
烯醇化酶ENO1抑制非小细胞肺癌细胞上皮间质转换
2013-09-09 08:51周鑫张莹韩廼珺郭素萍肖汀程书钧高燕宁张开泰
中国肺癌杂志 2013年5期
周鑫 张莹 韩廼珺 郭素萍 肖汀 程书钧 高燕宁 张开泰
Elizabeth Hay于1968年提出上皮间质转换的概念,并指出这种转换在一定条件下是可以逆转的[1]。事实上,EMT过程在后生动物的整个生命周期中都起到重要作用,如发育、组织损伤修复、纤维化,以及某些病理过程(如恶性肿瘤的转移)[2]。最新的统计[3]显示,肺癌死亡率仍高居各类癌症之首。非小细胞肺癌(non-small cell lung cancer, NSCLC)主要起源于上皮组织,因此大部分癌细胞维持上皮样特性,小部分细胞会通过EMT获得成纤维细胞样形态,失去胞间粘附并获得移动能力[4]。在分子水平上,这部分间质样细胞通常丢失E-钙粘蛋白(E-cadherin)、桥粒斑蛋白(Desmoplakin)和紧密连接蛋白(zonula occluden, ZO-1)等上皮样细胞表面分子,而获得间质样细胞标志如N-钙粘蛋白(N-cadherin)和波形蛋白(Vimentin),伴随着相关转录因子Twist、Snail和Slug的表达上调[5]。EMT是包含一系列精细复杂的变化,多种信号通路都参与其中,主要包括Wnt信号通路、TGFβ信号通路、Notch信号通路、Hedgehog信号通路、磷脂酰肌醇激酶(phosphatidyl inositol 3-kinase, PI3K)信号通路、核因转录因子(nuclear factor-κB, NF-κB)信号通路和ERK信号通路等;此外,还有多种microRNA参与这一过程的调控,如miR-155、miR-200和miR-205[6,7]。
除了肺癌,在诸如结肠癌[8]、胃癌[9]和乳腺癌[10]等多种实体瘤的研究中,都发现发生EMT的肿瘤细胞具有更高的恶性程度。并且,原发灶的肿瘤细胞出现EMT,常常预示着较差的临床预后。一项针对EMT相关转录因子Slug的临床研究[11]发现,肺癌组织中其mRNA水平越高,患者的术后复发几率越高,生存期越短。
相比于正常组织,肿瘤组织即使在有氧条件下糖酵解过程也明显加强,并且相关酶类活性和表达水平都有所提高,即Warburg效应[12]。随后更多研究发现,糖酵解过程相关酶类如己糖激酶(hexokinase-2, HK-2)、乳酸脱氢酶(lactate dehydrogenase-A, LDH-A)、甘油醛-3-磷酸脱氢酶(glyceraldehyde-phosphate dehydrogenase,GAPDH)和ENO1,实际上是具有包括催化活性在内的多功能蛋白[13]。编码ENO1的mRNA可以通过选择性翻译,表达另一种短异构体c-Myc启动子结合蛋白1(c-Myc promoter binding protein-1, MBP-1)[14]。多种证据表明在乳腺癌[15]、NSCLC[16]、丙肝病毒相关肝细胞肝癌[17]、前列腺癌[18]和神经胶质瘤[19]中,ENO1和MBP-1都参与了癌症的发展过程。在人乳腺癌细胞MCF-7中过表达MBP-1,可以抑制细胞的侵袭能力[15]。在胃癌中,ENO1和MBP-1可以通过抑制环氧合酶(cyclooxygenase-2, COX-2)表达抑制细胞EMT过程,从而抑制细胞侵袭和转移能力[20]。目前,ENO1对肺癌细胞EMT的影响,没有明确阐述。
本研究旨在揭示ENO1对NSCLC细胞系A549的EMT过程的影响,及其潜在的分子机制。
1 材料与方法
1.1 野生型和突变型ENO1基因的克隆和真核表达质粒的构建 以人胚胎cDNA文库作为模板,利用LATaq聚合酶(Takara,日本)克隆野生型ENO1全长;并构建至pcDNATM3.1/myc-His(-)A载体(Invitrogen,美国)中。随后,利用PCR引入点突变的方法,将野生型ENO1第94和97位氨基酸对应密码子ATG突变为CTG。突变型ENO1m同样构建于pcDNATM3.1/myc-His(-)A载体中。目的片段序列测定由上海生工生物工程公司完成。
1.2 细胞培养、脂质体介导的细胞转染 人NSCLC细胞系A549(ATCC,美国),在含有10%胎牛血清的RPMI-1640(Invitrogen,美国)完全培养基中置于5%CO2、37oC条件下培养。当细胞融合度达到90%时进行质粒转染,按照LipofectamineTM2000(Invitrogen,美国)产品说明书操作。转染质粒24 h后,替换含终浓度为600 μg/mL G418的完全培养基中进行筛选,并维持选择压力,从而获得稳定过表达ENO1的细胞亚群,命名为A549-ENO1。对照组转染空载质粒,命名为A549-Ctrl。
1.3 划痕实验 当细胞融合度达到100%时,在培养皿中均匀划出三道划痕。磷酸盐缓冲液洗涤去除划下的细胞。加入含2%胎牛血清的RPMI-1640培养基,于37oC、5%CO2条件下培养,此时记为第0天,并拍照记录。随后,每隔24 h更换含2%胎牛血清的培养基并拍照记录。
1.4 TGFβ-1诱导上皮间质转换实验 当细胞融合度在90%时,吸尽培养基,并用磷酸盐缓冲液洗涤。细胞经无血清培养饥饿24 h后,添加指定浓度TGFβ-1(R&D,美国)处理24 h,并拍照记录。
1.5 EGF刺激实验 当细胞融合度在50%-70%时,吸尽培养基,并用磷酸盐缓冲液洗涤。细胞经无血清培养基饥饿4 h后,加入终浓度为20 ng/mL的EGF(Cell Signal,美国)处理1 h。
1.6 细胞总蛋白的提取和Western blot分析 以适量含有1%苯甲基磺酰氟(PMSF)、1%蛋白酶抑制剂和1%磷酸酶抑制剂的RIPA蛋白裂解液(普利莱,中国)提取细胞总蛋白。采用BCA Protein Assay Kit(Thermo,美国)进行蛋白定量后用于Western blot分析。所用主要抗体及稀释比:ENO1(1:1,500,奥维亚,中国)、ERK1/2(1:2,000,Santa Cruz,美国)、p-ERK1/2(1:1,000,Santa Cruz,美国)、p-MEK1/2(1:1,000,Cell Signal,美国)、E-cadherin(1:1,000,Santa Cruz,美国)、N-cadherin(1:1,000,BD,美国)、Vimentin(1:500,Santa Cruz,美国)和β-actin(1:5,000,Santa Cruz,美国)。
1.7 统计学分析 细胞划痕实验结果图采用ImageJ2X(National Institutes of Health)采集数据,SPSS Statistics 17.0进行两组独立样本t检验, P<0.05为差异有统计学意义。
2 结果
2.1 筛选稳定过表达ENO1的A549细胞 收集筛选后的A549-ENO1和对照A549-Ctrl总蛋白,进行Western blot检测。结果显示,相比于对照组,A549-ENO1的ENO1表达水平更高(图1)。
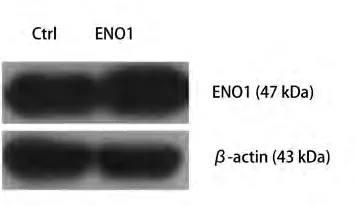
图 1 Western blot鉴定外源性ENO1基因在A549细胞中的过表达Fig 1 The over-expression of exogenous ENO1 was examined by Western blot
2.2 过表达ENO1抑制细胞运动 划痕24 h、48 h和72 h后,A549-ENO1划痕仅愈合了19.6%、38.4%和63.4%,均低于对照组A549-Ctrl的34.5%、60.1%和85.9%(P<0.05),表明ENO1过表达的A549细胞侧向运动能力明显下降(图2A)。
2.3 过表达ENO1抑制EMT过程 相比于对照组,A549-ENO1细胞中上皮样细胞标志物E-cadherin表达量明显上升,间质样细胞标志物N-cadherin和Vimentin的表达降低(图2B)。TGFβ-1诱导EMT实验结果显示,较高浓度的TGFβ-1(2 ng/mL)处理24 h才能使A549-ENO1群体中出现较多的成纤维样细胞;对照组A549-Ctrl在低浓度TGFβ-1(0.5 ng/mL)处理24 h后即出现较多的成纤维样细胞(图2C)。该结果说明ENO1过表达可以对TGFβ-1诱导A549细胞的EMT过程产生抑制效应。
2.4 ENO1抑制ERK磷酸化 相比于A549-Ctrl,A549-ENO1中磷酸化的ERK1/2水平明显下降(图3A)。细胞血清饥饿4 h后,用终浓度为20 ng/mL的EGF同时处理1 h后发现,A549-ENO1细胞中磷酸化的ERK1/2明显低于对照组,而MEK1/2的磷酸化几乎不受影响(图3B)。因此,我们推测ENO1可以通过抑制ERK1/2的磷酸化来抑制A549细胞的EMT。
2.5 全长ENO1可以抑制EMT 野生型ENO1wt在第94和97位氨基酸残基对应的密码子是ATG,定点突变型ENO1m在对应位置为CTG(图4A)。将野生型ENO1、突变型ENO1和空载质粒分别瞬时转染A549细胞48 h后,检测细胞中相关蛋白水平的变化,发现MBP-1表达没有明显的升高,并且突变型ENO1所产生的效应同野生型ENO1几乎完全相同(图4B)。这表明全长ENO1蛋白可以阻碍EMT过程。
3 讨论
本实验室利用新型蛋白质组研究体系建立了肺癌相关分泌/释放蛋白数据库,其中包含了ENO1在内的糖酵解途径关键酶[21];并且检测到在NSCLC患者和正常对照组外周血中,ENO1蛋白水平具有明显差异,因此,我们推测ENO1可能同NSCLC的发展具有密切联系[22]。本实验则针对ENO1在NSCLC细胞的EMT过程中的生物学功能进行了初步研究。
本研究在A549细胞系内稳定过表达ENO1,发现EMT相关分子标志物的改变和抑制ERK1/2磷酸化的现象,并导致A549细胞的运动能力下降。对比图2C中未添加TGFβ-1处理的细胞,我们发现过表达ENO1会引起A549细胞形态更趋近于上皮样细胞的变化,而对照组细胞更多地呈现出间质样细胞的梭形特征。根据这些现象,我们提出了ENO1通过抑制ERK1/2磷酸化来抑制EMT的假设。随后的TGFβ-1诱导EMT实验和EGF刺激ERK1/2活化的实验,均证实了这个假设。
在糖酵解过程中的很多酶都是多功能性的蛋白[13]。ENO1在不同的组织细胞中不同的生理和病理状态下,甚至是在不同的细胞亚定位均能够显示出不同的功能[23]。Hsu等[20]发现细胞核中的ENO1/MBP-1可以作为转录因子降低COX-2的转录水平,抑制胃癌细胞的EMT。对ENO1在NSCLC中抑制EMT的作用仍然没有解释清楚。
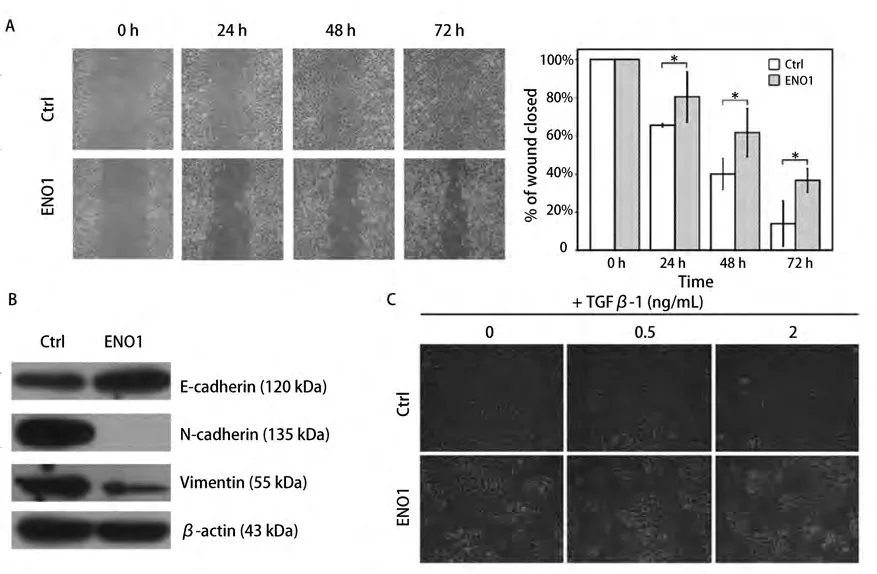
图 2 ENO1对A549细胞运动和EMT的抑制作用。A:划痕实验:在划痕后第24 h、48 h和72 h连续观察,过表达ENO1的A549细胞划痕恢复能力明显下降(*t检验,P<0.05)。图表纵坐标表示划痕宽度相对0 h时划痕宽度比,横坐标表示时间;B:Western blot结果显示,相对于A549-Ctrl,A549-ENO1细胞中,E-cadherin表达明显增加,N-cadherin和Vimentin表达量明显降低;C:梯度TGFβ-1诱导EMT实验:低剂量(0.5 ng/mL)处理A549-Ctrl细胞即可出现大量间质样细胞,而A549-ENO1细胞需要较高剂量(2 ng/mL)处理才能出现一定数量的间质样细胞。Fig 2 ENO1 inhibits A549 cell mobility and EMT process. A: Wound-healing assay: a continuous observation showed that over-expression of ENO1 resulted in limitation of A549 mobility (*t-test, P<0.05). In the chart, Y-axis represents the relative wound width and X-axis represents time period; B:Western blot assay: compared to A549-Ctrl, E-cadherin up-regulated in A549-ENO1, with down-regulation of N-cadherin and Vimentin; C: gradient dose of TGFβ-1 inducing EMT: quantity of mesenchymal-like cells were obtained in A549-Ctrl population after low dose (0.5 ng/mL) of TGFβ-1 inducing. Relative lower ratio of mesenchymal-like cells formed in A549-ENO1 population after high dose (2 ng/mL) of TGFβ-1 treatment.

图 3 ENO1抑制A549细胞ERK1/2的磷酸化。A:同对照组相比A549-ENO1细胞中ERK1/2磷酸化水平明显受限;B:20 ng/mL的EGF处理细胞,A549-ENO1的ERK1/2磷酸化水平仍然明显低于对照组细胞,而MEK1/2磷酸化活化几乎不受影响。Fig 3 ENO1 suppresses ERK1/2 phosphorylation in A549. Western blot result showed: A: Compared to A549-Ctrl group, ERK1/2 phosphorylation was inhibited by ENO1 over-expression; B: ERK1/2 phosphorylation level of A549-ENO1 was still lower than that of control group after EGF treatment, whereas MEK1/2 phosphorylation was almost not affected by ENO1 over-expression.
本研究结果显示全长ENO1蛋白本身具有抑制EMT的效应。为了排除MBP-1的干扰,我们构建了在ATG密码子处不产生选择性翻译MBP-1的突变型ENO1,并检测到过表达野生型ENO1和突变型ENO1在蛋白水平上对EMT相关分子标志物的改变作用几乎完全相同。而MBP-1对ERK信号通路的影响,则需要进一步的实验验证。

图 4 野生型和点突变ENO1具有相同作用。A:序列比对发现ENO1突变型存在两个ATG→CTG;B:Western blot检测显示出瞬时过表达野生型和突变型ENO1对E-cadherin、N-cadherin、p-ERK1/2变化影响是相似的。Fig 4 Wild type and point mutant ENO1 have the same effect.A: Sequence comparison: two point mutation ATG->CTG existed in mutant ENO1; B: Western blot assay: highly similar effect on E-cadherin, N-cadherin and p-ERK1/2 appeared after transient overexpression of wild type and mutant ENO1 in A549.
本研究不仅初步证实了ENO1的多功能性,为“糖酵解相关酶是多功能蛋白”的理论提供了新证据,而且也从影响细胞内信号传导的角度揭示ENO1抑制EMT的分子机理。经典的MAPK通路涉及到了Raf1→MEK1/2→ERK1/2的信号传递和放大过程[24],磷酸化的ERK1/2激活转录因子c-Jun和c-Fos表达,进而促进MMP-2等基因表达,降解胞外基质,促进细胞运动[25]。当细胞接受MEK1/2特异性抑制剂U0126处理后,ERK1/2信号通路被阻断,弱化TGFβ诱导EMT的效果,表明ERK1/2通路的活化是细胞EMT过程的一个必要条件[7]。ERK1/2的第204/187位酪氨酸残基和MEK1/2第218/222位丝氨酸残基的磷酸化,分别指示了这两种激酶的活化状态,并且MEK1/2是目前已知唯一能够活化ERK1/2的激酶[26,27]。基于这些条件,本实验分析了ENO1的过表达对ERK1/2信号通路的影响,进而发现了一个有趣的现象,即ENO1过表达引起活性状态的ERK1/2降低,而不影响MEK1/2活化。在另外一个研究[18]中,研究人员发现前列腺癌细胞中MBP-1可以通过同MEK5的直接相互作用抑制MEK5/BMK1的非经典MAPK信号通路。因此,我们推测在ERK1/2磷酸化/去磷酸化调节过程中,ENO1可能通过某种机制抑制了MEK1/2的催化活性,但不会影响MEK1/2的活化;也有可能是ENO1过表达,增强了PTPs和MKPs等磷酸酶活性,但这些推论需要更多的实验证据来支持。
猜你喜欢
波谱学杂志(2022年1期)2022-03-15
皮肤病与性病(2021年3期)2021-07-30
中国产前诊断杂志(电子版)(2020年4期)2021-01-11
中国医学物理学杂志(2020年11期)2020-12-12
东北大学学报(自然科学版)(2020年5期)2020-05-22
天津医科大学学报(2019年6期)2019-08-13
满族文学(2018年6期)2018-12-27
学生天地·小学低年级版(2017年12期)2018-04-16
分析化学(2017年12期)2017-12-25
安徽医科大学学报(2015年9期)2015-12-16
