
Programmed death 1 and programmed death ligand 1 expressions in patients with chronic hepatitis B
2013-07-03 09:20
Hangzhou, China
Programmed death 1 and programmed death ligand 1 expressions in patients with chronic hepatitis B
Wen-Jin Zhang, Chuan-Hui Peng and Shu-Sen Zheng
Hangzhou, China
BACKGROUND:The role of programmed death 1 (PD-1) and programmed death ligand 1 (PD-L1) in persistent HBV infection is controversial. Increasing PD-1 and PD-L1 expression has been found in hepatitis B patients during immune clearance phase, but not in HBV-tolerant patients. We investigated PD-1 and PD-L1 expression and inflammation in chronic hepatitis B.METHODS:Twenty patients with chronic hepatitis B participated in this study. Fifteen patients were in the immune clearance phase, and 5 were in the immune inactive phase. Circulating HBV-specific T cells were analyzed by flow cytometric detection of major histocompatibility complex (MHC) class I peptide complexes, known as pentamers. Intra-hepatic PD-1 and PD-L1 expressions were analyzed by immunostaining.
RESULTS:The frequency of pentamers, including core 18-27 (1.88%±0.36%), env 335-343 (1.85%±0.37%), and pol 575-583 (1.56%±0.29%) was 8.30-, 7.71- and 8.48-fold greater during immune clearance phase than those during the immune inactive phase. In addition, more than 70% of circulating pentamers were PD-1 positive. During immune clearance phase, the numbers of intra-hepatic PD-1 and PD-L1 positive cells were 108±23/HPF and 97±20/HPF respectively, in contrast, there was a paucity of PD-1 and PD-L1 positive cells in the immune inactive phase. The numbers of intra-hepatic PD-1 and PD-L1 positive cells were positively correlated with serum alanine aminotransferase and the number of intra-hepatic CD8+T cells. Immunofluorescence showed that almost all of the intrahepatic CD8+T cells were PD-1 and CCR6 positive. These cells aggregated around macrophage inflammatory protein-3 alpha (MIP3α) positive cells and mixed with PD-L1 positive cells.
CONCLUSIONS:PD-1 and PD-L1 expressions were significantly correlated with inflammation. CCR6 and PD-1 co-expressed in the same cells; these cells were increased both in circulation and the inflamed liver and aggregated around MIP3αpositive cells. The mixture of CCR6 and PD-1, MIP3αand PD-L1 positive cells created immune response compartments which played an important role in specific immune response in HBV immune clearance.
(Hepatobiliary Pancreat Dis Int 2013;12:394-399)
programmed death 1 receptor; hepatitis B, chronic; liver
Introduction
The HBV-specific immune response plays a critical role in determining the outcomes of HBV infection.[1-3]Persistent HBV infection is always attributed to malfunction of HBV-specific immune responses including T-cell proliferation, cytokine production and cytotoxicity.[1-3]Up-regulation of programmed death 1 (PD-1) and programmed death 1 ligand (PD-L1/B7H1) dampens the HBV-specific immune response. Blocking the PD-1/PD-L1 interactionin vitrocan reverse the exhausted cytokine production and the proliferation of HBV-specific T cells.[2-5]
A recent longitudinal analysis of chronic hepatitis B patients revealed that the levels of PD-1 and PD-L1 expression were increased significantly during the inflammatory flare period and decreased gradually when inflammation regressed.[6,7]Zhang et al[6]also reported that PD-1 signals were increased during inflammation to offset the positive co-stimulatory signal. In acute hepatitis B patients, delayed up-regulation of PD-1 and PD-L1 were attributed to severe liver inflammation and finally led to acute liver failure.[7]In vitroanalysis revealed that co-stimulatory and co-inhibitory molecules were up-regulated synchronously after onsetof stimulation.[8]These results suggested that during the immune response, increasing co-inhibitory signals offset co-stimulatory signals to avert severe tissue damage by an over-vigorous immune response.
PD-L1 expression was detected not only in inflamed tissue, but also at low levels in normal nonlymphoid tissues such as the liver, lung, heart and kidney.[9-11]Eppihimer et al[12]suggested the vascular endothelium is the source of PD-L1 mRNA in peripheral tissues. Thus PD-1/PD-L1 interaction in normal tissues may prevent tissue inflammation and contribute to homeostatic maintenance.
In this study, we analyzed PD-1 and PD-L1 expression in chronic hepatitis B patients. Along with PD-1 and PD-L1 detection, features of the immune response, such as HBV-specific T cells clone expansion, chemotaxis, and antigen engulfment and presentation, were measured by flow cytometry and immunostaining.
Methods
Subjects
Twenty chronic hepatitis B patients were enrolled in this study from August 2008 to August 2011; all were human leukocyte antigen A2 (HLA-A2) positive. All of these patients were negative for anti-HCV, anti-HDV, anti-HIV-1, and anti-HIV-2 antibodies. Fifteen patients were in the immune clearance phase, with HBV-DNA load>5×103copies/mL and serum ALT levels nearly 5 times the upper limit (50 U/L) of normality. For comparison, 5 patients with normal serum ALT and high HBV-DNA levels >5×103copies/mL, were considered immune inactive subjects. Serum HBV-DNA was quantified using a sensitive real-time polymerase chain reaction technique. Liver biopsy was performed for all patients. Written informed consent was obtained from each patient, and the study protocol was approved by the Ethics Committee of our hospital.
Pentamers and PD-1 analysis
The following antibodies were used for peripheral blood mononuclear cell (PBMC) staining: anti-CD8-FITC, anti-PD-1-APC, and anti-HLA-A2 (BioLegend, San Diego, CA, USA). HBV-specific CD8+T cells were identified with HLA-A2 pentamers containing core 18-27 peptide (FLPSDFFPSV), env 335-343 peptide (FLLTRILTI), and pol 575-583 peptide (FLLSLGIHL), all labeled with fluorochrome PE. PBMCs from peripheral blood were incubated for 15 minutes at 37 ℃ with 1 µg of PE-labeled pentamers complex in PBS and then washed twice. The cells were then incubated with saturating concentrations of directly conjugated anti-CD8-FITC and PD-1-APC mAb for 15 minutes. After washing twice, the cells were analyzed immediately on a FACSCalibur flow cytometer using CellQuest software (BD Biosciences, East Rutherford, NJ, USA).
Lymphocytes were gated according to their physical parameters. Pentamer positive responses were reported as the percentage of pentamer positive T cells in the total CD8+population. Frequencies of pentamer positive cells exceeding 0.08% of CD8+cells were considered positive responses. The expansion (n-fold) was calculated as the ratio of the pentamer positive T cells (percent) between the experimental and background levels.
Immunohistochemical staining
Acetone-fixed liver tissue cryosections (5 µm) were incubated with anti-PD-1, anti-PD-L1, and anti-CD8 (Abcam plc, Cambridge, UK) antibodies overnight at 4 ℃ after blocking endogenous peroxidase activity with 0.3% H2O2. Positive cells (brown color) were counted in two different high-power fields (HPFs) by two independent observers. Immunofluorescence staining was also performed. In brief, liver tissues were incubated overnight with monoclonal primary antibodies followed by secondary antibodies for 45 minutes at room temperature. Anti-PD-1, anti-PD-L1 (both diluted 1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-CD8, anti-CCR6, anti-MIP3α (all diluted 1:50; Dako, Glostrup, Denmark) and HBsAg antibody were used as primary antibodies. Fluorescein isothiocyanate-conjugated immunoglobulin G and fluorochrome PE-conjugated immunoglobulin G (Santa Cruz Biotechnology) were used as secondary antibodies.
Statistical analysis
Data were analyzed by SPSS 13.0 software. Quantitative data are presented as mean±SD. Comparisons between groups were made using Student'sttest. APvalue of <0.05 was considered statistically significant.
Results
Proliferation of HBV-specific T cells in peripheral circulation
The frequency of pentamers, multiple HBV-specific CD8+T cells, was assessed in chronic hepatitis B patients. In immune clearance episodes, HBV-specific CD8+T cells, including core 18-27 (1.88%±0.36%), env 335-343 (1.85%±0.37%), and pol 575-583 (1.56%±0.29%), were increased significantly (20-fold greater than background, which never exceeded 0.08%, Fig. 1A). In comparison,patients during the immune inactive phase, the frequencies of core 18-27 (0.23%±0.027%), env 335-343 (0.24%±0.041%), and pol 575-583 (0.18%±0.036%) were significantly fewer than those in the immune clearance phase (Fig. 1B). The frequency of core 18-27, env 335-343, and pol 575-583 were 8.30, 7.71 and 8.48 times greater in the immune clearance phase than those in the immune inactive phase (Fig. 1C).
PD-1 expression on total circulating CD8+T and pentamer cells
The levels of PD-1 expression on total circulating CD8+T cells and HBV-specific CD8+T cells were detected in chronic hepatitis B patients. PD-1 expression on total CD8+T cells was increased more significantly in chronic hepatitis B patients in the immune clearance phase than that in the immune inactive phase. The PD-1 frequency on total circulating CD8+T cells was 31.2%±6.1% in the immune clearance phase, whereas it was 11.7%±4.1% in the immune inactive phase. The frequency of PD-1 expression was significantly higher on pentamer positive cells (core 18-27, env 335-343, and pol 575-583 were 72.3%±12.9%, 71.3%±12.4%, and 70.5%± 10.0%, respectively) than that on total circulating CD8+T cells (31.2%±6.1%) (Fig. 2).
Analysis of intra-hepatic CD8, PD-1, and PD-L1 expression
Serially sectioned liver biopsies from patients with chronic hepatitis B were examined for CD8, PD-1 and PD-L1 expression by immunohistochemistry. During the immune clearance phase, the mean number of intrahepatic CD8+cells was 60±14 per HPF, but only a few CD8+cells were observed in the liver in the immune inactive phase. In inflamed liver tissue, CD8+cells infiltrated the lobular regions and aggregation of CD8+cells were observed in fibrous septa. Intra-hepatic PD-1 and PD-L1 expression was also detected throughout inflamed liver tissue. The mean numbers of PD-1 and PD-L1 positive cells were 108±23/HPF and 97±20/HPF, respectively. In contrast, few PD-1 and PD-L1 positive cells were observed in liver tissue from patients in the immune inactive phase (Fig. 3).
Intra-hepatic PD-1 and PD-L1 expression and immune response
Intra-hepatic PD-1 and PD-L1 expressions were detected by immunofluorescence in association with features of the immune response including chemokine production, PD-1 positive cell migration, distribution and aggregation, and the engulfment and presentationof HBV antigen. Almost all of the intra-hepatic CD8+cells were PD-1 positive (Fig. 4). In addition, all of these intra-hepatic PD-1 positive cells expressed the homing receptor CCR6 and aggregated around the cells expressing macrophage inflammatory protein-3 alpha (MIP3α), ligand of CCR6. PD-L1 positive cells, which co-expressed HBsAg, mixed with PD-1 positive cells in inflamed liver (Fig. 5).
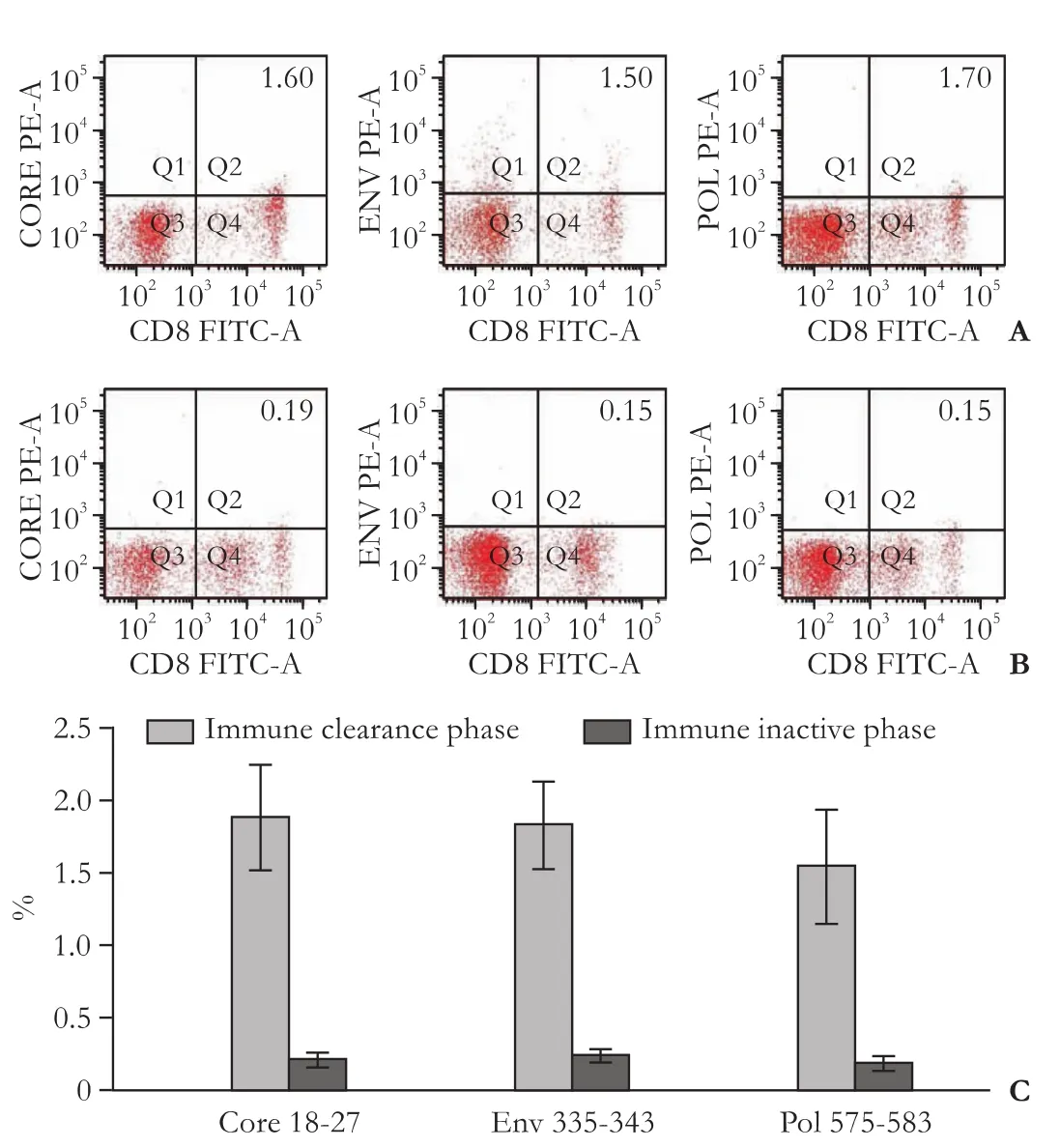
Fig. 1.Representative dot plots of pentamer expression on circulating CD8+T cells in chronic hepatitis B patients during immune clearance (A) and immune inactive phase (B);C: The comparison of circulating pentamer frequency between patients in immune clearance phase and those in the immune inactive phase.

Fig. 2.Representative dot plots of PD-1 expression on total circulating CD8+T cells in chronic hepatitis B patients during the immune inactive phase (A) and those in the immune clearance phase (B);C: Representative dot plots of PD-1 expression on circulating pentamer positive cells in chronic hepatitis B patients during the immune clearance phase;D: The comparison of PD-1 expression on total circulating CD8+T cells between patients in the immune clearance phase and those in the immune inactive phase (*:P<0.05);E: The comparison of PD-1 frequency between pentamers and total circulating CD8+T cells during the immune clearance phase (#:P<0.01, vs CD8).
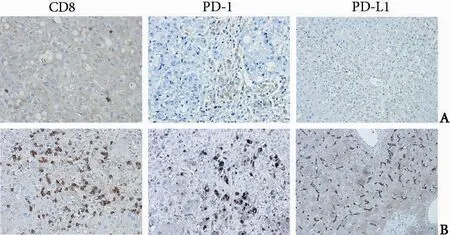
Fig. 3.Immunohistochemical staining of intra-hepatic CD8, PD-1, and PD-L1 positive cells in patients with chronic hepatitis B (original magnification ×400).A: Representative figures from patients in the immune inactive phase;B: Representative figures from patients in the immune clearance phase.

Fig. 4.Co-localization of PD-1 with CD8, CCR6, MIP3α, and PD-L1 as shown by immunofluorescence double staining in liver biopsy specimens from patients with chronic hepatitis B during the immune clearance phase (original magnification ×200).A: PD-1 (red) co-localized with CD8+(green) cells. The 2-color merged panels show co-localization (yellow);B-D: PD-1 (green) co-localized with CCR6, MIP3α, and PD-L1 (red) positive cells. The 2-color merged panels show co-localization (yellow).
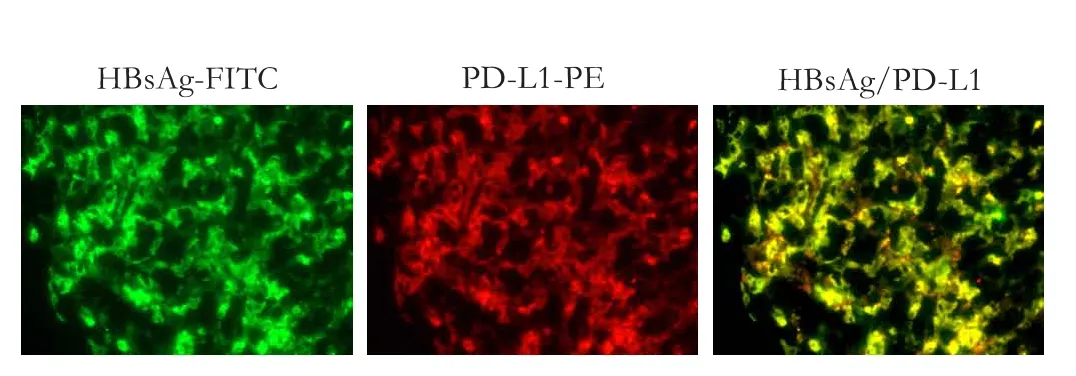
Fig. 5.Co-localization of PD-L1 with HBsAg as shown by immunofluorescence double staining in liver biopsy specimens from patients with chronic hepatitis B during the immune clearance phase (original magnification ×200). HBsAg (green) co-localized with PD-L1 (red) positive cells. The 2-color merged panels show co-localization (yellow).
Discussion
It is well recognized that activation of naive T cells requires T-cell receptor (TCR) and co-stimulatory signals. When naive T cells differentiate into memory cells, their activation processes change. Memory T cells can be activated by co-stimulatory signal in the absence of the TCR signal.[9,13,14]As memory T cells can be easily activated, their co-inhibitory molecule system should play key roles in chronic infection.
PD-1, a member of the CD28 family, is a negative regulator of the immune system. Expression of PD-1 is induced on lymphocytes upon activationin vitro, and predominantly expressed on activated T cellsin vivo.[9,15]PD-L1, a ligand for PD-1, is constitutively expressed in nonlymphoid tissues such as the heart, lung, placenta, kidney and liver and is always upregulated during the period of immune response.[9-11]Cell surface PD-L1 expression is generally induced on APCs upon activation. Intra-hepatic PD-L1 expression is significantly up-regulated on Kupffers, liver sinusoidal endothelial cells (LSECs), dendritic cells and vascular endothelial cells during the period of inflammation.[9]The expression of PD-1 and PD-L1 on activated T cells and APCs suggested that the interaction of this pair of co-inhibitory molecules at the effector phase of immune responses may limit the degree of immune response.[6,7,9]
Our results showed that in the setting of immune clearance phase of chronic hepatitis B, pentamers, multiple HBV-specific CD8+T cells including core 18-27, env 335-343, and pol 575-583, proliferated more than 20 times higher than background levels. More than 70% of circulating pentamers were PD-1 positive. Immunostaining also showed that in the inflamed livertissue, PD-1 positive cells expressing homing receptor CCR6, expressed predominantly on memory T cells,[16-18]aggregated around the MIP3α positive cells. MIP3α, the ligand of CCR6, is always produced by the phagocytic cells such as Kupffers and dendritic cells and this production is significantly increased by inflammation mediators such as TNF-α in inflamed liver.[18]Increasing MIP3α production in inflamed liver attracted CCR6-expressing effector memory T cells to the site of immune response to fulfill their cytotoxic effect. These data suggested that CCR6-expressing effector T cells are systematically concentrated from circulating to the inflamed liver to form functional compartments via the MIP3α-CCR6 interactions. CCR6-expressing effector T cells could be directed to the appropriate site to execute cytotoxic effects. This should minimize the severity of the immune response. A previous study[19]revealed that the great majority of intra-hepatic CD8+T cells in chronic hepatitis B are not anti-HBV specific. Our data revealed that more than 70% of pentamers were CCR6 positive and the PD-1 positive T cells were not detected in the spleen and lymph node (data not shown) in chronic hepatitis B during the immune clearance phase. Based on these data, we conclude that PD-1-expressing HBV-specific CD8+T cells were also recruited to the inflamed liver via MIP3α-CCR6 interaction. These results were consistent with the previous studies in which circulating activated T cells were attracted to the liver and were trapped in the inflamed liver to perform their function.[20-23]
In our study, the numbers of intra-hepatic PD-1 and PD-L1 positive cells were positively correlated with the degree of hepatitis (data not shown). Intrahepatic PD-L1 expression on non-parenchymal cells such as Kupffers, LSECs and vascular endothelial cells, was increased significantly during immune clearance episodes. When effector/memory T cells are attracted to the inflamed liver by MIP3α, they first contact with Kupffers, LSECs and vascular endothelial cells. The interaction between PD-1 on effector memory T cells and PD-L1 on non-parenchymal cells can limit the degree of the immune response at the sites of the inflamed liver by increasing negative regulatory signal. In this study, immunofluorescence showed that in the milieu of the inflamed liver, PD-1 positive cells always mixed with PD-L1 positive cells. Almost all of the PD-L1 positive cells were HBsAg-positive. Thus, PD-L1 expressing non-parenchymal cells engulfed HBsAg and presented it to the PD-1 positive effector/memory T cells. Based on these data, we concluded that the level of HBV-specific immune response was always limited by the interaction between PD-1 and PD-L1.
The positive correlation existed between the intrahepatic PD-1/PD-L1 expression and the levels of serum ALT and number of intra-hepatic CD8+cells (data not shown). So we concluded that during the immune clearance phase, the higher level of co-inhibitory signals limit the level of immune response. Thus the interaction between PD-1 and PD-L1 in the inflamed liver of chronic hepatitis B patients may set upper threshold for HBV-specific immune response. If HBV was eliminated successfully under this upper threshold, the patients would recover; otherwise, HBV will remain and replicate to sufficient levels to trigger the next immune response episode.
PD-1 immunotherapy was used recently in clinical trials. In a stage I clinical trial anti-PD-1 monoclonal antibody therapy demonstrated a lack of significant side effects.[7,9,24-26]If anti-PD-1 monoclonal antibody is used, the upper threshold of immune response in patients with chronic hepatitis B may be moderately elevated, and then the optimal HBV-specific immune response may help the clearance of HBV.
Contributors:ZWJ and ZSS proposed the study. ZWJ performed research and wrote the first draft. PCH collected and analyzed the data. All authors contributed to the design and interpretation of the study and to further drafts. ZSS is the guarantor.
Funding:This study was supported by a grant from the Zhejiang Provincial Natural Science Foundation (Y2110169).
Ethical approval:The study protocol was approved by the Ethics Committee of our hospital.
Competing interest:No benefits in any form have been received or will be received from a commercial party related directly or indirectly to the subject of this article.
1 Das A, Hoare M, Davies N, Lopes AR, Dunn C, Kennedy PT, et al. Functional skewing of the global CD8 T cell population in chronic hepatitis B virus infection. J Exp Med 2008;205: 2111-2124.
2 Peng G, Li S, Wu W, Tan X, Chen Y, Chen Z. PD-1 upregulation is associated with HBV-specific T cell dysfunction in chronic hepatitis B patients. Mol Immunol 2008;45:963-970.
3 Evans A, Riva A, Cooksley H, Phillips S, Puranik S, Nathwani A, et al. Programmed death 1 expression during antiviral treatment of chronic hepatitis B: Impact of hepatitis B e-antigen seroconversion. Hepatology 2008;48:759-769.
4 Chen L, Zhang Z, Chen W, Zhang Z, Li Y, Shi M, et al. B7-H1 up-regulation on myeloid dendritic cells significantly suppresses T cell immune function in patients with chronic hepatitis B. J Immunol 2007;178:6634-6641.
5 Boni C, Fisicaro P, Valdatta C, Amadei B, Di Vincenzo P, Giuberti T, et al. Characterization of hepatitis B virus (HBV)-specific T-cell dysfunction in chronic HBV infection. J Virol 2007;81:4215-4225.
6 Zhang Z, Zhang JY, Wherry EJ, Jin B, Xu B, Zou ZS, et al.Dynamic programmed death 1 expression by virus-specific CD8 T cells correlates with the outcome of acute hepatitis B. Gastroenterology 2008;134:1938-1949.
7 Kassel R, Cruise MW, Iezzoni JC, Taylor NA, Pruett TL, Hahn YS. Chronically inflamed livers up-regulate expression of inhibitory B7 family members. Hepatology 2009;50:1625-1637.
8 Zhang WJ, Xie HY, Duan X, Wan YL, Peng CH, Shi SH, et al. Study of human B7 homolog 1 expression in patients with hepatitis B virus infection. World J Gastroenterol 2012;18: 3681-3695.
9 Iwai Y, Terawaki S, Ikegawa M, Okazaki T, Honjo T. PD-1 inhibits antiviral immunity at the effector phase in the liver. J Exp Med 2003;198:39-50.
10 Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med 2000;192:1027-1034.
11 Dong H, Zhu G, Tamada K, Chen L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat Med 1999;5:1365-1369.
12 Eppihimer MJ, Gunn J, Freeman GJ, Greenfield EA, Chernova T, Erickson J, et al. Expression and regulation of the PD-L1 immunoinhibitory molecule on microvascular endothelial cells. Microcirculation 2002;9:133-145.
13 Müllbacher A, Flynn K. Aspects of cytotoxic T cell memory. Immunol Rev 1996;150:113-127.
14 Flynn K, Müllbacher A. The generation of memory antigenspecific cytotoxic T cell responses by CD28/CD80 interactions in the absence of antigen. Eur J Immunol 1997;27:456-462.
15 Karrar A, Broomé U, Uzunel M, Qureshi AR, Sumitran-Holgersson S. Human liver sinusoidal endothelial cells induce apoptosis in activated T cells: a role in tolerance induction. Gut 2007;56:243-252.
16 Baba M, Imai T, Nishimura M, Kakizaki M, Takagi S, Hieshima K, et al. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J Biol Chem 1997;272:14893-14898.
17 Liao F, Rabin RL, Smith CS, Sharma G, Nutman TB, Farber JM. CC-chemokine receptor 6 is expressed on diverse memory subsets of T cells and determines responsiveness to macrophage inflammatory protein 3 alpha. J Immunol 1999;162:186-194.
18 Dieu-Nosjean MC, Vicari A, Lebecque S, Caux C. Regulation of dendritic cell trafficking: a process that involves the participation of selective chemokines. J Leukoc Biol 1999;66: 252-262.
19 Maini MK, Boni C, Lee CK, Larrubia JR, Reignat S, Ogg GS, et al. The role of virus-specific CD8(+) cells in liver damage and viral control during persistent hepatitis B virus infection. J Exp Med 2000;191:1269-1280.
20 Wuensch SA, Pierce RH, Crispe IN. Local intrahepatic CD8+T cell activation by a non-self-antigen results in full functional differentiation. J Immunol 2006;177:1689-1697.
21 Mehal WZ, Juedes AE, Crispe IN. Selective retention of activated CD8+T cells by the normal liver. J Immunol 1999;163:3202-3210.
22 John B, Crispe IN. TLR-4 regulates CD8+T cell trapping in the liver. J Immunol 2005;175:1643-1650.
23 John B, Crispe IN. Passive and active mechanisms trap activated CD8+T cells in the liver. J Immunol 2004;172:5222-5229.
24 Armah GE, Breiman RF, Tapia MD, Dallas MJ, Neuzil KM, Binka FN, et al. Immunogenicity of the pentavalent rotavirus vaccine in African infants. Vaccine 2012;30:A86-93.
25 Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer 2012;12:252-264.
26 Kline J, Gajewski TF. Clinical development of mAbs to block the PD1 pathway as an immunotherapy for cancer. Curr Opin Investig Drugs 2010;11:1354-1359.
January 10, 2013
Accepted after revision May 31, 2013
Author Affiliations: Division of Hepatobiliary Pancreatic Surgery, First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou 310003, China (Zhang WJ, Peng CH and Zheng SS)
Shu-Sen Zheng, MD, PhD, FACS, Division of Hepatobiliary Pancreatic Surgery, First Affiliated Hospital, Zhejiang University School of Medicine, Hangzhou 310003, China (Tel: 86-571-87236570; Email: shusenzheng@zju.edu.cn)
© 2013, Hepatobiliary Pancreat Dis Int. All rights reserved.
10.1016/S1499-3872(13)60061-2
 Hepatobiliary & Pancreatic Diseases International2013年4期
Hepatobiliary & Pancreatic Diseases International2013年4期
- Hepatobiliary & Pancreatic Diseases International的其它文章
- Effect of L-cysteine on remote organ injury in rats with severe acute pancreatitis induced by bile-pancreatic duct obstruction
- Impact of periampullary diverticula on the outcome and fluoroscopy time in endoscopic retrograde cholangiopancreatography
- The diagnostic value of high-frequency ultrasonography in biliary atresia
- Endobiliary radiofrequency ablation for malignant biliary obstruction
- Hepatic abscess associated with Salmonella serotype B in a chronic alcoholic patient
- Biliary-colonic fistula caused by cholecystectomy bile duct injury