
CO2摩尔分数升高对兴安鹿蹄草光合生理特性的影响1)
2013-06-28 09:12:10崔大练马玉心
东北林业大学学报 2013年1期
崔大练 马玉心
(浙江海洋学院,舟山,316000)
蔡体久
(东北林业大学)
自19世纪70年代工业革命以来,由于人类活动的影响,大气CO2摩尔分数正逐步上升。目前,已由 100 a 前的 260 ~280 μmol·mol-1上升到 350 μmol·mol-1左右,并继续以每年 1 ~2 μmol·mol-1的速度增加[1],据估算,到2050年大气CO2摩尔分数将升高到 450 ~550 μmol·mol-1。21 世纪末将达到 700 μmol·mol-1[2]。随着 CO2及其他温室气体摩尔分数升高,大气温度及降水也将发生变化。CO2摩尔分数的升高不但会引起全球变暖和气候变化,而且对植物特别是森林植物有着直接的影响,将直接影响植物的光合作用过程和生长发育[3-4]。因此,对植物在高CO2摩尔分数下的光合响应特性研究便成为当前国际上一个较为重要和热点的研究课题。植物叶片光合速率在高CO2摩尔分数下可以提高40% ,甚至61%,目前的研究多数集中在栽培植物,特别是对生长在控制环境中的小苗和幼树进行研究[5-7]。对北方针叶林下的草本植物,特别是常绿草本植物在高摩尔分数CO2影响下的光合响应特性的研究更缺乏。北方针叶林作为森林生态系统的一个重要主体,对于全球碳平衡起着重要的调节作用。由于温度的升高,枯枝落叶层的腐烂速度加速,反馈性的使森林内的CO2摩尔分数增加得更快,而草本植物与地被层最接近,对CO2摩尔分数的升高在光合特性方面反应最直接、最敏感。
兴安鹿蹄草(Pyrola dahurica(H.Andr.)Kom.)为鹿蹄草科常绿草本植物,主要分布于我国东北地区,是寒温带针阔混交林的指示植物[8]169-171。兴安鹿蹄草的叶片在冬季仍然存活,冬季被雪覆盖,在高寒地区形成了独特的适应寒冷的机制[9]。其叶片革质,四季常绿,是冬季鹿、狍等动物的主要食物[8]169-170,也是重要药物[10]。因此,野外自然生长条件下,对东北针阔混交林下兴安鹿蹄草叶片光合作用在高CO2摩尔分数下的短期响应特性进行研究,不仅有助于揭示CO2摩尔分数的改变对我国寒温带针阔混交林植物的影响机理,为研究温室效应对寒温带森林的影响提供参考数据,也可为我国寒温带森林产量生态学研究提供基础资料。
1 研究地概况
试验于2006—2007年在黑龙江省尚志县帽儿山地区中国国家森林观测实验站进行。地理位置:东经 127°30'~127°34',北纬 45°20'~45°25'。该地区自然概况为年平均气温2.8℃,≥10年积温为2 495.74 ℃ ,最冷月(1 月份)平均气温-19.7 ℃,最热月(7月份)平均气温21℃(表1)。无霜期为120~140 d。年降水量724 mm,年蒸发量1094 mm。土壤为暗棕壤。地带性植被为红松阔叶混交林,主要有红松(Pinus koraiensis Sieb.et Zucc.)、胡桃楸(Juglans mandshurica Maxim.)、水曲柳(Fraxinus mandshurica Rupr.)、黄波椤(Phellodendron amurense Pupr.)、白桦(Betula platyphylla Suk.)及山杨(Populus davidiana Dode)等[11]。

表1 2006年4—9月份兴安鹿蹄草样地的气温变化 ℃
2 材料与方法
试验材料为兴安鹿蹄草。分布于中国东北、朝鲜北部、俄罗斯远东地区。多年生常绿草本植物。其新叶淡绿色,老叶暗绿色。花蕾形成于3—4月份,果实9月份成熟。
试验于2006—2007年4—9月份在帽儿山生态实验站进行,逐月(每月15日)测定兴安鹿蹄草老叶与新叶光合特性。利用LI-6400便携式光合测定系统,配备红、篮人工光源及CO2钢瓶测量不同CO2摩尔分数下植物的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间 CO2摩尔分数(Ci),CO2摩尔分数的梯度设置为:0、50、200、350、700、1 000、1300、1600 μmol·mol-1,叶室 CO2摩尔分数通过 LI-6400外带CO2气源(液化CO2小钢瓶),由系统内的自动程序控制CO2吸收器(Scrubber)来完成,。有效光辐射强度(PA,R)控制在 800 μmol·m-2·s-1。测定最少稳定时间为120 s,当测定结果变化率小于0.1时,光合测定系统自动记录数据。每个CO2摩尔分数下测量3个植株的3个正常叶片。数据导入计算机后,用Excel程序统计分析,绘制图表。
3 结果与分析
3.1 净光合速率(Pn)对CO2摩尔分数升高的响应
如表2所示,兴安鹿蹄草老叶4、5、6月份的净光合速率(Pn)都随着CO2摩尔分数的升高而逐渐升高。从表2中可以看出,5月份的净光合速率明显高于4、6月份,而且当 CO2摩尔分数超过700 μmol·mol-1时,5月份与6月份的净光合速率差异极显著。在5月份,当CO2摩尔分数由350 μmol·mol-1增加到 700 μmol·mol-1时,Pn有一个急速上升过程,之后变化逐渐缓和;CO2摩尔分数从350 μmol·mol-1增至 1 600 μmol·mol-1时,5 月份的Pn增幅达154.2%。在6月份,当CO2摩尔分数由200 μmol·mol-1增加到 350 μmol·mol-1时,Pn有一个急速上升过程,CO2摩尔分数从200 μmol·mol-1增至 1 600 μmol· mol-1时,6 月份的 Pn增幅达97.13%。4月份 CO2摩尔分数从50 μmol·mol-1增至 1 600 μmol·mol-1过程中,Pn几乎呈直线上升,增幅为385.1%。从表2中可以看出,CO2的饱和点为 1 300 μmol·mol-1。
4、5、6 月份的 CO2补偿点分别为 45.3、48.4、61.2 mol·mol-1。CO2补偿点是区分 C3和 C4植物的重要参数,C3植物 CO2补偿点较高,常大于30 mol·mol-1;C4植物很低,总是在 10 mol·mol-1以下。从兴安鹿蹄草的CO2补偿点来看(并结合解剖学特征,后续发表),该植物应为C3植物。C3植物的CO2补偿点受一些环境因素的影响[12-13]。在一定的温度范围内,C3补偿点随温度升高而升高[14-16]。
如表2所示,兴安鹿蹄草新叶的净光合速率于6—9月份随CO2摩尔分数的升高而呈上升趋势。当 CO2摩尔分数从 50 μmol·mol-1到 700 μmol·mol-1时,净光合速率(Pn)表现为直线上升;当CO2摩尔分数大于 700 μmol·mol-1时,净光合速率保持平衡,变化不大。所以,兴安鹿蹄草新叶CO2饱和点的摩尔分数为 700 μmol·mol-1。从表 2 中还可看出,当胞间 CO2摩尔分数高于 200 μmol·mol-1时,7月份兴安鹿蹄草新叶的净光合速率最大,6月份净光合速率最小,2个月的净光合速率差异显著;8、9月份净光合速率大于6月份的,8、9月份的净光合速率相近,差异不大。6、7、8、9月份新叶的光呼吸速率分别为-1.330、-0.932、-1.523、-1.795 μmol·m-2·s-1,9 月份的光呼吸速率是最高的。

表2 4—9月份兴安鹿蹄草老叶、新叶在不同CO2摩尔分数下的净光合速率变化
3.2 气孔导度(Gs)对CO2摩尔分数升高的响应
CO2既是光合作用的原料同时也是影响气孔导度的重要因素。当胞间CO2摩尔分数增高时,光合作用效率升高,同时气孔导度减少[17]。
从表3可以看出,老叶与新叶在不同月份随着胞间CO2摩尔分数的升高,气孔导度呈现不同的变化规律。兴安鹿蹄草老叶在4月份随着胞间CO2摩尔分数的升高,气孔导度呈降低的趋势。不过气孔导度降低幅度并不大,胞间CO2摩尔分数为1 600 μmol·mol-1时的气孔导度,比胞间 CO2摩尔分数为0时的气孔导度降低了8.48%。从表3中可以看出,兴安鹿蹄草老叶4、5月份的气孔导度明显高于6月份。

表3 4—9月份兴安鹿蹄草老叶、新叶随着CO2摩尔分数变化气孔导度的变化规律
从表3中可以看出,5月份气孔导度呈单峰曲线,最高峰出现在胞间CO2摩尔分数为350 μmol·mol-1处,当胞间 CO2摩尔分数从 0 到 350 μmol·mol-1时,气孔导度逐渐增加,而大于 350 μmol·mol-1时气孔导度逐渐下降。从5月份的数据来看,当胞间CO2摩尔分数小于大气CO2摩尔分数时(350 μmol·mol-1),气孔导度逐渐增加,让更多的CO2进入气孔,保证叶片的光合作用。但是当胞间CO2摩尔分数大于大气CO2摩尔分数时,由于植物为保持胞间CO2分压始终低于大气CO2分压,可以调节气孔关闭来降低胞间CO2摩尔分数,因此,气孔导度降低。6月份气孔导度也呈单峰曲线,最高峰出现在胞间 CO2摩尔分数为 1 000 μmol·mol-1处。6月份兴安鹿蹄草所生活的环境基本在林荫下,光照较少,且叶片处于老龄化叶片,光合产物较少,胞间CO2增加,使保卫细胞内糖摩尔分数增加数量有限,调节气孔关闭的效果并不明显。
6月份以后兴安鹿蹄草老叶相继死去,新叶代替老叶。6月份的新叶处于幼龄阶段。其气孔导度的平均值略高于6月份的老叶。7月份的兴安鹿蹄草新叶气孔导度与6月份的差异不显著(F=0.853 0,FCcrit=4.600),8月份达到最大值,9月份迅速下降。6、7月份气孔导度较低,可能与叶片处于幼龄阶段有关。8月份叶片展开,处于生长旺盛期,光合作用也处于旺盛时期,而且此期温度较高,许多草本相继枯萎,所以此期的气孔导度最大。9月份由于温度开始降低,所以兴安鹿蹄草的叶片气孔导度也处于最低值。
3.3 蒸腾速率(Tr)对CO2摩尔分数升高的响应
蒸腾速率(Tr)的大小取决于气孔导度与叶面饱和蒸气压亏缺(Vp,d,l)。气孔的开闭程度影响植物叶片水分的散失。从表4可以看出,蒸腾速率的大小与气孔导度的大小成显著正相关。4、5、6月份老叶的蒸腾速率与气孔导度的相关系数分别为0.955 9、0.974 3、0.634 2;6、7、8、9 月份新叶的蒸腾速率与气孔导度的相关系数分别为0.991 1、0.974 5、0.946 0、0.839 1。从表4 中可看出,9 月份的蒸腾速率比8月份略高。而气孔导度9月份却低于8月份。这主要是由于影响蒸腾作用的因素除了有气孔导度外还受空气温湿度的影响[18],9月份空气相对湿度较低。
9月份空气湿度变小,气孔与外界的饱和水气压差变大,加速了蒸腾作用。从表4中还可以看出,5、6月份老叶的蒸腾速率变化不大(F=0.669 6,FCcrit=4.600 0),而从表3中可以看出6月份的兴安鹿蹄草老叶气孔导度显著低于5月份,差异很大(F=16.903 3,FCcrit=4.600)。这说明蒸腾速率除了与气孔导度有关之外,还与温度有很大的关系。气孔导度降低会使叶面温度有所升高。6月份气孔导度变小,叶片温度也相应升高,较高的温度对叶片蒸腾起加强作用[19]。
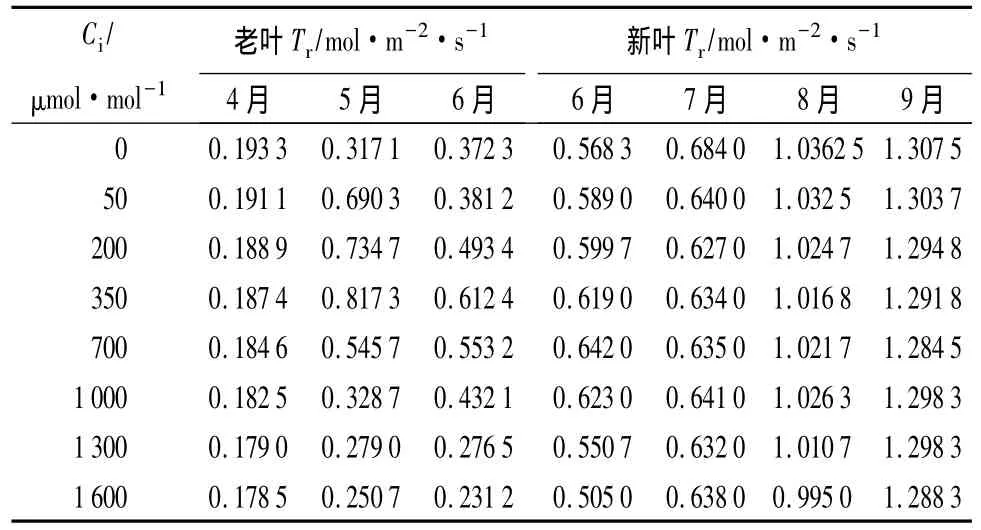
表4 4—9月份兴安鹿蹄草老叶、新叶在不同CO2摩尔分数下的蒸腾速率变化
3.4 饱和蒸汽压亏缺(Vp,d,l)对 CO2 摩尔分数升高的响应
饱和蒸汽压亏缺(Vp,d,l)为在同温度下饱和蒸汽压和实际蒸汽压之差,它是影响叶片蒸腾的重要因素之一。从表5中可以看出,各个月份兴安鹿蹄草叶片饱和蒸汽压亏缺几乎不随胞间CO2摩尔分数的变化而变化。但各个月份差异较大。兴安鹿蹄草老叶5月份饱和蒸汽压亏缺最大,6月份次之,4月份最小。这与老叶的蒸腾速率变化规律是一致的。兴安鹿蹄草新叶饱和蒸汽压亏缺由大到小的顺序为7、6、8、9月。这与蒸腾速率的变化规律不一致,说明兴安鹿蹄草新叶的蒸腾速率不只受饱和蒸汽压亏缺一个因素的影响。

表5 4—9月份兴安鹿蹄草老叶、新叶在不同CO2摩尔分数下的饱和蒸汽压亏缺变化
3.5 水分利用效率(WU,E)对CO2摩尔分数升高的响应
水分利用效率是指消耗单位质量的水所固定营养物质的量,是由光合速率和蒸腾速率两个方面的因素决定的,是光合特性与蒸腾特性的综合反映[20]。CO2的升高对水分利用效率有显著的影响,这主要是由于在高摩尔分数CO2条件下气孔关闭,外界CO2不易进入气孔。植物可以利用胞间一定量CO2和水分进行光合作用,消耗单位质量的水所固定CO2的数量增加,所以随着CO2摩尔分数的增加WU,E也在增加[21]。从表6中可以看出,兴安鹿蹄草老叶与新叶在各个月份随着CO2摩尔分数的升高WU,E都表现出逐渐增高的趋势。但是各个月份之间具有明显的差异,4月份老叶,8、9月份新叶WU,E较低,且差异不显著。外界的环境因子也对植物WU,E起着十分重要的作用,4月份温度较低,所以WU,E也较低;8、9月气候干燥,降水量少,土壤湿度很低,能被兴安鹿蹄草利用的水分十分有限,且蒸腾强度相对较高,故WU,E较小。这与刘金祥等[17]的研究结果是一致的。5月份老叶初始阶段(CO2摩尔分数<350 μmol·mol-1)WU,E升高缓慢;从胞间CO2摩尔分数为 350 μmol·mol-1开始 WU,E直线上升,而且高于所有月份;胞间CO2摩尔分数为1 600 μmol·mol-1时的 WU,E值比 CO2摩尔分数为 350 μmol·mol-1时的升高了 728.9%。6、7 月份新叶WU,E在胞间 CO2摩尔分数小于 700 μmol·mol-1时高于其他各月份的,大于 700 μmol·mol-1时,WU,E低于5月份老叶的,高于其他月份的;6月份老叶WU,E对高摩尔分数CO2比较敏感,当CO2摩尔分数大于1 000 μmol·mol-1时,WU,E有一个明显的升高过程。
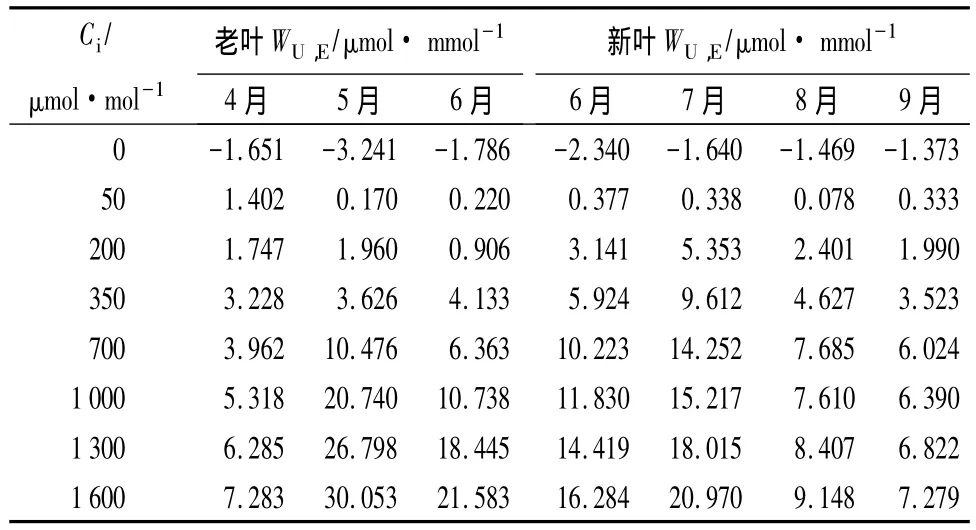
表6 4—9月份兴安鹿蹄草老叶、新叶在不同CO2摩尔分数下的水分利用效率变化
4 结论与讨论
Nozomi et al.[22]指出,红花鹿蹄草(P.incarnata)由于受到光线及雪后融雪的影响,68%的年度净产量形成于树冠枝叶完全展开之前(即4—5月份)。Hiroshi[23]指出,日本鹿蹄草(P.japonica)全年的发育和物质积累分为2个阶段,第一阶段为4—6月份,主要表现为老叶及其他构件的发育;第二阶段为7—翌年3月份,表现为新叶及地下茎的发育和物质的积累。其第二阶段又分为2个亚阶段,7—9月份是第一个亚阶段,10—翌年3月份为第二个亚阶段。净光合产量的最大值出现在5月份,原因是在5月份呼吸损耗低的缘故。7—9月份净光和产量很低甚至为负值,原因在于其强烈的呼吸导致净光合产值减少。10—翌年3月份处于一个稳定阶段。兴安鹿蹄草老叶4—6月份的发育相当于日本鹿蹄草第一阶段发育。从表2中可看出5月份的净光合速率明显高于4、6月份的,与Hiroshi[23]研究的结论是一致的。兴安鹿蹄草新叶7—9月份的发育相当于日本鹿蹄草第二阶段发育的第一个亚阶段。7月份光合速率最高,6月份光合速率最小。各个月份差异不显著(F=0.220 1,FCcrit=2.946 7)。由于文中未对呼吸作用做探讨,其光合净产量变化机制留后续讨论。
5月份光呼吸速率最大,6月份光呼吸速率次之,4月份光呼吸速率最小。原因是温度影响了酶的活性,进而对光合速率和呼吸速率都会有影响。5月份的兴安鹿蹄草处于林下非密闭环境,温度较高,光照最强,所以其光呼吸速率也最强,可能对维护其高温下生长是有利的。6月份由于空气温度过高,使得酶失活,这样也会使得光合速率与呼吸速率降低。
植物通过控制气孔的开闭程度来协调对CO2的需要和水分的消耗。气孔导度就是反映气孔开闭程度的一个重要生理指标。影响气孔导度的因素很多,除了温度、辐射外,还与光照、相对湿度、饱和水气压差和CO2摩尔分数有关。王玉辉等[24]及周丽娜等[25]指出,气孔导度与温度成正相关,温度降低气孔导度也随之降低,。而且从4月份气孔导度变化规律看出,在低温下,气孔导度变化不大。所以气孔导度在不同的月份呈不同的变化规律。并且,同一植物的不同生育期所表现的变化也不尽一致[26]。兴安鹿蹄草各个月份气孔导度随CO2升高表现不同变化也证明了这一点。有的研究表明当温度较低时,随着气温的升高气孔导度随着升高,但是当温度超过一定阈值,气孔导度反而下降[25,27]。从表1中可以看出,4月份兴安鹿蹄草气孔导度主要受温度的影响,气孔导度很低,5月份较高,6、7月份由于气温处于全年最高月份,气孔导度反而减小较少,这与前人的研究结果是一致的。有的研究结果表明,随着CO2摩尔分数的升高,C3植物的气孔导度在一开始是下降的,并可引起蒸腾速率的下降,但某些种类气孔导度的下降则是短期,之后迅速升高[28]。从表3中5—9月份气孔导度的变化规律说明了这一点。
4、5月份气孔导度高于6月份的,说明气孔导度除了受温度影响之外,还受光照的影响。由于兴安鹿蹄草生长于针阔混交林下,在4、5月份,由于阔叶树还未长叶或叶片还相当小,此时阳光可以直接照射到地面,林下的照度比6月份的照度大,正由于强烈光照的影响,兴安鹿蹄草的气孔导度较大。左应海等[29]也指出,光合有效辐射、土壤相对含水量与气孔导度之间呈显著正相关。5月份由于气温与光照的综合作用,使兴安鹿蹄草老叶的气孔导度明显高于4、6月份。Hiroshi[30]指出,兴安鹿蹄草在5月份光合效率是最高的。气孔导度的增加可能与其光合效率的增加是相适应的。
总之,随着CO2摩尔分数的升高,兴安鹿蹄草老叶与新叶的净光合速率都有逐渐增高的趋势。兴安鹿蹄草老叶5月份的净光合速率明显高于4、6月份。气孔导度呈现不同的变化规律。兴安鹿蹄草老叶与新叶的蒸腾速率的大小与气孔导度的大小呈显著正相关。各个月份兴安鹿蹄草叶片饱和蒸汽压亏缺几乎不随胞间CO2摩尔分数的变化而变化。老叶的饱和蒸汽压亏缺由大到小的顺序为5、6、4月份,新叶的饱和蒸汽压亏缺由大到小的顺序为7、6、8、9月份。兴安鹿蹄草老叶与新叶在各个月份随着CO2摩尔分数的升高WU,E都表现出逐渐增高的趋势。5月份老叶从CO2摩尔分数为350 μmol·mol-1开始WU,E直线上升,而且高于所有月份。
[1]Genthon C,Barnola J M,Raynaud D,et al.Vostok ice core:climatic response to CO2and orbital forcing changes over the last climatic cycle[J].Nature,1987,329:414-418.
[2]肖玲,王开运,张远彬,等.岷江冷杉根际土壤微生物对大气CO2浓度和温度升高的响应[J].应用生态学报,2006,17(5):773-777.
[3]陈德祥,李意德,骆土寿,等.短期CO2浓度升高对雨林树种盘壳栎光合特性的影响[J].生态学报,2004,24(8):1622-1628.
[4]蒋跃林,张庆国,杨书运,等.28种园林植物对大气CO2浓度增加的生理生态反应[J].植物资源与环境学报,2006,15(2):1-6.
[5]孙加伟,赵天宏,付宇,等.CO2浓度升高对玉米叶片光合生理特性的影响[J].玉米科学,2009,17(2):81-85.
[6]蒋高明,渠春梅.北京山区辽东栎林中几种木本植物光合作用对CO2浓度升高的响应[J].植物生态学报,2000,24(2):204-208.
[7]张小全,徐德应,赵茂盛,等.CO2增长对杉木中龄林针叶光合生理生态的影响[J].生态学报,2000,20(3):390-396.
[8]胡文光,胡琳贞,宋滋圃.中国植物志:第56卷[M].北京:科学出版社,1990:169-171.
[9]马玉心,崔大练.兴安鹿蹄草叶片雪盖初期与雪盖后期保护酶活性的变化规律[J].植物生态学报,2009,33(2):355-360.
[10]王军宪,陈新民,李宏,等.鹿衔草化学成分的研究:羟基肾叶鹿蹄草甙的结构鉴定[J].植物学报,1994,36(11):895-897.
[11]马玉心,蔡体久,宋丽萍,等.兴安鹿蹄草(Pyrola dahurica(H.Andr.)Kom.)雪盖前后丙二醛及渗透调节物质含量的变化[J].生态学报,2007,27:(11):4596-4602.
[12]周云龙,刘宁,刘全儒,等.植物生物学[M].3版,北京:高等教育出版社,2011.
[13]蔡时青,许大全.大豆叶片CO2补偿点和光呼吸的关系[J].植物生理学报,2000,26(6):545-550.
[14]Smith E W,Tolbert N E,Ku H S.Variables affecting the CO2compensation point[J].Plans Physiol,1976,58(2):143-146.
[15]Azcon-Bieto J,Farquhar G D,Caballero A.Effect of temperature.oxygen concentration,leaf age and seasonal variations on the CO2compensation point of Loliurn perenne L.[J].Planta,1981,152:497-504.
[16]Chen Z,Spreitzer R J.How various factors influence the CO2/O2specificity,of ribulose-1,5-bisphosphate carboxylase oxygenase[J].Photosynthesis Res,1992,31:157-164.
[17]刘金祥,麦嘉玲,刘家琼.CO2浓度增强对沿阶草光合生理特性的影响[J].中国草地,2004,26(3):13-17.
[18]张利平,王新平,刘立超,等.沙坡头主要建群植物油蒿和柠条的气体交换特征研究[J].生态学报,1998,18(2):133-137.
[19]Kinball B A,朱建国,程磊,等.开放系统中农作物对空气CO2浓度增加的响应[J].应用生态学报,2002,13(10):1323-1338.
[20]王旭军,吴际友,廖德志,等.响叶杨光合蒸腾和水分利用效率对光强及CO2浓度升高的响应[J].南京林业大学学报:自然科学版,2009,33(2):55-59.
[21]林伟宏.植物光合作用对大气CO2浓度升高的反应[J].生态学报,1998,18(5):529-538.
[22]Nozomi I,Yasuo Y,Shigeru M,et al.Seasonal pattern of photosynthetic production in a subalpine evergreen herb,Pyrola incarnata[J].J Plant Res,2003,116:199-206.
[23]Hiroshi K.Studies on the life history of an evergreen herb,Pyrola japonica,population on a forest floor in a warm temperate region 1.growth,net production and matter economy[J].Bot Mag Tokyo,1998,98:383-392.
[24]王玉辉,周广盛.松嫩草地羊草叶片光合作用生理生态特征分析[J].应用生态学报,2001,12(1):75-79.
[25]周丽娜,刘艳,张蕾,等.林下参叶片气孔导度与环境因子的关系研究[J].安徽农业科学,2009,37(33):16616-16618.
[26]王润佳,高世铭,张绪成.高大气CO2浓度下C3植物叶片水分利用效率升高的研究进展[J].干旱地区农业研究,2010,28(6):190-195.
[27]苏天星,杨再强,黄海静,等.不同光质对温室甜椒气孔导度的影响[J].干旱气象,2010,28(4):443-448.
[28]Bunce J.Stomatal,conductance,photosynthesis and respiration of temperate deciduous tree seedling grown outdoors at an elevated concentration of carbon dioxide[J].Plant Cell Environ,1992,15:541-549.
[29]左应梅,陈秋波,邓权权,等.土壤水分、光照和空气湿度对木薯气孔导度的影响[J].生态学杂志,2011,30(4):689-693.
[30]Hiroshi K.Studies on the life history of an evergreen herb,Pyrola japonica,population on a forest floor in a warm temperate region.2.photosynthesis,respiration and gross production[J].Bot Mag Tokyo,1989,102:521-532.
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
金山(2020年9期)2020-11-10 07:15:08
节水灌溉(2020年9期)2020-09-16 00:32:12
东坡赤壁诗词(2017年3期)2017-07-05 08:09:17
创新作文(小学版)(2017年29期)2017-04-04 02:04:22
语文世界(初中版)(2017年1期)2017-03-06 19:10:29
牡丹(2015年3期)2015-05-30 10:48:04
大江南北(2014年3期)2014-11-23 06:16:21
