
小寨子沟自然保护区领春木种群数量特征与空间分布格局*
2013-07-12 07:30姬慧娟陈思多马文宝何建社刘兴良
四川林业科技 2013年6期
姬慧娟,陈思多,马文宝,何建社,张 利,刘兴良
(1.四川省林业科学研究院,四川 成都 610081;2.阿坝州科学技术研究院,四川 汶川 623000)
1 前言
植物种群数量特征是植物个体生存能力与外界环境相互作用的结果[1],其种群年龄结构和生命表以及存活曲线不仅反映种群生存现状,也能反映种群的数量动态及变化趋势,并在很大程度上体现植物与环境间的抗争关系,尤其对于濒危植物的保护和利用研究具有重要意义[2]。种群空间分布格局反映了种群个体在水平空间上彼此间的相互关系,是由种群本身生物学特性和环境条件的综合影响所决定的。种群空间分布格局的分析是研究种群生物学特性、种内和种间关系以及种群与环境关系的重要手段,一直是植物生态学中的研究热点[3],对空间格局的分析可以反映种群过去所经历的一系列生态学过程[4],是研究种群特征、种内和种间关系以及种群与环境关系的重要手段[5]。
领春木为领春木科领春木属(Euptelea)落叶灌木或小乔木,古老的残遗植物,为典型的东亚植物区系成分的特征种,被列为国家Ⅲ级重点保护植物[6]。已有研究主要涉及其系统位置的确定[7~9],领春木的种子休眠与萌发特性[10],而对该种的种群数量动态与空间分布格局研究报道很少。在北川小寨子沟自然保护区领春木林主要分布在海拔1 800 m~2 200 m 的局部地区,林冠不齐,乔木层主要以领春木为优势种,通过在不同生境的领春木种群中开展种群数量特征与空间分布格局研究,能够反映领春木种群在不同生境中的扩展方式、繁殖能力,及其对不同生境的适应能力,研究结果将为领春木种群的合理保护和资源保存提供科学依据。
2 研究地概况
小寨子沟自然保护区地处四川盆地西缘,属于横断山脉东缘的龙门山系中段,介于东经103°45'~104°26'与北纬31°50'~32°13'之间。该地区属典型的亚热带季风气候,据保护区管理处所处位置蔡家坪(海拔1 690 m)的观测,年均温9.2℃,年均降水量约800 mm,年日照1 527.5 h,平均相对湿度80%。由于小寨子沟自然保护区山体高大,气候垂直分布带明显,形成了多种多样的小气候环境[11],因此植被垂直带谱明显,植被垂直带谱为:常绿落叶阔叶混交林(1 300 m~2 000 m)、落叶阔叶林(1 000 m~2 300 m)、亚高山针叶林(2 500 m~3 400 m)、亚高山灌丛(3 400 m~3 600 m)、亚高山草甸(3 400 m~3 600 m)、高山灌丛(3 600 m~3 800 m)、高山草甸(3 800 m~4 200 m)、高山流石滩稀疏植被(4 200 m 以上),部分植被类型在分布海拔范围上有重叠现象[12]。
3 研究方法和数据处理
3.1 取样方法
由于在北川小寨子沟自然保护区领春木主要分布在海拔1 600 m~2 200 m 的河岸带中,沿河岸在领春木典型分布的区域设置20 m×30 m 的样方进行植物群落学调查,共调查5个样方。记录以下指标:各样方的海拔、经纬度、坡度、坡向、植物总盖度;胸径大于0.2 cm 的乔木的树高、胸径、冠幅,胸径小于0.2 cm 的乔木的高度和株数;灌木和草本的种类。表1是对调查区5个样方生境状况的统计。对领春木的统计以基株为单位进行,一是由种子萌发的实生苗长成的植株,二是由根系或树桩萌蘖生成的植株,如果一个树桩上有多个无性系分株,则以其中胸径最大的分株记录。同时以每个20 m×30 m的样方的两个垂直边为坐标轴,记录每株领春木的坐标(x,y),用于种群空间格局分析。
3.2 数据处理
3.2.1 大小级结构图的绘制
由于领春木为濒危植物,生长锥钻取年轮对其种群破坏较大,因此,本研究用胸径大小作为度量领春木年龄大小级的指标。根据调查得到的数据,将DBH <0.2 cm 的领春木幼苗作为大小级结构的第Ⅰ级;将胸径0.2 cm≤DBH <2.5 cm 的领春木作为第Ⅱ级;其后,以DBH=5 cm 作为步长增加一级,即2.5 cm≤DBH <7.5 cm 作为第Ⅲ级,以此类推,将小寨子沟自然保护区中的领春木种群划分为7个大小级。以大小级作为纵坐标,各大小级中个体百分比作为横坐标,绘制总体和每个样地中领春木种群的大小级结构柱状图。
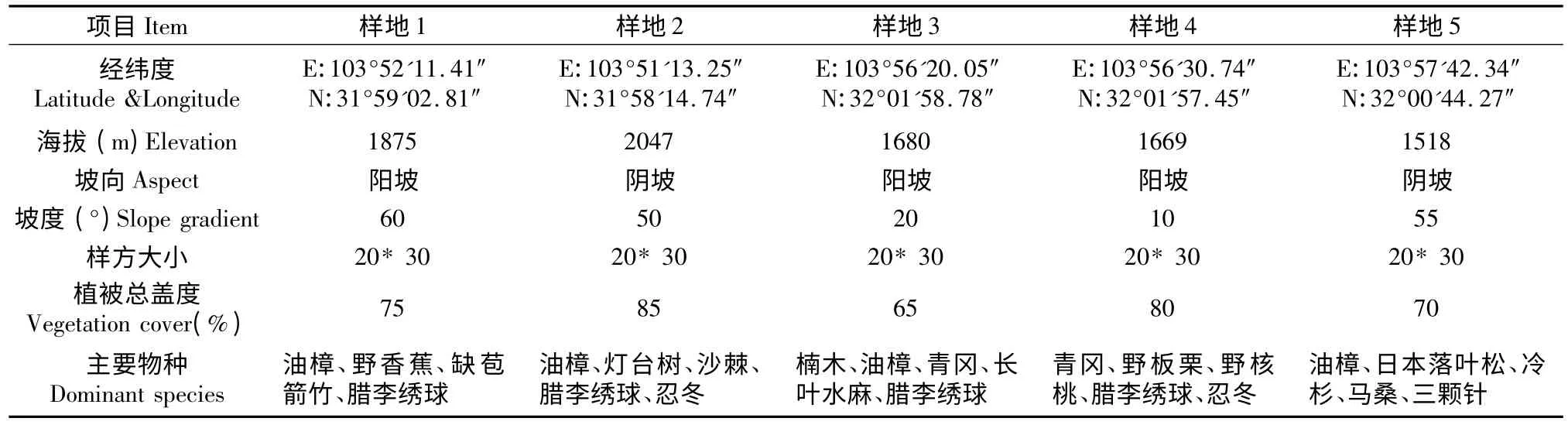
表1 小寨子沟自然保护区领春木生境状况Table 1 Habitat condition of E.pleiospermum in Xiaozhaizigou Nature Reserve
3.2.2 静态生命表的编制和存活曲线的绘制
统计各大小级内的领春木株数,按照静态生命表的编制方法,分别对小寨子自然保护区中领春木总体和5个样地的领春木种群编制静态生命表。静态生命表一般包括如下参数[5,13]:
x 为大小级,在静态生命表中代替年龄等级;
Nx为在x 大小级内出现的领春木个体数;
Lx为存活数标准化,Lx=(Nx+Nx+1)/2;
lgLx为Lx取以10 为底的对数;
Dx为从x 到x +1 大小级的死亡数,Dx=Nx-Nx+1;
Qx为从x 到x+1 的死亡率,Qx=Dx/Nx;
T 为从x 大小级起超过x 大小级的存活个体总数,T=∑Lx;
Ex为生命期望,表示第x 大小级的个体在未来所能存活的平均年数,Ex=Tx/Nx。
以生命表的大小级为横坐标、lgLx为纵坐标,绘制总体和每个样地的领春木种群的存活曲线。
3.2.3 种群空间分布格局
本研究以样方为基础采用相邻格子样方进行种群空间格局分析。根据记录领春木的坐标(x,y),在坐标平面上把各样方划分成5m×5m 的小样方,记录各个小样方内领春木的个体数,用于空间分布格局的拟合。格局分析指标包括以下3个[5,13]:

此式可用于离散分布理论的拟合,统计学基础是波松分布。若c=l,则个体分布符合波松分布,是随机分布;若c >l,则个体分布趋向于集群分布;若c<l,则趋向于均匀分布。该值的显著性可以用t 检验:
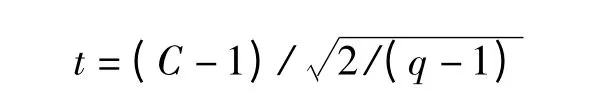
比较t 与t0.05(q-1)确定其差异显著性。
平均拥挤度系数:
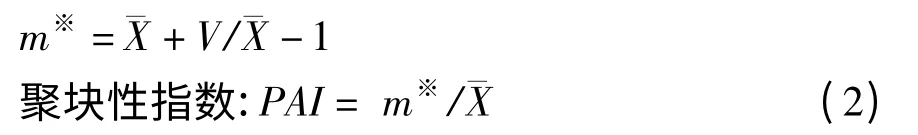
平均拥挤度是指每个个体在一个样方中的平均其它个体数,即每个个体的平均邻居数。聚块性指数可用于聚集程度的度量,以客观反映格局强度,由于它考虑了空间格局本身的性质,并不涉及密度,其值越大,聚集性越强。PAI=l,则个体分布为随机分布;PAI >l,则为集群分布;PAI <1,则为均匀分布。
Morisita 指数:
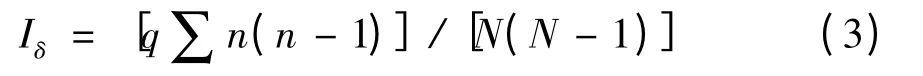
Iδ=l,则为随机分布;Iδ>l,则为聚集分布;Iδ<1,则个体分布为均匀分布。该方法的检验可用F 检验:
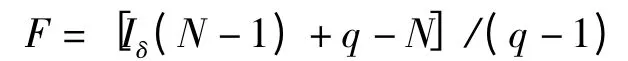
显著性水准值可以从F 表中查得,分子自由度为q-1,分母自由度为∞。
以上公式中,V 为样本方差,为样方中观测值的平均值,q 为小样方数,n 为样方中的观测值,N 为样方内个体总数。
4 结果与分析
4.1 领春木种群的大小级结构
从图1a~f 中可以看出,各样方的种群大小级结构与整个北川小寨子沟自然保护区种群特征基本一致。由图1a可以看出,整个自然保护区中,领春木种群胸径在2.5 cm 以下的幼龄树(Ⅰ和Ⅱ级)较少,个体数为50株,占领春木样本数的21.83%;胸径为2.5 cm~12.5 cm 的中龄树(Ⅲ和Ⅳ级)最多,个体数为158株,占领春木样本数的68.99%;Ⅴ级以上的老龄树最少,个体数仅为21株,占总样本数的9.18%。由图1a~f 还可看出,Ⅰ级幼苗所占的比例非常低,依次为5.24%、6.56%、1.75%、6.67%、2.56%,在样地5 中甚至没有Ⅰ级幼苗,表明领春木幼苗严重缺乏。而且,纵观整个自然保护区领春木种群的大小级结构,均呈中龄树比例最大、幼龄树和老龄树比例次之的纺锤型结构。以上结果表明,北川小寨子沟自然保护区的领春木种群属于衰退型种群。
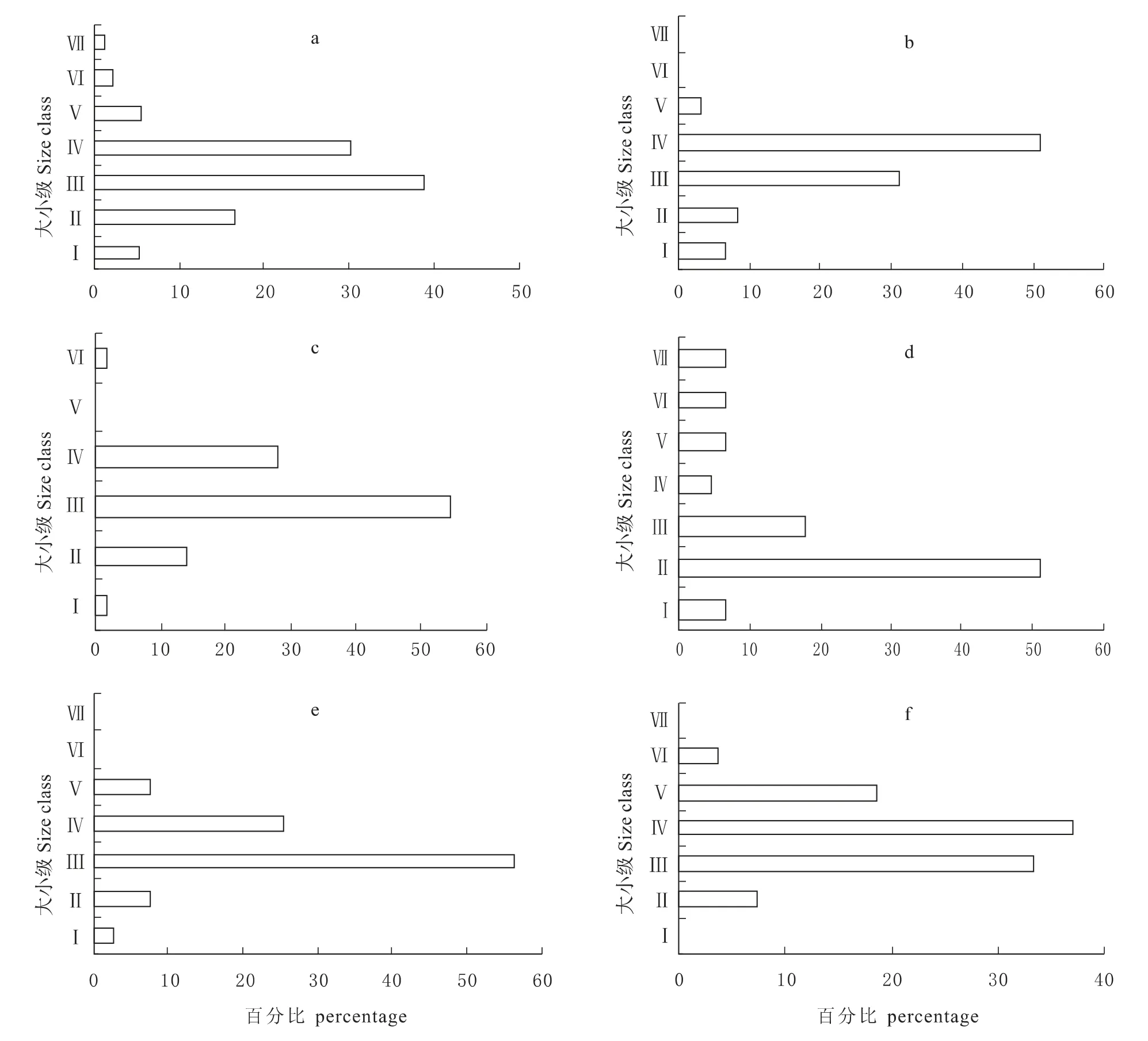
图1 领春木种群的大小级结构Fig.1 The size structure of E.pleiospermum population
4.2 领春木种群的静态生命表和存活曲线
4.2.1 领春木种群的静态生命表
按照静态生命表的编制方法,根据5个样方中调查所得的数据编制了领春木种群的静态生命表(表2)。从表2 样方合计中可以看出,第Ⅰ、Ⅱ级个体数均小于其后面一级,导致出现死亡率为负的现象;在第Ⅳ级时死亡率出现一个高峰,在第Ⅶ级达到最大值,这与存活数一栏第Ⅶ级领春木数量极少相吻合。
表2 中样方1 与样方5 对比可以看出,5个样方中领春木种群生命表特征基本能反映领春木种群在整个自然保护区中的典型特征,但在死亡率、生命期望等数量动态参数上有差异。样方3 的死亡率在第Ⅳ、Ⅵ两级均有相同的最大值,这是因为第Ⅴ级在该样方中不存在所致,这与表2 样方合计中在最后一个大小级死亡率达到最大是不相矛盾的。

表2 北川小寨子沟自然保护区领春木种群静态生命表Table 2 Static life table of E.pleiospermum in Xiaozhaizigou Nature Reserve
4.2.2 领春木种群的存活曲线
北川小寨子沟自然保护区中领春木种群的存活曲线(图2)。从该图可以看出,5个样方中领春木的存活曲线基本上均呈Deevey Ⅰ型,以这5个样方调查所得的数据为基础汇总后绘制而成的整个北川小寨子沟自然保护区领春木种群的存活曲线亦接近DeeveyⅠ型。胸径小于0.2 cm (第Ⅰ龄级)的领春木存活率较低,幼树(第Ⅱ龄级)的领春木存活率就有所升高,中龄级种群存活率达到最高,然后随着龄级的增加,存活率开始下降,至老龄期存活率最低,死亡率达到最大。
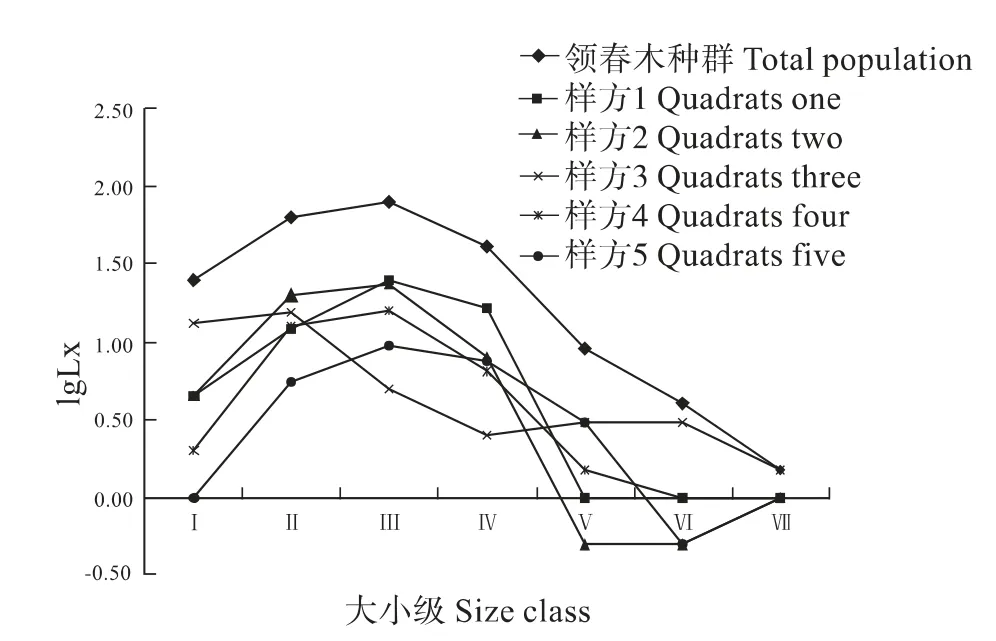
图2 领春木种群的存活曲线Fig.2 Survivorship curves of E.pleiospermum population
4.3 领春木种群的空间分布格局
从表3可以看出,在北川小寨子沟自然保护区所取的5个样方均呈现集群分布。但是聚集程度有所差异,特别是样方3,由于其特殊的地域环境,使其呈现明显的集群分布,在调查过程中发现,在该样方中某些区域幼树大量聚集,构成了集群性很高的集群分布。
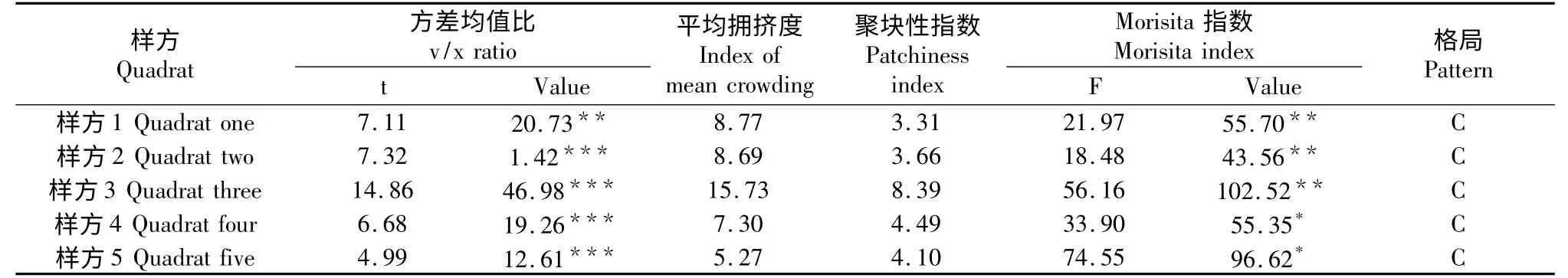
表3 北川小寨子沟自然保护区各样方中领春木种群的空间分布格局Table3 Spatial distribution patterns of E.pleiospermum population in different quadrats
5 讨论
北川小寨子沟自然保护区中领春木种群幼龄期个体缺乏,中龄期个体相对丰富,老龄期个体数量稀少,纺锤型的大小级结构表明种群属于衰退型(图1)。林冠层郁闭度较高,缺乏光照,且沿河岸带分布,物种丰富,种间竞争激烈,导致幼苗死亡率较高[14];另外,领春木的果实具翅,其中含有抑制物,使得带翅的种子萌发率仅为30%左右(未发表资料)是导致领春木种群幼龄期个体缺乏的主要原因。领春木种群幼苗的补充对群落的发展起着决定性的作用[13],如果群落中不出现重大干扰或者林窗,领春木的幼苗很难出现,没有幼苗的补充,随着演替的进行,领春木的个体不断死亡,数量逐渐减少,最终势必会被更新能力更强的物种替代[15]。与神农架地区河岸带领春木相比,北川领春木的萌蘖率比较低(35%),老龄期个体数量稀少,则是由于领春木为小乔木或灌木,其平均高度仅有5.5 m,达不到亚热带森林的林冠层,吸收不到充足的阳光,使其生理寿命减少(胸径很少超过22.5 cm)。
领春木种群静态生命表显示出领春木种群在第1、II 级时具有较高的生命期望,这表明领春木种群在北川小寨子沟自然保护区中将维持较长时间。Ⅳ级出现死亡率高峰,是由于随着个体的增大,对空间、光照和营养的竞争激烈,出现自疏现象造成的。这说明领春木种群在生长过程中,当胸径达到10 cm(即第Ⅳ级)左右时,种内和种间竞争增强,种群发展遇到一个瓶颈,而随后种群将趋于稳定发展,按照其生物学特性,随着大小级的增加死亡率逐渐升高。
然而对于北川小寨子沟自然保护区中的领春木种群来说,虽然其幼苗的存活率较低,但由于其幼树的存活率较高,其存活曲线接近Deevey I 型。在I级的领春木幼苗H <33 cm,缺乏光照使其生长受限。进入II、III 级后,领春木个体生长迅速,平均树高增加,更接近灌木层,竞争力增强,光照资源相对增大,存活率逐渐增高。随后,由于个体的长大,对光照的需求量大于灌木层所能获取的光照.对空间和矿质营养的需求量也逐渐增大,导致种群之间和种群内部为争夺光照、空间和矿质营养而发生激烈竞争,种内竞争产生自疏现象,种间竞争也导致部分领春木个体死亡,从而使存活率开始下降。到第Ⅶ级,个体接近生理寿命。死亡率达到最高峰。这与杨慧等对北京东灵山地区优势种之一的白桦和神农架地区河岸带领春木种群的研究结果一致[13,16]。
北川小寨子沟自然保护区的领春木种群空间分布格局总体为聚集分布。这与在更大尺度(100 m×100 m)上对领春木的研究结果一致[17],也与多数珍稀植物种群的空间分布格局一致[18]。人为干扰和自然环境通过影响其种群个体数量和密度而改变种群的分布格局。据我们观察,北川小寨子沟自然保护区的领春木种群分布范围收缩到河岸附近,表现为沿河流的“一带多岛”现象。贺善安和郝日明研究发现,鹅掌楸种群的地理分布格局因被多次分隔、隔离,在整体上也呈现“一带五岛”的分布格局[18]。领春木种群集中沿河岸分布,这是物种分布范围收缩的表现,是种群衰退的标志。
鉴于以上原因,不仅应加强对现有领春木种群的保护,而且更需要采取一定的措施促进其种群的延续与发展。减少领春木种子萌发和个体发育过程中受到高大乔木的遮荫和浓密的缺苞箭竹林的抑制,应对群落进行适度的干扰,开辟适当的林窗,扩大其种子扩散范围,促进领春木种子萌发,幼苗、幼树正常发育。
[1]Crawly MJ.Plant Ecology[M].London:Blackwell Scientific Publications,1986.
[2]Molles MC Jr.Ecology,Concept and Applications 2nd edn[M].New York,McGraw-Hill Companies,2002:186~437.
[3]Wiegand T.Moloney K A.Rings,circles.and null-models for point pattern analysis in ecology[J].Oikos,2004,104:209~229.
[4]Johnson EA,Fryer GI.Population dynamics in lodgepole pine-Engelmann spruce forest[J].Ecology,1989,70(5):1335~1345.
[5]张金屯.数量生态学[M].北京:科学出版社,2004:243~254.
[6]傅立国,金鉴明.中国植物红皮书:稀有濒危植物[M].第一版.北京:科学出版社,1992:680~681.
[7]李红芳,任毅.领春木茎次生木质部中导管穿孔板的变异[J].植物分类学报,2005,43(1):1~11.
[8]田先华,李红芳,庞承义,等.领春木(领春木科)导管穿孔板中纹孔膜残余的观察[J].西北植物学报,2005,25(7)1345~1349.
[9]潘开玉,路安民,温洁.领春木的染色体数目及配子体的发育[J].植物分类学报,1991,29:439~444.
[10]周佑勋.领春木种子休眠与萌发特性[J].中南林业科技大学学报,2009,29:51~54.
[11]赵颖,何兴金,秦自生,等.小寨子沟自然保护区种子植物区系分析[J].江 西科学,2004,22(5):32~36.
[12]罗辅燕.小寨子沟自然保护区的植被分类[J].内江师范学院学报,2005,20(4):72~76.
[13]魏新增,黄汉东,江明喜,等.神农架地区河岸带中领春木种群数量特征与空间分布格局[J].植物生态学报,2004,32(4):825~837.
[14]国家林业局国有林场和林木种苗工作总站.中国木本植物种子[M].北京:中国林业出版社,2001:175-176.
[15]蔡晟,刘学全,张家来,等.鄂西三峡库区大老岭珍稀树木群落特征研究[J].应用生态学报,2000,(11):165~168.
[16]杨慧,娄安加,高益军,等.北京东灵山地区白桦种群生活史特征与空间分布格局[J].植物生态学报,2007,31(2):272~282.
[17]江明喜,党海山,黄汉东,等.三峡库区香溪河流域河岸带种子植物区系研究[J].长江流域资源与环境,2004,18(2):368~374.
[18]张文辉,祖元刚,刘国彬.十种濒危植物的种群生态学特征及其致危原因分析[J].生态学报,2002,22(9):1512~1520.
[19]贺善安,郝日明.中国鹅掌楸自然种群动态及其致危生境的研究[J].植物生态学报,1999,23(1):87~95.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业调查规划(2020年3期)2020-06-03
运动(2017年1期)2017-09-03
湖北农业科学(2017年11期)2017-07-13
剑南文学(2016年15期)2016-08-22
广西农学报(2015年4期)2015-09-11
中国水利(2015年4期)2015-02-28
四川党的建设(2015年4期)2015-02-05
