theoretical Population Biology,1999,55(3):324-343.
[22] Li K,Liu F Y,Zhang C H.Essential characteristic of climate and plant recovery in Dry-hot valley.World Forestry Research,2009,22:24-27.
[23] Zhong X H.Degradation of ecosystem and ways of its rehabilitation and reconstruction in dry and hot valley.Resources and environment in the Yangtze Basin,2000,9(3):376-383.
[24] Qin J H.Studies on the Photosynthetic Ecophysiology and Adaptability to Stress Factors in the Dry Season in the Dry-Hot Valley,the Upper Reaches of the Yangtse River[D].Chengdu:Sichuan University,2006.
[25] Mills M H,Schwartz M W.Rare plants at the extremes of distribution:broadly and narrowly distributed rare species.Biodiversity and Conservation,2005,14(6):1401-1420.
[26] Jusaitis M,Polomka L,Sorensen B.Habitat specificity,seed germination and experimental translocation of the endangered herb Brachycome muelleri(Asteraceae).Biological Conservation,2004,116(2):251-266.
[27] Zheng Y L.Germplasm Conservation and Acclimatization Biology of the Endangered Trigonobalanus doichangensis(Fagaceae)[D].Kunming:Kunming Institute of Botany,Chinese Academy of Sciences,2010.
参考文献:
[1] 西南林学院,云南省林业厅.云南树木图志(中册).昆明:云南科技出版社,1990:230-231.
[2] 欧晓昆.金沙江干热河谷的资源植物及开发途径初探.云南大学学报:自然科学版,1994,16(3):262-270.
[3] 金振洲.滇川干热河谷种子植物区系成分研究.广西植物,1999,19(1):1-14.
[4] 金振洲,杨永平,陶国达.华西南干热河谷种子植物区系的特征、性质和起源.云南植物研究,1995,17(2):129-143.
[7] 汪书丽,李巧明.中国木棉居群的遗传多样性.云南植物研究,2007,29(5):529-536.
[10] 肖红,于伟东,施楣梧.木棉纤维的基本结构和性能.纺织学报,2005,26(4):4-6.
[13] 张风娟,李继泉,徐兴友,郭艾英,胡京蕊,杜淑欣,万方浩.环境因子对黄顶菊种子萌发的影响.生态学报,2009,29(4):1947-1953.
[17] 于顺利,陈宏伟,李晖.种子重量的生态学研究进展.植物生态学报,2007,31(6):989-997.
[20] 武高林,杜国祯.植物种子大小与幼苗生长策略研究进展.应用生态学报,2008,19(1):191-197.
[22] 李昆,刘方炎,张春华.干热河谷气候本质特征与植被恢复.世界林业研究,2009,22:24-27.
[23] 钟祥浩.干热河谷区生态系统退化及恢复与重建途径.长江流域资源与环境,2000,9(3):376-383.
[24] 秦纪洪.干热河谷几种常见植物光合生理生态及其环境的适应性研究[D].成都:四川大学,2006.
[27] 郑艳玲.濒危植物三棱栎的种质保存及驯化生物学研究[D].昆明:中科院昆明植物研究所,2010.
猜你喜欢
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
歌海(2021年6期)2021-02-01
幼儿园(2020年15期)2020-11-24
当代陕西(2020年9期)2020-08-04
艺术家(2020年5期)2020-07-09
Zoological Research(2019年3期)2019-06-28
海峡姐妹(2019年5期)2019-06-18
乡村地理(2018年4期)2018-03-23
中国工程咨询(2016年6期)2016-01-31
